
Nuclear Receptors Are Differentially Expressed and Activated in KAIMRC1 Compared to MCF7 and MDA-MB231 Breast Cancer Cells

 ,
,  , ,
, , 
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
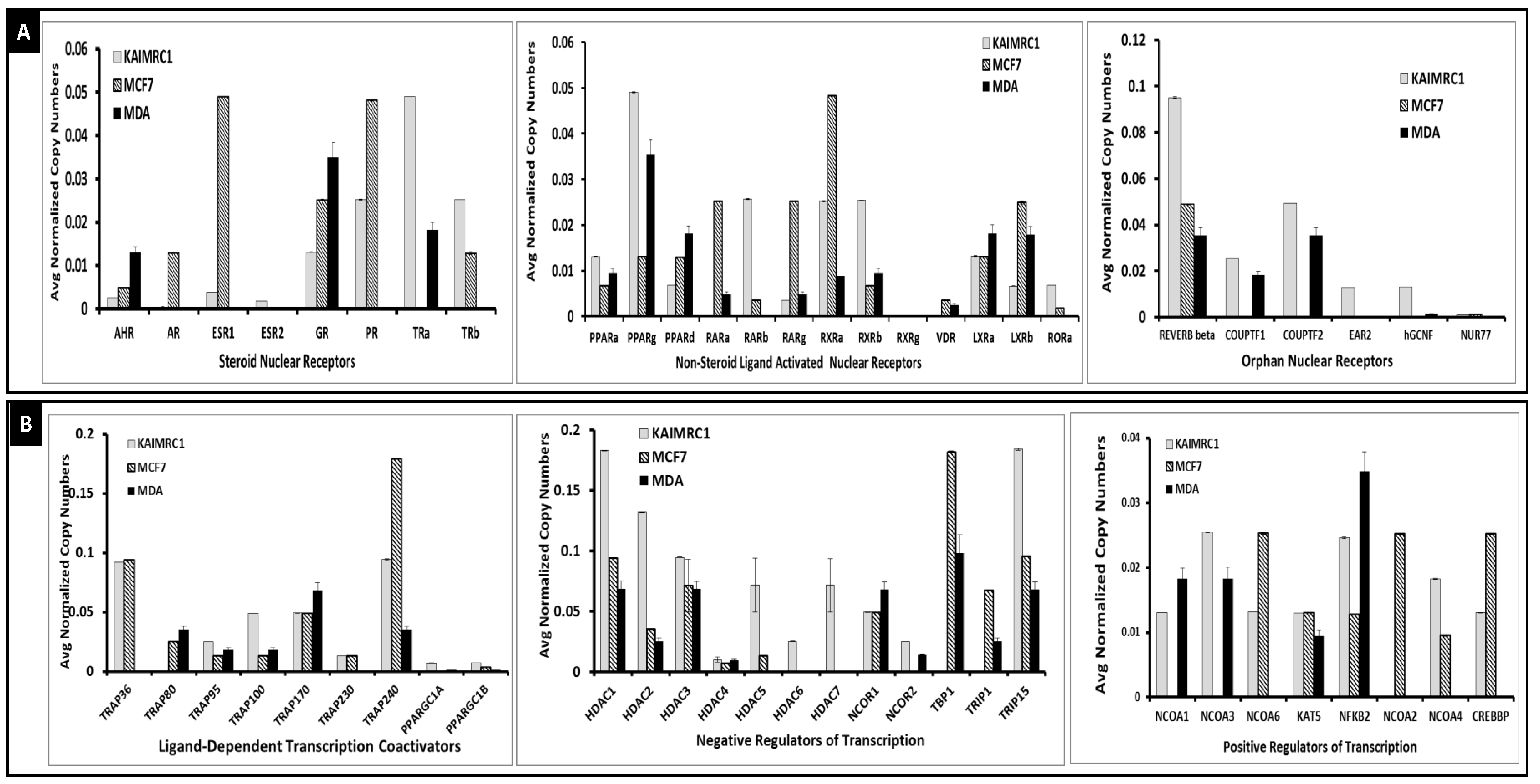
2.1. Nuclear Receptors and their Cofactor Expression in KAIMRC1 Compared to MCF7 and MDA-MB231
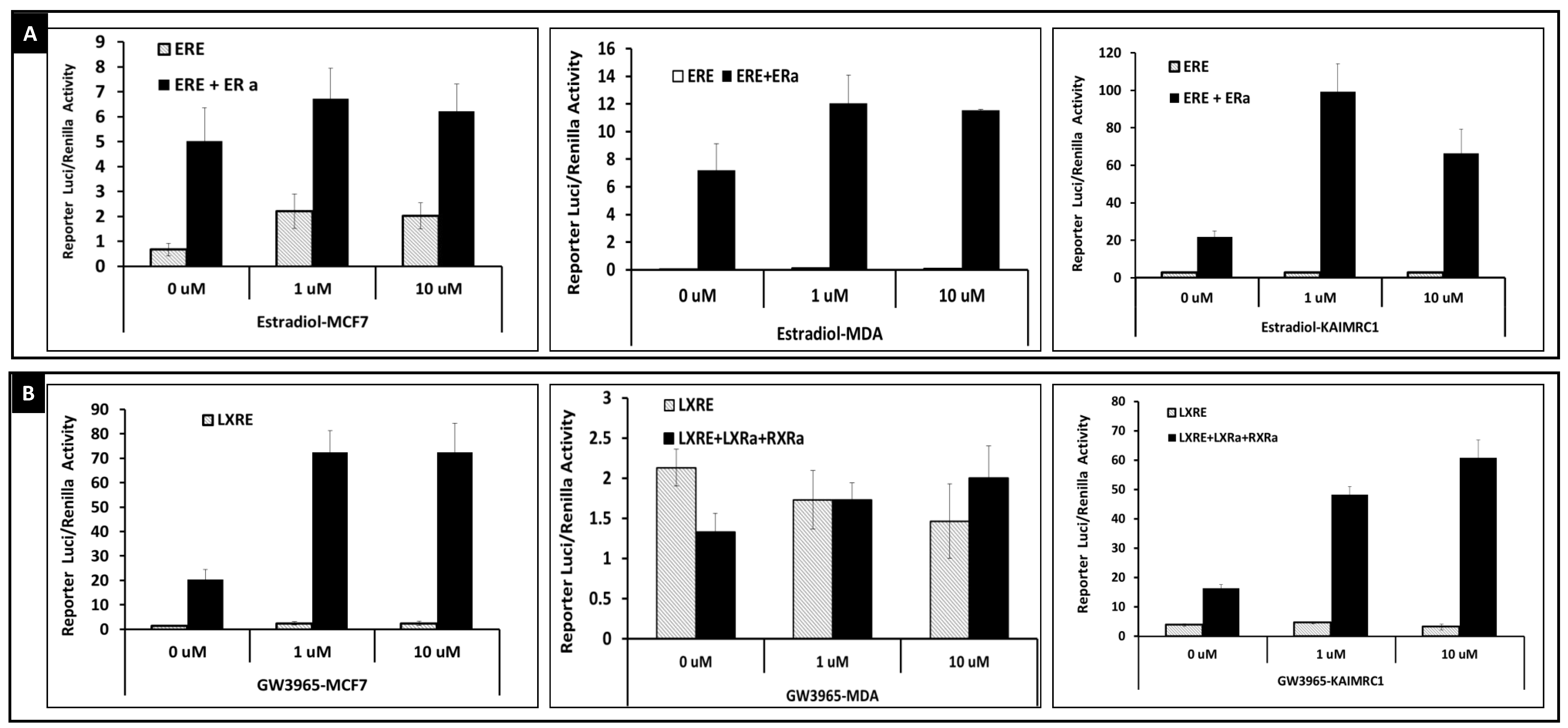
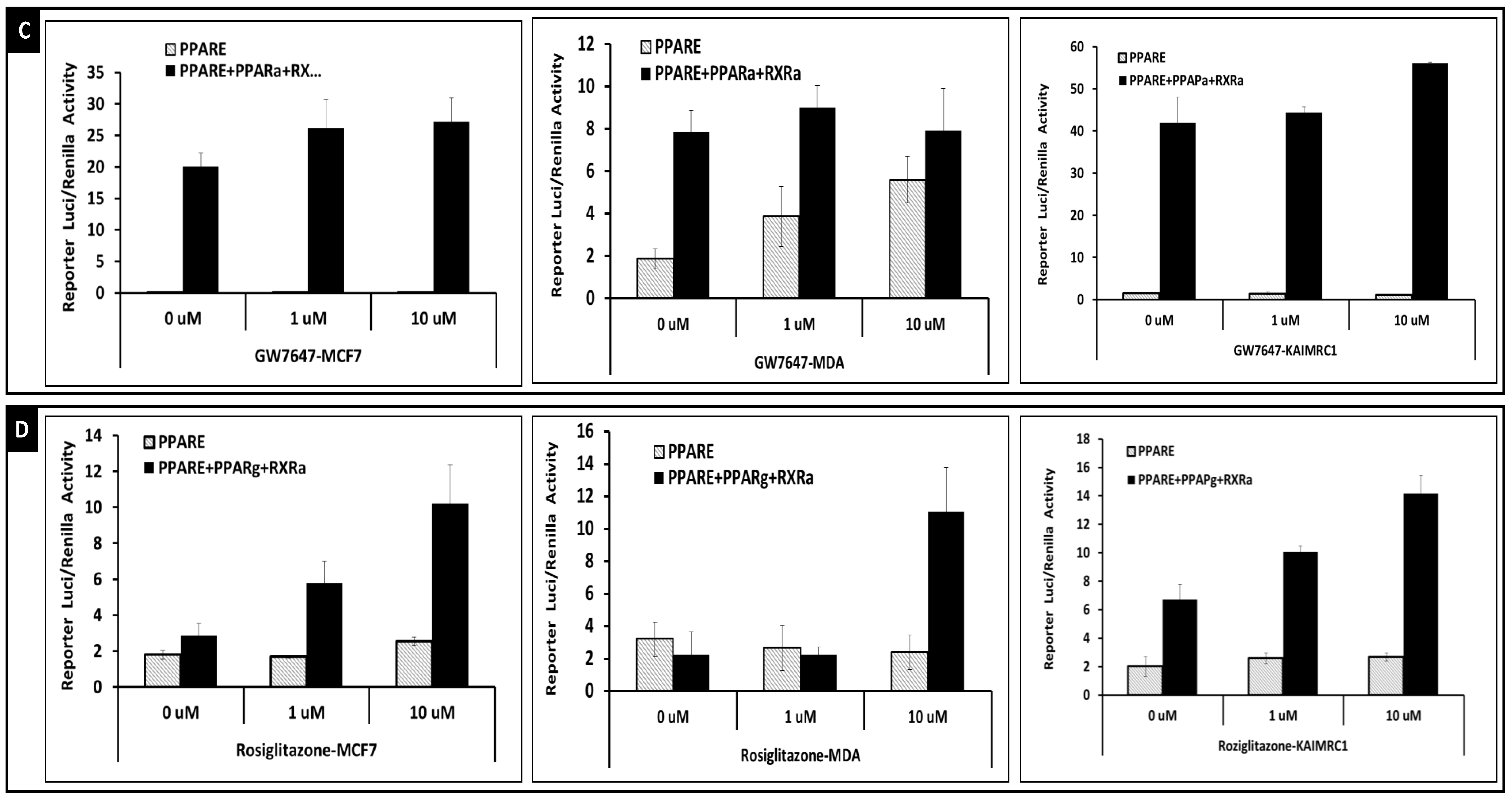
2.2. Transcriptional Activities of Nuclear Receptors in KAIMRC1, MCF7 and MDA-MB231 Cells
2.3. Effect of Nuclear Receptor Ligands on KAIMRC1 Proliferation Compared to MCF7 and MDA231 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plasmids
4.3. Cell Culture
4.4. Cell Transfection
4.5. Luciferase Reporter Assays
4.6. Cell Proliferation Assay
4.7. RNA Preparation and cDNA Synthesis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ali, R.; Samman, N.; Al Zahrani, H.; Nehdi, A.; Rahman, S.; Khan, A.L.; Balwi, M.A.; Alriyees, L.A.; Alzaid, M.; Askar, A.A.; et al. Isolation and characterization of a new naturally immortalized human breast carcinoma cell line, KAIMRC1. BMC Cancer 2017, 17, 803. [Google Scholar] [CrossRef]
- Conzen, S.D. Minireview: Nuclear receptors and breast cancer. Mol. Endocrinol. 2008, 22, 2215–2228. [Google Scholar] [CrossRef]
- Liao, L.; Kuang, S.Q.; Yuan, Y.; Gonzalez, S.M.; O’Malley, B.W.; Xu, J. Molecular structure and biological function of the cancer-amplified nuclear receptor coactivator SRC-3/AIB1. J. Steroid Biochem. Mol. Biol. 2002, 83, 3–14. [Google Scholar] [CrossRef]
- Anzick, S.L.; Kononen, J.; Walker, R.L.; Azorsa, D.O.; Tanner, M.M.; Guan, X.Y.; Sauter, G.; Kallioniemi, O.-P.; Trent, J.M.; Meltzer, P.S. AIB1, a steroid receptor coactivator amplified in breast and ovarian cancer. Science 1997, 277, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.A.; Cirrincione, C.; Henderson, I.C.; Citron, M.L.; Budman, D.R.; Goldstein, L.J. Estrogen-receptor status and outcomes of modern chemotherapy for patients with node-positive breast cancer. JAMA 2006, 295, 1658–1667. [Google Scholar] [CrossRef]
- Grimaldi, M.; Boulahtouf, A.; Delfosse, V.; Thouennon, E.; Bourguet, W.; Balaguer, P. Reporter Cell Lines for the Characterization of the Interactions between Human Nuclear Receptors and Endocrine Disruptors. Front. Endocrinol. (Lausanne) 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuclear Receptors Committee. A unified nomenclature system for the nuclear receptor superfamily. Cell 1999, 97, 161–163. [Google Scholar] [CrossRef]
- Weigel, N.L.; Zhang, Y. Ligand-independent activation of steroid hormone receptors. J. Mol. Med. (Berl) 1998, 76, 469–479. [Google Scholar] [CrossRef]
- Simons, S.S., Jr.; Edwards, D.P.; Kumar, R. Minireview: Dynamic structures of nuclear hormone receptors: New promises and challenges. Mol. Endocrinol. 2014, 28, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear Receptors, RXR, and the Big Bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Bourguet, W.; Germain, P.; Gronemeyer, H. Nuclear receptor ligand-binding domains: Three-dimensional structures, molecular interactions and pharmacological implications. Trends Pharmacol. Sci. 2000, 21, 381–388. [Google Scholar] [CrossRef]
- Smirnov, A.N. Nuclear receptors: Nomenclature, ligands, mechanisms of their effects on gene expression. Biochem. (Mosc) 2002, 67, 957–977. [Google Scholar] [CrossRef]
- Weigel, N.L.; Moore, N.L. Steroid receptor phosphorylation: A key modulator of multiple receptor functions. Mol. Endocrinol. 2007, 21, 2311–2319. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Weigel, N.L. Steroid receptor phosphorylation: Assigning function to site-specific phosphorylation. Biofactors 2009, 35, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Carey, L.A.; Dees, E.C.; Sawyer, L.; Gatti, L.; Moore, D.T.; Collichio, F.; Ollila, D.W.; Sartor, C.I.; Graham, M.L.; Perouet, C.M. The triple negative paradox: Primary tumor chemosensitivity of breast cancer subtypes. Clin. Cancer Res. 2007, 13, 2329–2334. [Google Scholar] [CrossRef]
- Lapidus, R.G.; Ferguson, A.T.; Ottaviano, Y.L.; Parl, F.F.; Smith, H.S.; Weitzman, S.A.; Baylin, S.B.; Issa, J.P.; Davidson, N.E. Methylation of estrogen and progesterone receptor gene 5′ CpG islands correlates with lack of estrogen and progesterone receptor gene expression in breast tumors. Clin. Cancer Res. 1996, 2, 805–810. [Google Scholar]
- Hudelist, G.; Czerwenka, K.; Kubista, E.; Marton, E.; Pischinger, K.; Singer, C.F. Expression of sex steroid receptors and their co-factors in normal and malignant breast tissue: AIB1 is a carcinoma-specific co-activator. Breast Cancer Res. Treat. 2003, 78, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Esslimani-Sahla, M.; Simony-Lafontaine, J.; Kramar, A.; Lavaill, R.; Mollevi, C.; Warner, M.; Gustafsson, J.-Å.; Rochefort, H. Estrogen receptor beta (ER beta) level but not its ER beta cx variant helps to predict tamoxifen resistance in breast cancer. Clin. Cancer Res. 2004, 10, 5769–5776. [Google Scholar] [CrossRef]
- Bonofiglio, D.; Gabriele, S.; Aquila, S.; Catalano, S.; Gentile, M.; Middea, E.; Giordano, F.; Andò, S. Estrogen receptor alpha binds to peroxisome proliferator-activated receptor response element and negatively interferes with peroxisome proliferator-activated receptor gamma signaling in breast cancer cells. Clin. Cancer Res. 2005, 11, 6139–6147. [Google Scholar] [CrossRef]
- Yin, Y.; Russell, R.G.; Dettin, L.E.; Bai, R.; Wei, Z.L.; Kozikowski, A.P.; Kopleovich, L.; Glazer, R.I. Peroxisome proliferator-activated receptor delta and gamma agonists differentially alter tumor differentiation and progression during mammary carcinogenesis. Cancer Res. 2005, 65, 3950–3957. [Google Scholar] [CrossRef]
- Ishay-Ronen, D.; Diepenbruck, M.; Kalathur, R.K.R.; Sugiyama, N.; Tiede, S.; Ivanek, R.; Bantug, G.; Morini, M.F.; Wang, J.; Hess, C.; et al. Gain Fat-Lose Metastasis: Converting Invasive Breast Cancer Cells into Adipocytes Inhibits Cancer Metastasis. Cancer Cell 2019, 35, 17–32. [Google Scholar] [CrossRef]
- Koeffler, H.P. Peroxisome proliferator-activated receptor gamma and cancers. Clin. Cancer Res. 2003, 9, 1–9. [Google Scholar]
- Stephen, R.L.; Gustafsson, M.C.; Jarvis, M.; Tatoud, R.; Marshall, B.R.; Knight, D.; Ehrenborg, E.; Harris, A.L.; Wolf, C.R.; Palmer, C.N.A. Activation of peroxisome proliferator-activated receptor delta stimulates the proliferation of human breast and prostate cancer cell lines. Cancer Res. 2004, 64, 3162–3170. [Google Scholar] [CrossRef]
- Seewaldt, V.L.; Kim, J.H.; Caldwell, L.E.; Johnson, B.S.; Swisshelm, K.; Collins, S.J. All-trans-retinoic acid mediates G1 arrest but not apoptosis of normal human mammary epithelial cells. Cell Growth Differ. 1997, 8, 631–641. [Google Scholar]
- Miller, V.A.; Rigas, J.R.; Benedetti, F.M.; Verret, A.L.; Tong, W.P.; Kris, M.G.; Gill, G.M.; Loewen, G.R.; Truglia, J.A.; Ulm, E.H.; et al. Initial clinical trial of the retinoid receptor pan agonist 9-cis retinoic acid. Clin. Cancer Res. 1996, 2, 471–475. [Google Scholar]
- Zinser, G.M.; Tribble, E.; Valrance, M.; Urben, C.M.; Knutson, J.C.; Mazess, R.B.; Strugnell, S.A.; Welsh, J. 1,24(S)-dihydroxyvitamin D2, an endogenous vitamin D2 metabolite, inhibits growth of breast cancer cells and tumors. Anticancer Res. 2005, 25, 235–241. [Google Scholar]
- Ariazi, E.A.; Clark, G.M.; Mertz, J.E. Estrogen-related receptor alpha and estrogen-related receptor gamma associate with unfavorable and favorable biomarkers, respectively, in human breast cancer. Cancer Res. 2002, 62, 6510–6518. [Google Scholar]
- Kraus, R.J.; Ariazi, E.A.; Farrell, M.L.; Mertz, J.E. Estrogen-related receptor alpha 1 actively antagonizes estrogen receptor-regulated transcription in MCF-7 mammary cells. J. Biol. Chem. 2002, 277, 24826–24834. [Google Scholar] [CrossRef]
- Levin, E.R. Integration of the extranuclear and nuclear actions of estrogen. Mol. Endocrinol. 2005, 19, 1951–1959. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, V.; Edwards, D.P. Receptor mechanisms of rapid extranuclear signalling initiated by steroid hormones. Essays Biochem. 2004, 40, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Acconcia, F.; Bresciani, F.; Weisz, A.; Trentalance, A. Distinct nongenomic signal transduction pathways controlled by 17beta-estradiol regulate DNA synthesis and cyclin D(1) gene transcription in HepG2 cells. Mol. Biol. Cell 2002, 13, 3720–3729. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yu, D.D.; Yan, D.L.; Hu, Y.; Chen, D.; Liu, Y.; Zhang, H.D.; Yu, S.R.; Cao, H.X.; Feng, J.F. Liver X receptor as a drug target for the treatment of breast cancer. Anticancer Drugs 2016, 27, 373–382. [Google Scholar] [CrossRef]
- Nguyen-Vu, T.; Vedin, L.L.; Liu, K.; Jonsson, P.; Lin, J.Z.; Candelaria, N.R.; Candelaria, L.P.; Addanki, S.; Williams, C.; Gustafsson, J.-Å.; et al. Liver x receptor ligands disrupt breast cancer cell proliferation through an E2F-mediated mechanism. Breast Cancer Res. 2013, 15, R51. [Google Scholar] [CrossRef]
- Bensinger, S.J.; Bradley, M.N.; Joseph, S.B.; Zelcer, N.; Janssen, E.M.; Hausner, M.A.; Shih, R.; Parks, J.S.; Edwards, P.A.; Jamieson, B.D.; et al. LXR signaling couples sterol metabolism to proliferation in the acquired immune response. Cell 2008, 134, 97–111. [Google Scholar] [CrossRef]
- Gottardis, M.M.; Lamph, W.W.; Shalinsky, D.R.; Wellstein, A.; Heyman, R.A. The efficacy of 9-cis retinoic acid in experimental models of cancer. Breast Cancer Res. Treat. 1996, 8, 85–96. [Google Scholar] [CrossRef]
- IJpenberg, A.; Tan, N.S.; Gelman, L.; Kersten, S.; Seydoux, J.; Xu, J.; Metzger, D.; Canaple, L.; Chambon, P.; Wahli, W.; et al. In vivo activation of PPAR target genes by RXR homodimers. EMBO J. 2004, 23, 2083–2091. [Google Scholar] [CrossRef]
- Narvaez, C.J.; Zinser, G.; Welsh, J. Functions of 1alpha,25-dihydroxyvitamin D(3) in mammary gland: From normal development to breast cancer. Steroids 2001, 66, 301–308. [Google Scholar] [CrossRef]
- Townsend, K.; Banwell, C.M.; Guy, M.; Colston, K.W.; Mansi, J.L.; Stewart, P.M.; Campbell, M.J.; Hewison, M. Autocrine metabolism of vitamin D in normal and malignant breast tissue. Clin. Cancer Res. 2005, 11, 3579–3586. [Google Scholar] [CrossRef] [PubMed]
- Milliken, E.L.; Zhang, X.; Flask, C.; Duerk, J.L.; MacDonald, P.N.; Keri, R.A. EB1089, a vitamin D receptor agonist, reduces proliferation and decreases tumor growth rate in a mouse model of hormone-induced mammary cancer. Cancer Lett. 2005, 229, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giguere, V.; Yang, N.; Segui, P.; Evans, R.M. Identification of a new class of steroid hormone receptors. Nature 1988, 331, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.D.; Liu, X.; Zuo, F.; Eisenbraun, T.L.; Wiley, S.R.; Kraus, R.J.; Mertz, J.E. Estrogen-related receptor alpha 1 functionally binds as a monomer to extended half-site sequences including ones contained within estrogen-response elements. Mol. Endocrinol. 1997, 11, 342–352. [Google Scholar]
- Yamada, S.; Kakuta, H. Retinoid X receptor ligands: A patent review (2007–2013). Expert Opin. Ther. Pat. 2014, 24, 443–452. [Google Scholar] [CrossRef]
- Qin, C.; Burghardt, R.; Smith, R.; Wormke, M.; Stewart, J.; Safe, S. Peroxisome proliferator-activated receptor gamma agonists induce proteasome-dependent degradation of cyclin D1 and estrogen receptor alpha in MCF-7 breast cancer cells. Cancer Res. 2003, 63, 958–964. [Google Scholar]
- Bonofiglio, D.; Cione, E.; Vizza, D.; Perri, M.; Pingitore, A.; Qi, H.; Catalano, S.; Rovito, D.; Genchi, G.; Andò, S. Bid as a potential target of apoptotic effects exerted by low doses of PPARgamma and RXR ligands in breast cancer cells. Cell Cycle. 2011, 10, 2344–2354. [Google Scholar] [CrossRef]
- Andruska, N.; Mao, C.; Cherian, M.; Zhang, C.; Shapiro, D.J. Evaluation of a luciferase-based reporter assay as a screen for inhibitors of estrogen-ERalpha-induced proliferation of breast cancer cells. J. Biomol. Screen. 2012, 17, 921–932. [Google Scholar] [CrossRef]
- Guarnaccia, V. Retinoic Acid and Breast Cancer:how to Improve the Therapeutic Effect on the Basis of Molecular Knowledge. Ph.D. Thesis, Oncology and Genetics, Mario Negri Institute for Pharmacological Research, University of Siena, Milano, Italy, 2010. Available online: http://www3.unisi.it/ricerca/dottorationweb/genetica_medica/Tesi/Guarnaccia%20PhD%20thesis.pdf (accessed on 15 April 2019).
- Sanchez-Martinez, R.; Zambrano, A.; Castillo, A.I.; Aranda, A. Vitamin D-dependent recruitment of corepressors to vitamin D/retinoid X receptor heterodimers. Mol. Cell Biol. 2008, 28, 3817–3829. [Google Scholar] [CrossRef]
- Evans, R.M. The steroid and thyroid hormone receptor superfamily. Science 1988, 240, 889–895. [Google Scholar] [CrossRef]
- Germain, P.; Chambon, P.; Eichele, G.; Evans, R.M.; Lazar, M.A.; Leid, M.; Lera, A.R.D.; Lotan, R.; Mangelsdorf, D.J.; Gronemeyer, H. International Union of Pharmacology. LX. Retinoic acid receptors. Pharmacol. Rev. 2006, 58, 712–725. [Google Scholar] [CrossRef]
- Song, I.S.; Jeong, Y.J.; Jeong, S.H.; Kim, J.E.; Han, J.; Kim, T.H.; Jang, S.W. Modulation of Mitochondrial ERbeta Expression Inhibits Triple-Negative Breast Cancer Tumor Progression by Activating Mitochondrial Function. Cell Physiol. Biochem. 2019, 52, 468–485. [Google Scholar]
- Ortega-Olvera, C.; Ulloa-Aguirre, A.; Angeles-Llerenas, A.; Mainero-Ratchelous, F.E.; Gonzalez-Acevedo, C.E.; Hernandez-Blanco, M.L.; Ziv, E.; Avilés-Santa, L.; Pérez-Rodríguez, E.; Torres-Mejía, G. Thyroid hormones and breast cancer association according to menopausal status and body mass index. Breast Cancer Res. 2018, 20, 94. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Lu, J.; Zhao, B.; Wang, W.; Zhao, Y. Review of the possible association between thyroid and breast carcinoma. World, J. Surg. Oncol. 2018, 16, 130. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, S.; Gustafsson, J.A. Nuclear receptors: Recent drug discovery for cancer therapies. Endocr. Rev. 2019. [Google Scholar] [CrossRef]
- Lavinsky, R.M.; Jepsen, K.; Heinzel, T.; Torchia, J.; Mullen, T.M.; Schiff, R.; Del-Rio, A.L.; Ricote, M.; Ngo, S.; Gemsch, J.; et al. Diverse signaling pathways modulate nuclear receptor recruitment of N-CoR and SMRT complexes. Proc. Natl. Acad. Sci. USA 1998, 95, 2920–2925. [Google Scholar] [CrossRef] [Green Version]
- Girault, I.; Lerebours, F.; Amarir, S.; Tozlu, S.; Tubiana-Hulin, M.; Lidereau, R.; Bièche, I. Expression analysis of estrogen receptor alpha coregulators in breast carcinoma: Evidence that NCOR1 expression is predictive of the response to tamoxifen. Clin. Cancer Res. 2003, 9, 1259–1266. [Google Scholar]
- Keeton, E.K.; Brown, M. Cell cycle progression stimulated by tamoxifen-bound estrogen receptor-alpha and promoter-specific effects in breast cancer cells deficient in N-CoR and SMRT. Mol. Endocrinol. 2005, 19, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Keeton, E.K.; Brown, M. Coregulator expression and breast cancer: Improving the predictive power of estrogen receptor alpha. Clin. Cancer Res. 2003, 9, 1229–1230. [Google Scholar]
- Liu, N.; Zhao, R.; Ma, Y.; Wang, D.; Yan, C.; Zhou, D.; Yin, F.; Li, Z. The Development of Epigenetics and Related Inhibitors for Targeted Drug Design in Cancer Therapy. Curr. Top. Med. Chem. 2018, 18, 2380–2394. [Google Scholar] [CrossRef]
- Takimoto, G.S.; Graham, J.D.; Jackson, T.A.; Tung, L.; Powell, R.L.; Horwitz, L.D.; Horwitz, K.B. Tamoxifen resistant breast cancer: Coregulators determine the direction of transcription by antagonist-occupied steroid receptors. J. Steroid. Biochem. Mol. Biol. 1999, 69, 45–50. [Google Scholar] [CrossRef]
- Kastner, P.; Krust, A.; Turcotte, B.; Stropp, U.; Tora, L.; Gronemeyer, H.; Chambon, P. Two distinct estrogen-regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B. EMBO J. 1990, 9, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Vedin, L.L.; Lewandowski, S.A.; Parini, P.; Gustafsson, J.A.; Steffensen, K.R. The oxysterol receptor LXR inhibits proliferation of human breast cancer cells. Carcinogenesis 2009, 30, 575–579. [Google Scholar] [CrossRef]
- Kilgore, M.W.; Tate, P.L.; Rai, S.; Sengoku, E.; Price, T.M. MCF-7 and T47D human breast cancer cells contain a functional peroxisomal response. Mol. Cell Endocrinol. 1997, 129, 229–235. [Google Scholar] [CrossRef]
- Alimirah, F.; Peng, X.; Yuan, L.; Mehta, R.R.; von Knethen, A.; Choubey, D.; Mehta, R.G. Crosstalk between the peroxisome proliferator-activated receptor gamma (PPARgamma) and the vitamin D receptor (VDR) in human breast cancer cells: PPARgamma binds to VDR and inhibits 1alpha,25-dihydroxyvitamin D3 mediated transactivation. Exp. Cell Res. 2012, 318, 2490–2497. [Google Scholar] [CrossRef]
- Kogai, T.; Kanamoto, Y.; Brent, G.A. The modified firefly luciferase reporter gene (luc+) but not Renilla luciferase is induced by all-trans retinoic acid in MCF-7 breast cancer cells. Breast Cancer Res. Treat. 2003, 78, 119–126. [Google Scholar] [CrossRef]
- Tanaka, T.; Dancheck, B.L.; Trifiletti, L.C.; Birnkrant, R.E.; Taylor, B.J.; Garfield, S.H.; Thorgeirsson, U.; Luca, L.M.D. Altered localization of retinoid X receptor alpha coincides with loss of retinoid responsiveness in human breast cancer MDA-MB-231 cells. Mol. Cell Biol. 2004, 24, 3972–3982. [Google Scholar] [CrossRef]
- Tavera-Mendoza, L.E.; Westerling, T.; Libby, E.; Marusyk, A.; Cato, L.; Cassani, R.; Cameron, L.A.; Ficarro, S.B.; Marto, J.A.; Klawitter, J.; et al. Vitamin D receptor regulates autophagy in the normal mammary gland and in luminal breast cancer cells. Proc. Natl. Acad. Sci. USA 2017, 114, E2186–E2194. [Google Scholar] [CrossRef] [Green Version]
- Turunen, M.M.; Dunlop, T.W.; Carlberg, C.; Vaisanen, S. Selective use of multiple vitamin D response elements underlies the 1 alpha,25-dihydroxyvitamin D3-mediated negative regulation of the human CYP27B1 gene. Nucleic Acids Res. 2007, 35, 2734–2747. [Google Scholar] [CrossRef]
- Zheng, W.; Duan, B.; Zhang, Q.; Ouyang, L.; Peng, W.; Qian, F.; Wang, Y.; Huang, S. Vitamin D-induced vitamin D receptor expression induces tamoxifen sensitivity in MCF-7 stem cells via suppression of Wnt/beta-catenin signaling. Biosci. Rep. 2018, 38. [Google Scholar]
- Welsh, J. Vitamin D and breast cancer: Past and present. J. Steroid Biochem. Mol. Biol. 2018, 177, 15–20. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors, they can be purchased as indicated in Materials and Methods. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nehdi, A.; Ali, R.; Alhallaj, A.; Alzahrani, H.; Samman, N.; Mashhour, A.; Baz, O.; Barhoumi, T.; Alghanem, B.; Khan, A.; et al. Nuclear Receptors Are Differentially Expressed and Activated in KAIMRC1 Compared to MCF7 and MDA-MB231 Breast Cancer Cells. Molecules 2019, 24, 2028. https://doi.org/10.3390/molecules24112028
Nehdi A, Ali R, Alhallaj A, Alzahrani H, Samman N, Mashhour A, Baz O, Barhoumi T, Alghanem B, Khan A, et al. Nuclear Receptors Are Differentially Expressed and Activated in KAIMRC1 Compared to MCF7 and MDA-MB231 Breast Cancer Cells. Molecules. 2019; 24(11):2028. https://doi.org/10.3390/molecules24112028
Chicago/Turabian StyleNehdi, Atef, Rizwan Ali, Alshaimaa Alhallaj, Hajar Alzahrani, Nosaibah Samman, Abdullah Mashhour, Omar Baz, Tlili Barhoumi, Bandar Alghanem, Abdullatif Khan, and et al. 2019. "Nuclear Receptors Are Differentially Expressed and Activated in KAIMRC1 Compared to MCF7 and MDA-MB231 Breast Cancer Cells" Molecules 24, no. 11: 2028. https://doi.org/10.3390/molecules24112028
APA StyleNehdi, A., Ali, R., Alhallaj, A., Alzahrani, H., Samman, N., Mashhour, A., Baz, O., Barhoumi, T., Alghanem, B., Khan, A., Alriyees, L., & Boudjelal, M. (2019). Nuclear Receptors Are Differentially Expressed and Activated in KAIMRC1 Compared to MCF7 and MDA-MB231 Breast Cancer Cells. Molecules, 24(11), 2028. https://doi.org/10.3390/molecules24112028