
Paris Polyphylla Inhibits Colorectal Cancer Cells via Inducing Autophagy and Enhancing the Efficacy of Chemotherapeutic Drug Doxorubicin

 , and
, and 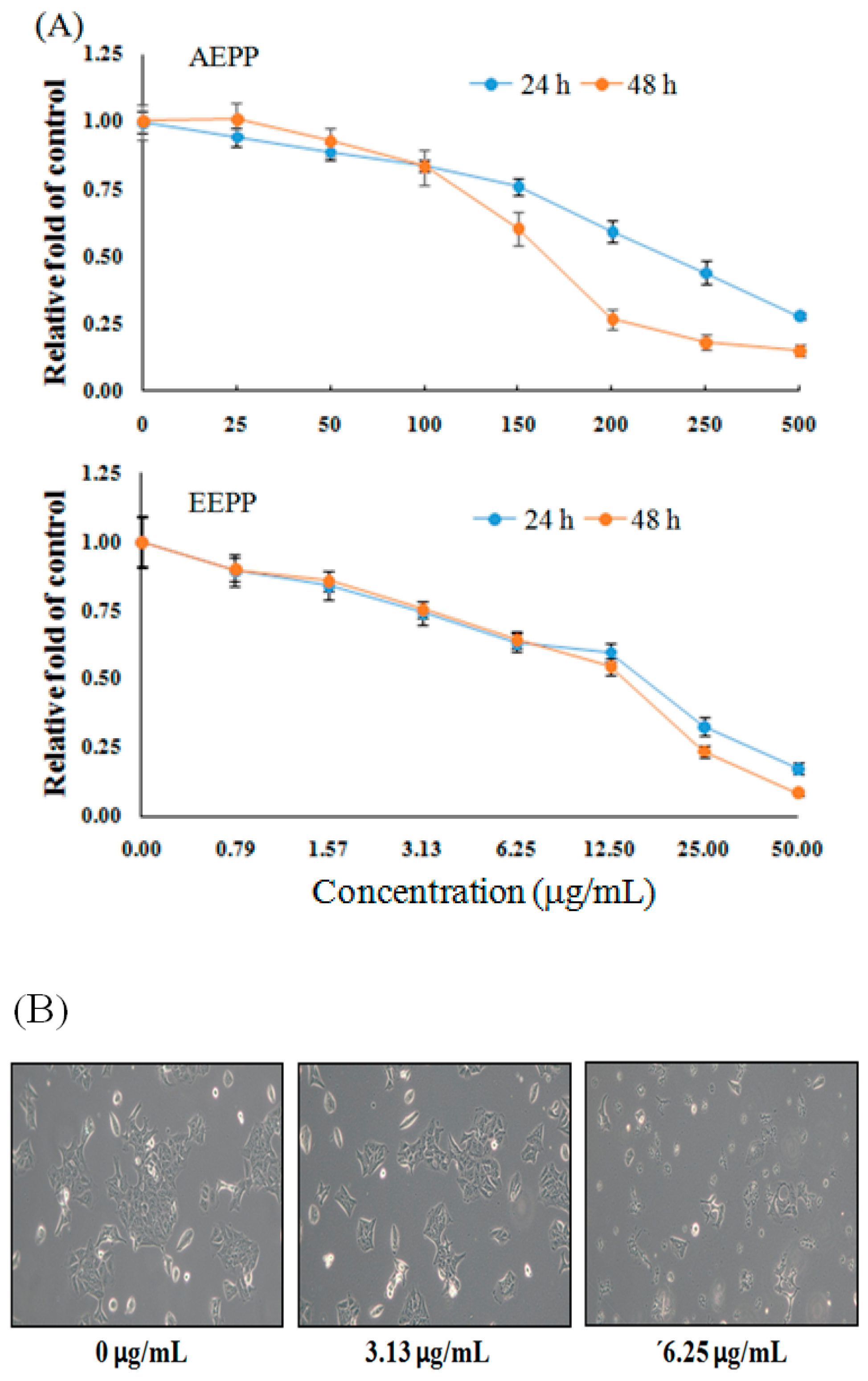{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
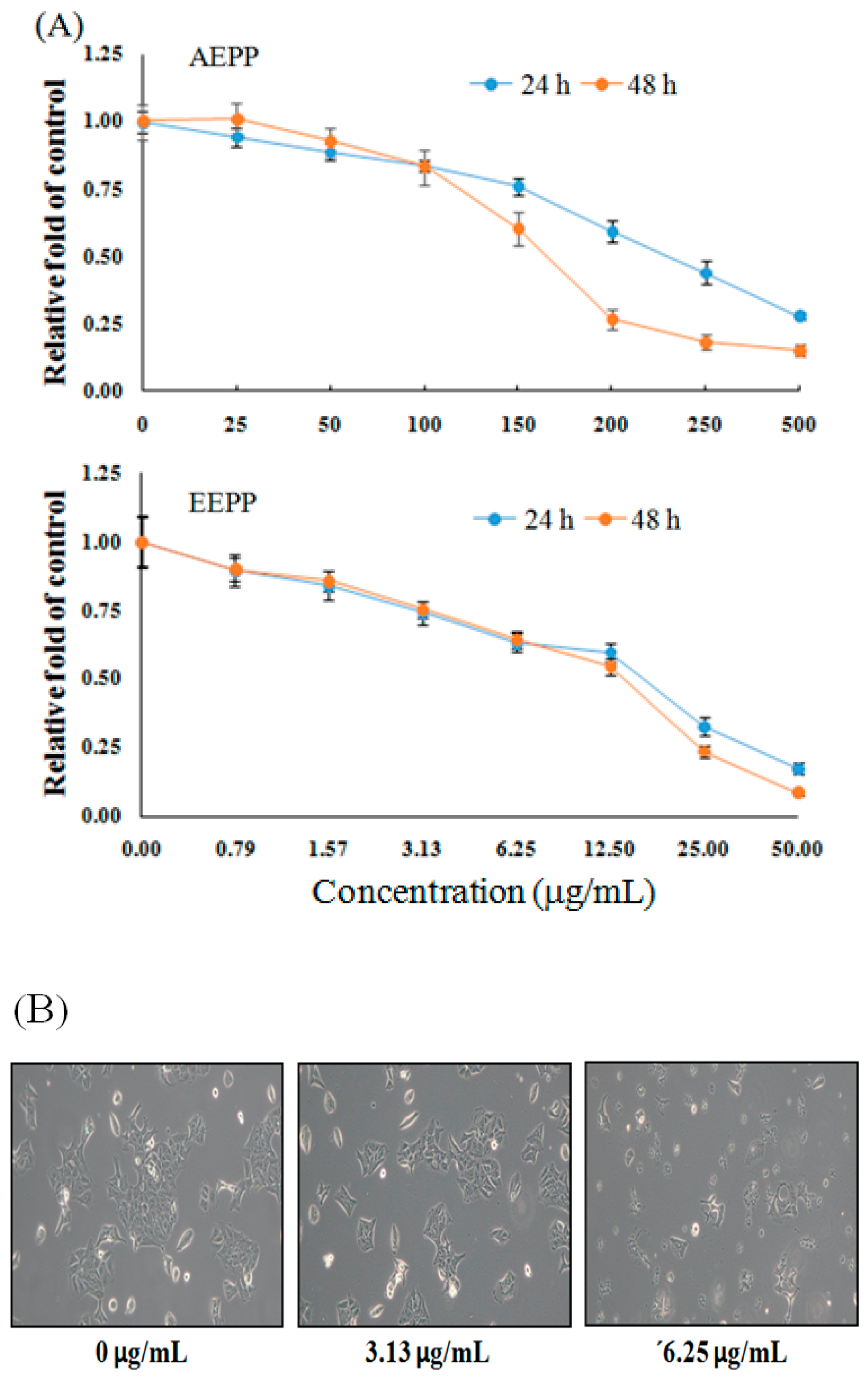
2.1. Treatment Effect of P. polyphylla on Colorectal Cancer Cell Growth
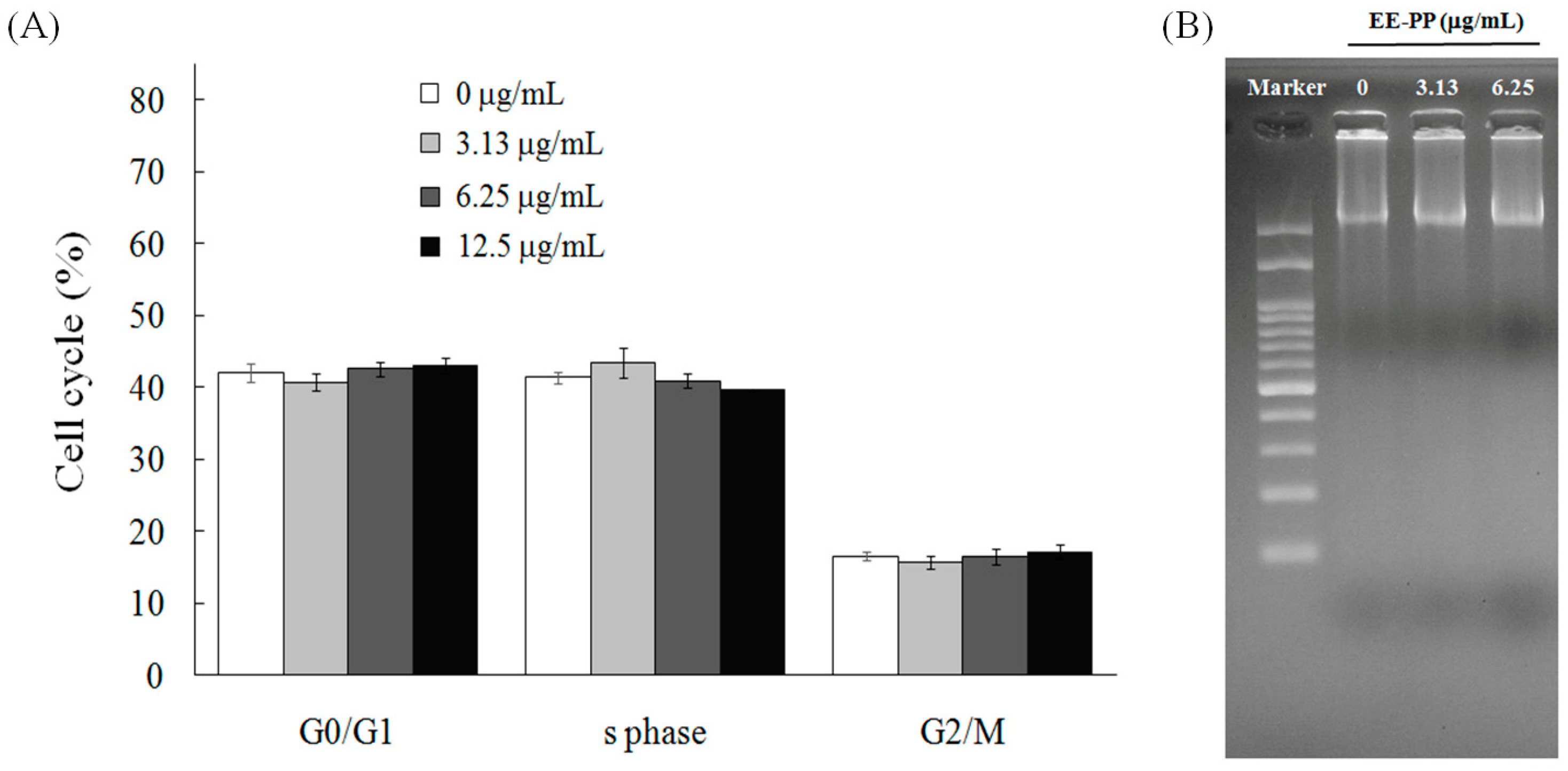
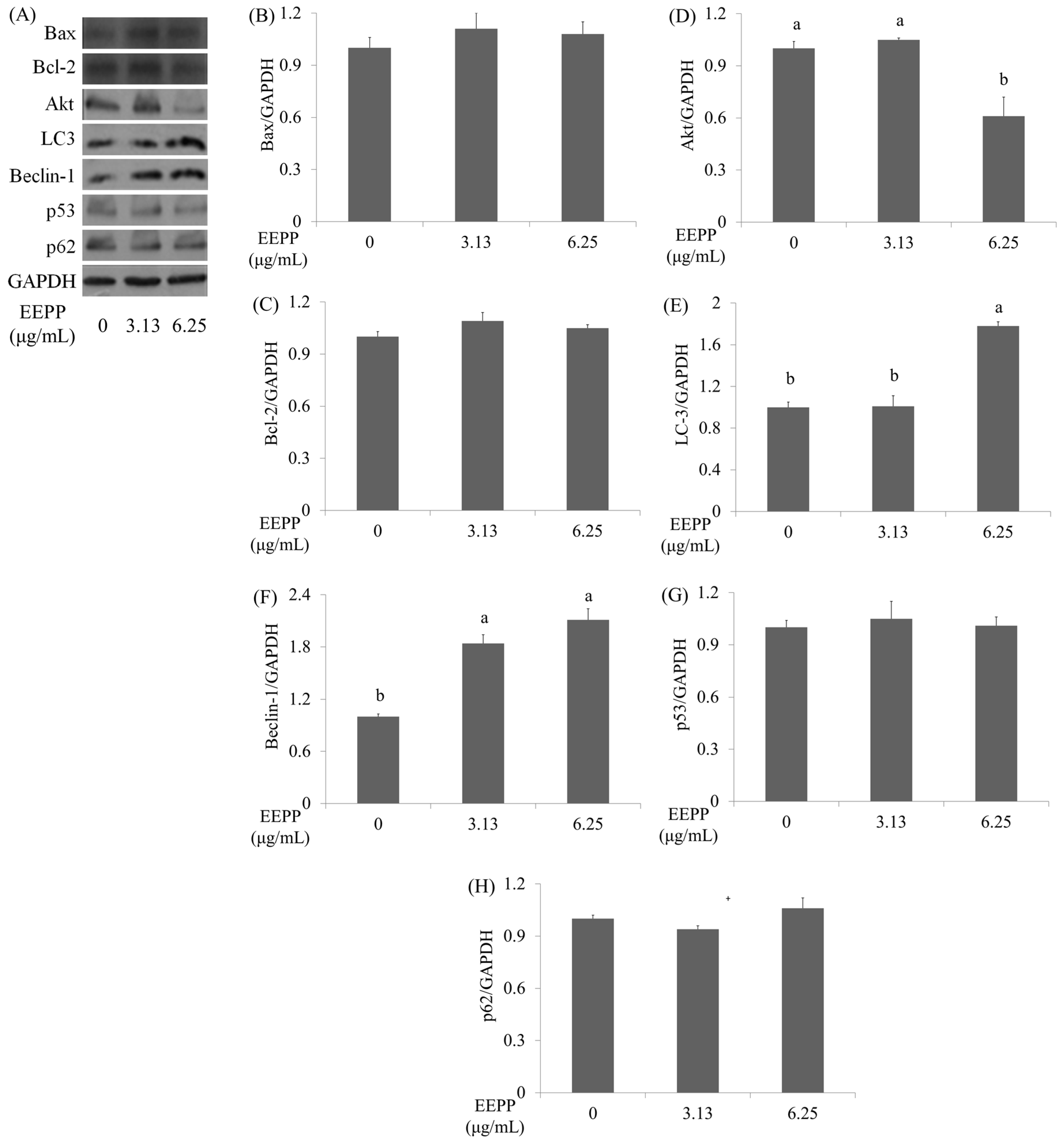
2.2. EEPP Treatment Causes Autophagic Cell Death in Colorectal Carcinoma Cells
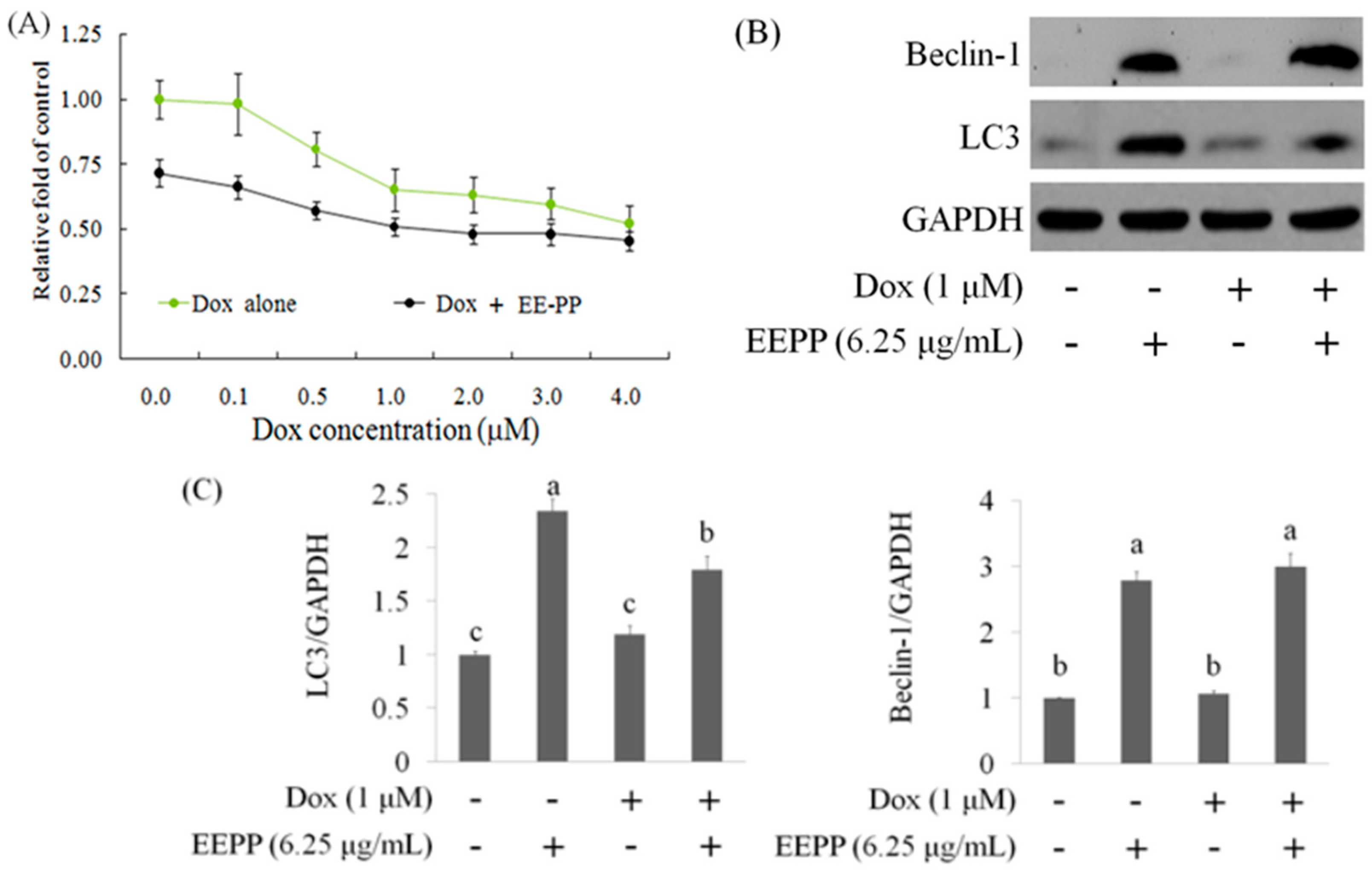
2.3. Effect of EEPP–Doxorubicin Combination Treatment on Autophagy Induction in Colorectal Carcinoma Cells
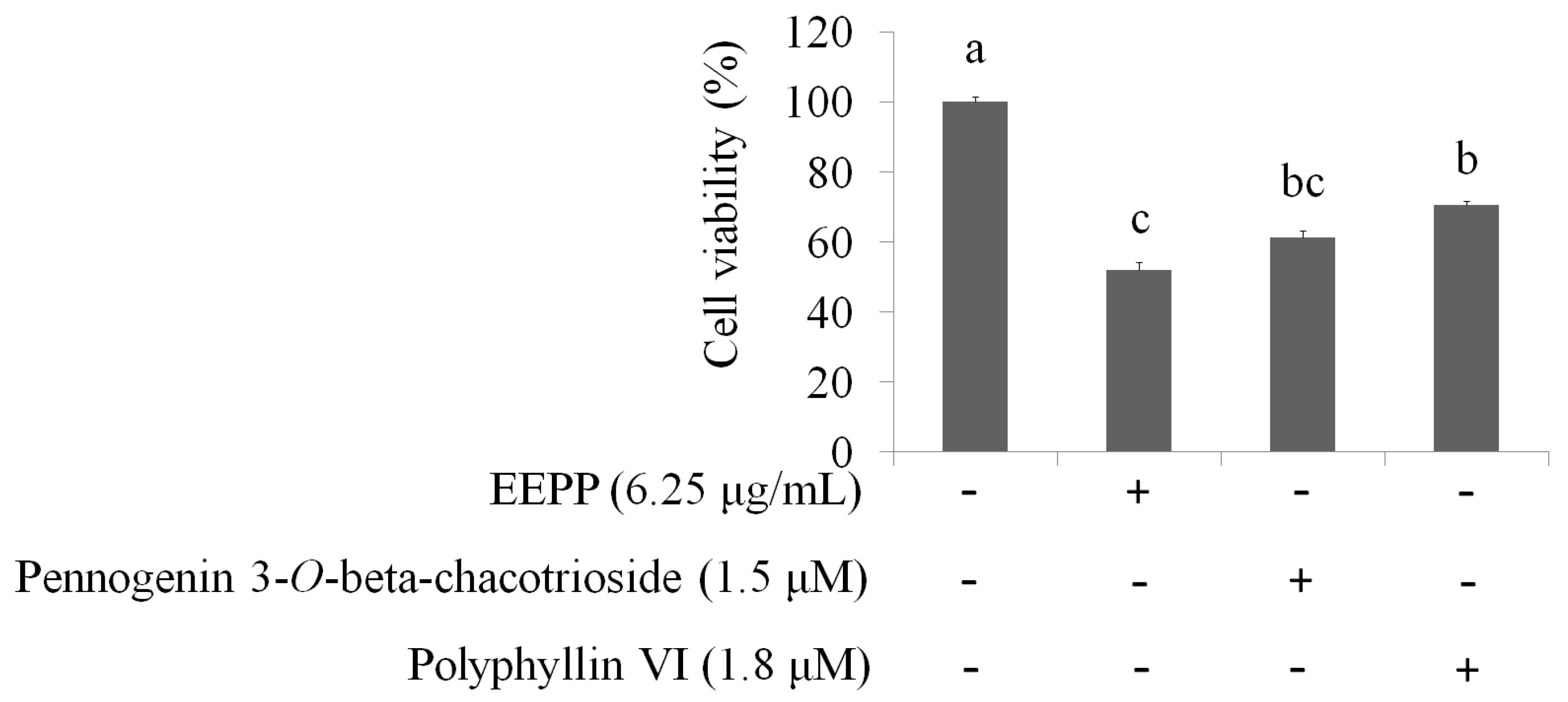
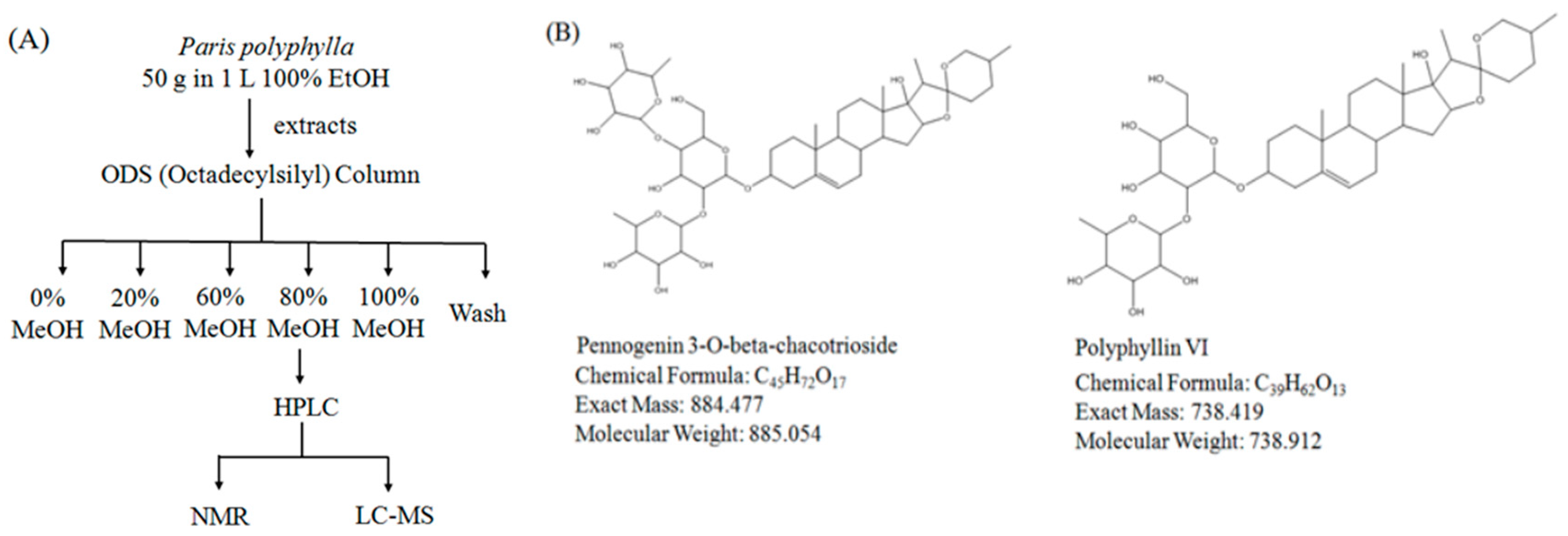
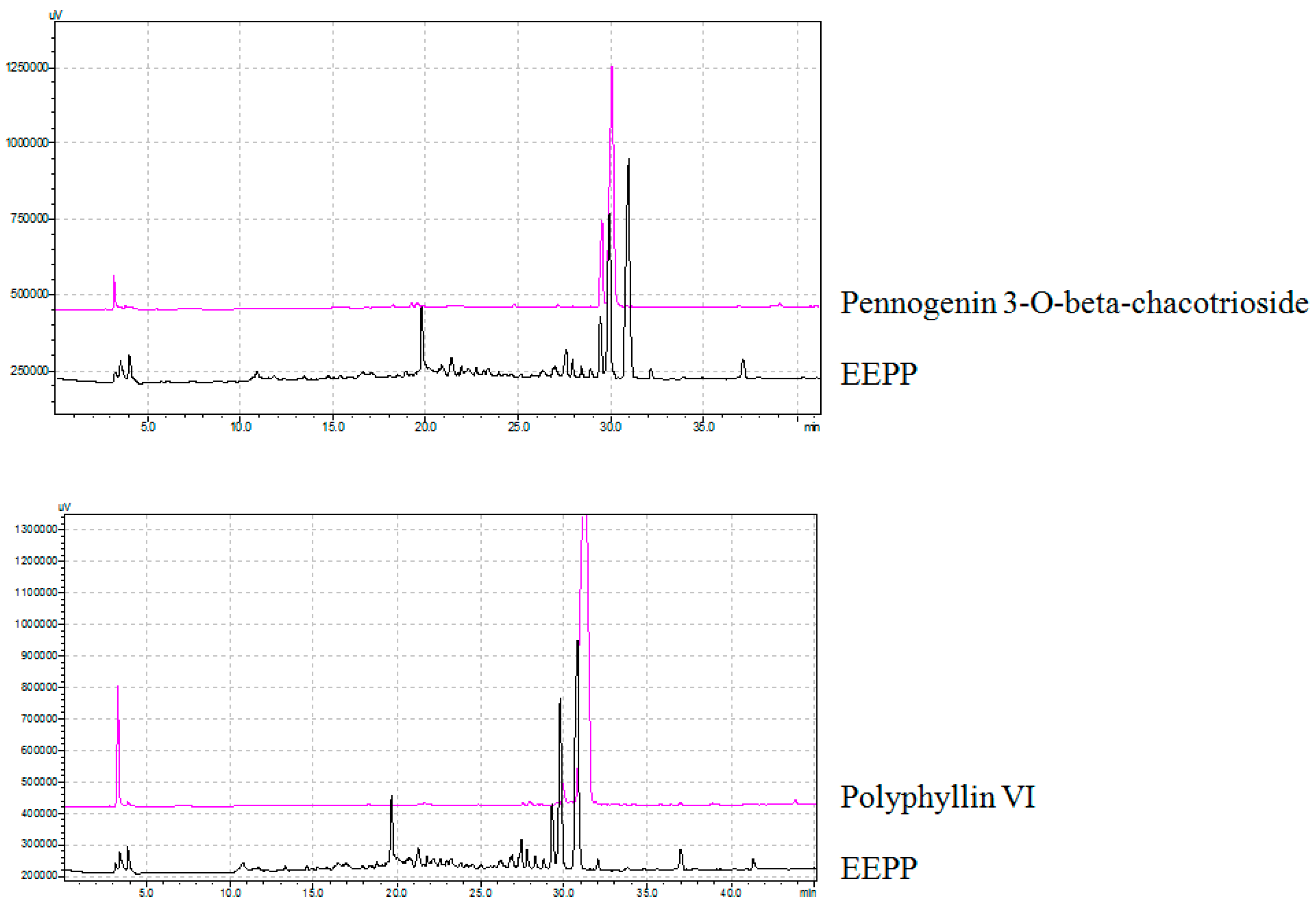
2.4. Isolation and Identification of Active Compounds from EEPP
3. Materials and Methods
3.1. Chemicals
3.2. Cell Culture
3.3. Cell Viability
3.4. Cell Cycle
3.5. DNA Ladder
3.6. Western Blot
3.7. Method of Isolation and Identification of Active Compounds
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EEPP | ethanolic extract of Paris polyphylla |
| LC-3 | light chain-3 |
| TCM | traditional Chinese medicine |
| PCD | programmed cell death |
| Dox | doxorubicin |
References
- Man, S.; Gao, W.; Wei, C.; Liu, C. Anticancer drugs from traditional toxic Chinese medicines. Phytother. Res. 2012, 26, 1449–1465. [Google Scholar] [CrossRef]
- Zhang, C.; Jia, X.; Bao, J.; Chen, S.; Wang, K.; Zhang, Y.; Li, P.; Wan, J.B.; Su, H.; Wang, Y.; et al. Polyphyllin VII induces apoptosis in Hep G2 cells through ROS-mediated mitochondrial dysfunction and MAPK pathways. BMC Compl. Alt. Med. 2016, 16, 58. [Google Scholar] [CrossRef]
- Li, Y.; Gu, J.F.; Zou, X.; Wu, J.; Zhang, M.H.; Jiang, J. The anti-lung cancer activities of steroidal saponins of P. polyphylla Smith var. chinensis (Franch.) Hara through enhanced immunostimulation in experimental Lewis tumor-bearing C57BL/6 mice and induction of apoptosis in the A549 cell line. Molecules 2013, 18, 12916–12936. [Google Scholar] [CrossRef]
- He, H.; Zheng, L.; Sun, Y.P.; Zhang, G.W.; Yue, Z.G. Steroidal saponins from Paris polyphylla suppress adhesion migration and invasion of human lung cancer A549 cells via down-regulating MMP-2 and MMP-9. Asian Pac. J. Cancer Prev. 2014, 15, 10911–10916. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Sun, Y.; Fan, L.; Zhang, F.; Meng, J.; Han, J. Paris saponin VII inhibits growth of colorectal cancer cells through Ras signaling pathway. Biochem. Pharmacol. 2014, 88, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Li, F.R.; Jiao, P.; Yao, S.T.; Sang, H.; Qin, S.C.; Zhang, W.; Zhang, Y.B.; Gao, L.L. Paris polyphylla Smith extract induces apoptosis and activates cancer suppressor gene connexin26 expression. Asian Pac. J. Cancer Prev. 2012, 13, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.W.; Tai, C.J.; Choong, C.Y.; Lin, Y.C.; Lee, B.H.; Shi, Y.C.; Tai, C.J. Aqueous extract of Paris polyphylla (AEPP) inhibits ovarian cancer via suppression of peroxisome proliferator-activated receptor-gamma coactivator (PGC)-1alpha. Molecules 2016, 21, 727. [Google Scholar] [CrossRef]
- Ernst, E.; Cassileth, B.C. The prevalence of complementary/alternative medicine in cancer: A systematic review. Cancer 1998, 83, 777–782. [Google Scholar] [CrossRef]
- Patterson, R.E.; Neuhouser, M.L.; Hedderson, M.M. Types of alternative medicine used by patients with breast, colorectal, or prostate cancer: Predictors, motives, and costs. J. Alternat. Complement. Med. 2002, 8, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.A.; Sanders, T.; Palmer, J.L.; Greisinger, A.; Singletary, S.E. Complementary/alternative medicine use in a comprehensive cancer center and the implications for oncology. J. Clin. Oncol. 2000, 18, 2505–2514. [Google Scholar] [CrossRef]
- Xu, W.; Towers, A.D.; Li, P.; Collet, J.P. Traditional Chinese medicine in cancer care: Perspectives and experiences of patients and professionals in China. Eur. J. Cancer Care 2006, 15, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Qi, F.; Li, A.; Inagaki, Y. Chinese herbal medicines as adjuvant treatment during chemo- or radio-therapy for cancer. Biosci. Trends 2010, 4, 297–307. [Google Scholar] [PubMed]
- Youns, M.; Hoheisel, J.D.; Efferth, T. Traditional Chinese Medicines (TCMs) for molecular targeted therapies of tumours. Curr. Drug Discov. Technol. 2010, 7, 37–45. [Google Scholar] [CrossRef]
- Coleman, M.P.; Quaresma, M.; Berrino, F. Cancer survival in five continents: A worldwide population-based study (CONCORD). Lancet Oncol. 2008, 9, 730–756. [Google Scholar] [CrossRef]
- Cancer Registry Annual Report; Department of Health, Executive Yuan: Taiwan, 2009. Available online: https://www.mohw.gov.tw/lp-137-2.html (accessed on 23 May 2019).
- De Dosso, S.; Sessa, C.; Saletti, P. Adjuvant therapy for colorectal cancer: Present and perspectives. Cancer Treatment Rev. 2009, 35, 160–166. [Google Scholar] [CrossRef]
- Kopetz, S.; Freitas, D.; Calabrich, A.F.C.; Hoff, P.M. Adjuvant chemotherapy for stage II colorectal cancer. Oncology 2008, 22, 260–270. [Google Scholar]
- Ye, M.X.; Zhao, Y.L.; Li, Y.; Miao, Q.; Li, Z.K.; Ren, X.L.; Song, L.Q.; Yin, H.; Zhang, J. Curcumin reverses cis-platin resistance and promotes human lung adenocarcinoma A549/DDP cell apoptosis through HIF-1α and caspase-3 mechanisms. Phytomedicine 2012, 19, 779–787. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Mizushima, N.; Ohsumi, Y.; Yoshimori, T. Autophagosome formation in mammalian cells. Cell Struct. Funct. 2002, 27, 421–429. [Google Scholar] [CrossRef]
- Liu, Y.L.; Yang, P.M.; Shun, C.T.; Wu, M.S.; Weng, J.R.; Chen, C.C. Autophagy potentiates the anti-cancer effects of the histone deacetylase inhibitors in hepatocellular carcinoma. Autophagy 2010, 6, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Nahimana, A.; Attinger, A.; Aubry, D.; Greaney, P.; Ireson, C.; Thougaard, A.V.; Tjørnelund, J.; Dawson, K.M.; Dupuis, M.; Duchosal, M.A. The NAD biosynthesis inhibitor APO866 has potent antitumor activity against hematologic malignancies. Blood 2009, 113, 3276–3286. [Google Scholar] [CrossRef]
- Kim, J.Y.; Cho, T.J.; Woo, B.H.; Choi, K.U.; Lee, C.H.; Ryu, M.H.; Park, H.R. Curcumin-induced autophagy contributes to the decreased survival of oral cancer cells. Arch. Oral Biol. 2012, 57, 1018–1025. [Google Scholar] [CrossRef]
- Tai, C.J.; Wang, C.K.; Tai, C.J.; Lin, Y.F.; Lin, C.S.; Jian, J.Y.; Chang, Y.J.; Chang, C.C. Aqueous extract of Solanum nigrum leaves induces autophagy and enhances cytotoxicity of cisplati, doxorubicin, docetaxel, and 5-fluorouracil in human colorectal carcinoma cells. Evid. Based Complement. Alternat. Med. 2013, 2013, 514719. [Google Scholar] [PubMed]
- Cummings, J.; Smyth, J.F. DNA topoisomerase I and II as targets for rational design of new anticancer drugs. Ann. Oncol. 1993, 4, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Lee, B.H.; Pan, T.M. Red mold dioscorea-induced G2/M arrest and apoptosis in human oral cancer cells. J. Sci. Food Agric. 2010, 90, 2709–2715. [Google Scholar] [CrossRef] [PubMed]
- Bushell, M.; Poncet, D.; Marissen, W.E.; Flotow, H.; Lloyd, R.E.; Clemens, M.J.; Morley, S.J. Cleavage of polypeptide chain initiation fctor eIF4GI during apoptosis in lymphoma cells: Characterization of an internal fragment generated by caspase-3-mediated cleavage. Cell. Death Differ. 2000, 7, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Lee, B.H.; Huang, Y.C.; Hsu, Y.W.; Pan, T.M. Ankaflavin, a novel Nrf-2 activator for attenuating allergic airway inflammation. Free Radic. Biol. Med. 2012, 53, 1643–1651. [Google Scholar] [CrossRef]
Sample Availability: Not available. |


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.-T.; Uen, W.-C.; Choong, C.-Y.; Shi, Y.-C.; Lee, B.-H.; Tai, C.-J.; Tai, C.-J. Paris Polyphylla Inhibits Colorectal Cancer Cells via Inducing Autophagy and Enhancing the Efficacy of Chemotherapeutic Drug Doxorubicin. Molecules 2019, 24, 2102. https://doi.org/10.3390/molecules24112102
Lin L-T, Uen W-C, Choong C-Y, Shi Y-C, Lee B-H, Tai C-J, Tai C-J. Paris Polyphylla Inhibits Colorectal Cancer Cells via Inducing Autophagy and Enhancing the Efficacy of Chemotherapeutic Drug Doxorubicin. Molecules. 2019; 24(11):2102. https://doi.org/10.3390/molecules24112102
Chicago/Turabian StyleLin, Liang-Tzung, Wu-Ching Uen, Chen-Yen Choong, Yeu-Ching Shi, Bao-Hong Lee, Cheng-Jeng Tai, and Chen-Jei Tai. 2019. "Paris Polyphylla Inhibits Colorectal Cancer Cells via Inducing Autophagy and Enhancing the Efficacy of Chemotherapeutic Drug Doxorubicin" Molecules 24, no. 11: 2102. https://doi.org/10.3390/molecules24112102
APA StyleLin, L.-T., Uen, W.-C., Choong, C.-Y., Shi, Y.-C., Lee, B.-H., Tai, C.-J., & Tai, C.-J. (2019). Paris Polyphylla Inhibits Colorectal Cancer Cells via Inducing Autophagy and Enhancing the Efficacy of Chemotherapeutic Drug Doxorubicin. Molecules, 24(11), 2102. https://doi.org/10.3390/molecules24112102