Spermidine Enhances Heat Tolerance of Rice Seeds by Modulating Endogenous Starch and Polyamine Metabolism

Abstract
:1. Introduction
2. Results
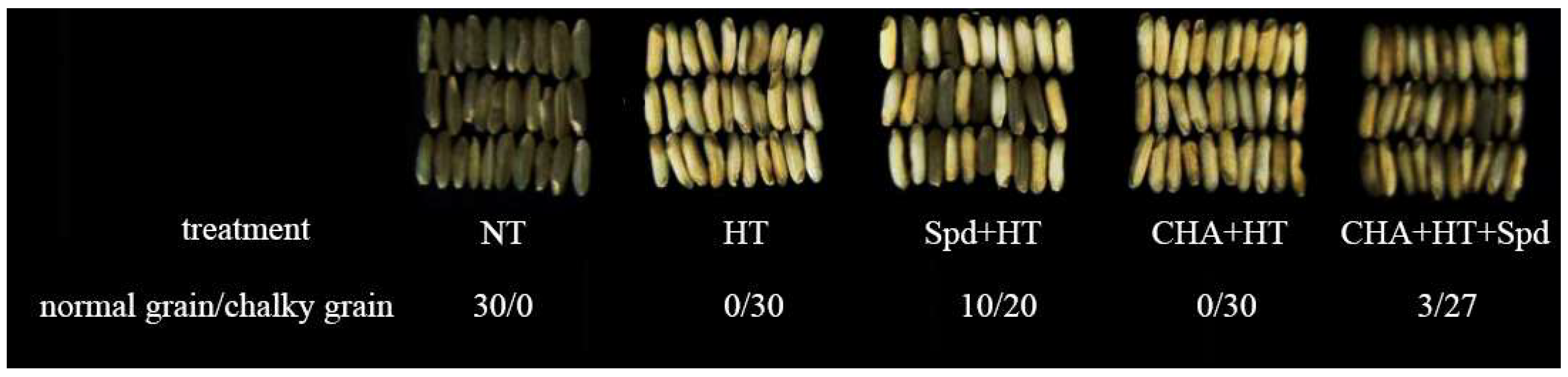
2.1. Effects of Exogenous Spermidine on Rice Seed Quality under High Temperature Stress during the Early Developmental Stage of Seeds
2.2. Exogenous Spd Could Increase Rice Seed Germination and Seedling Growth under High Temperature Stress
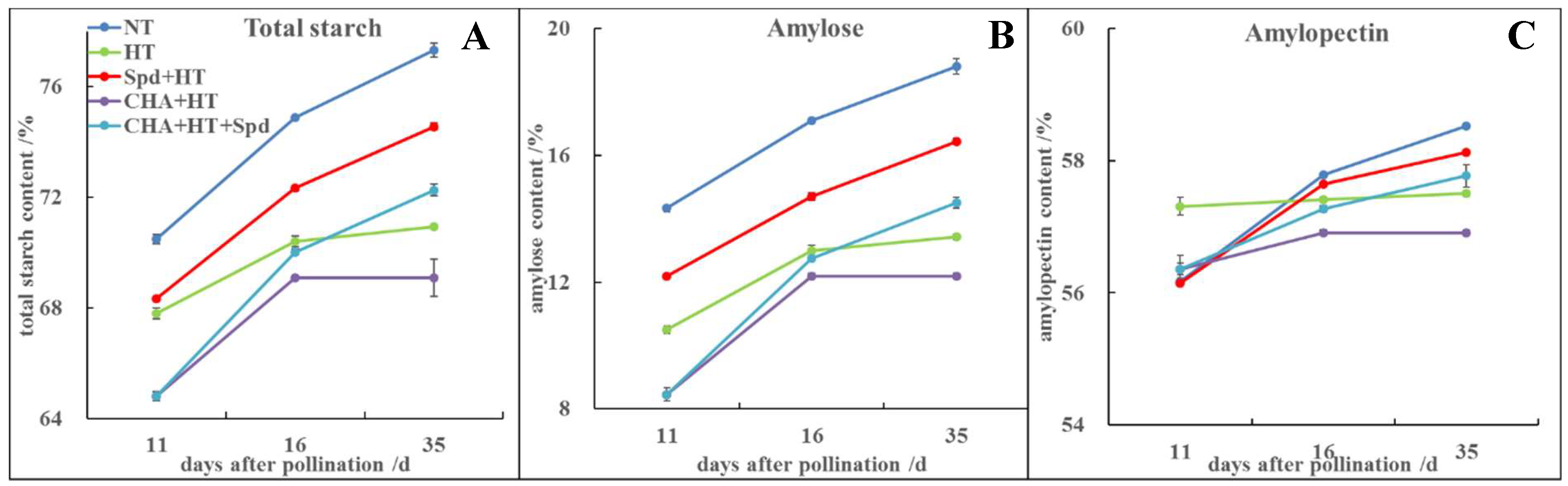
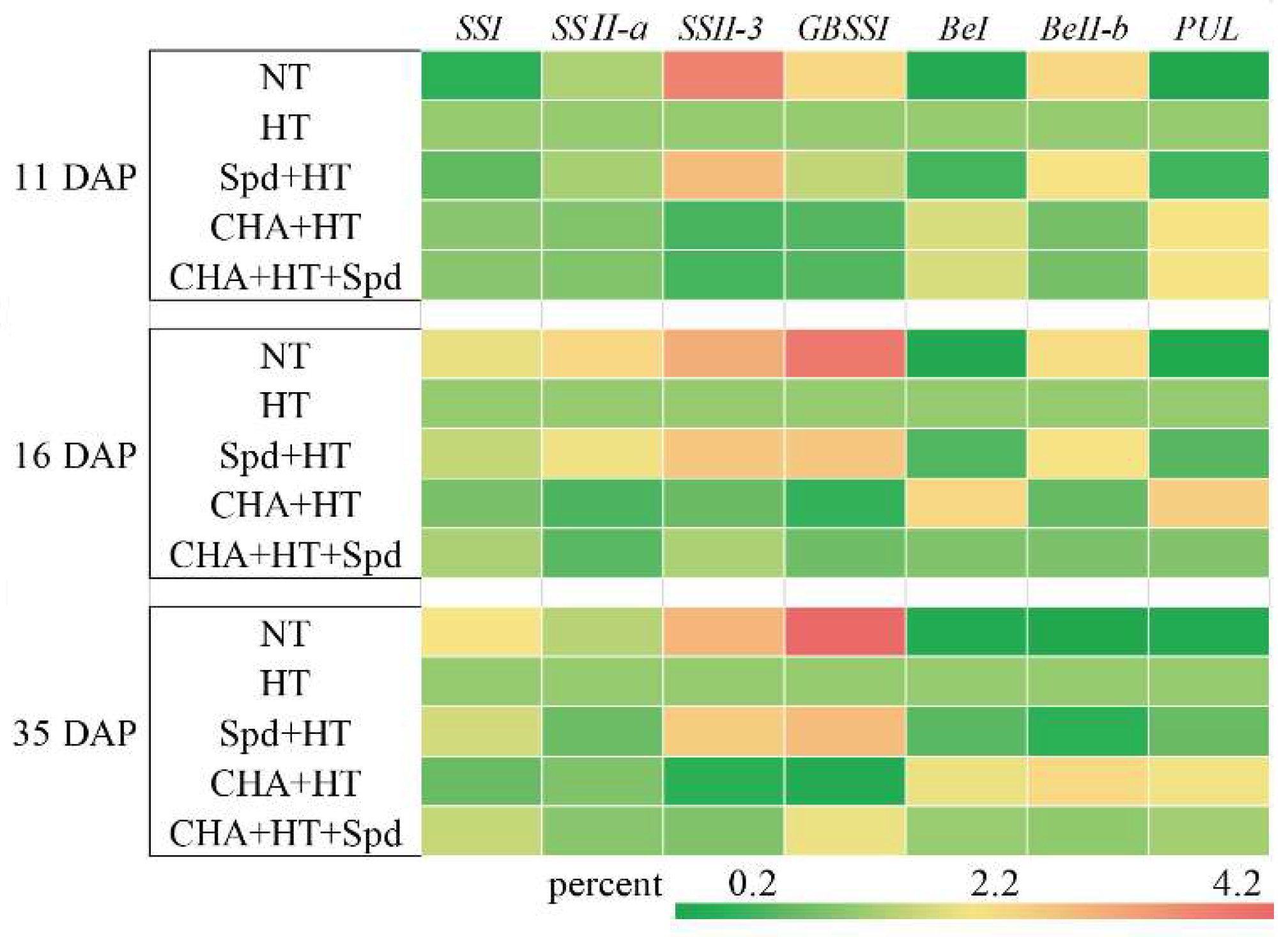
2.3. Exogenous Spd Could Increase the Starch Content in Rice Seed under High Temperature Stress
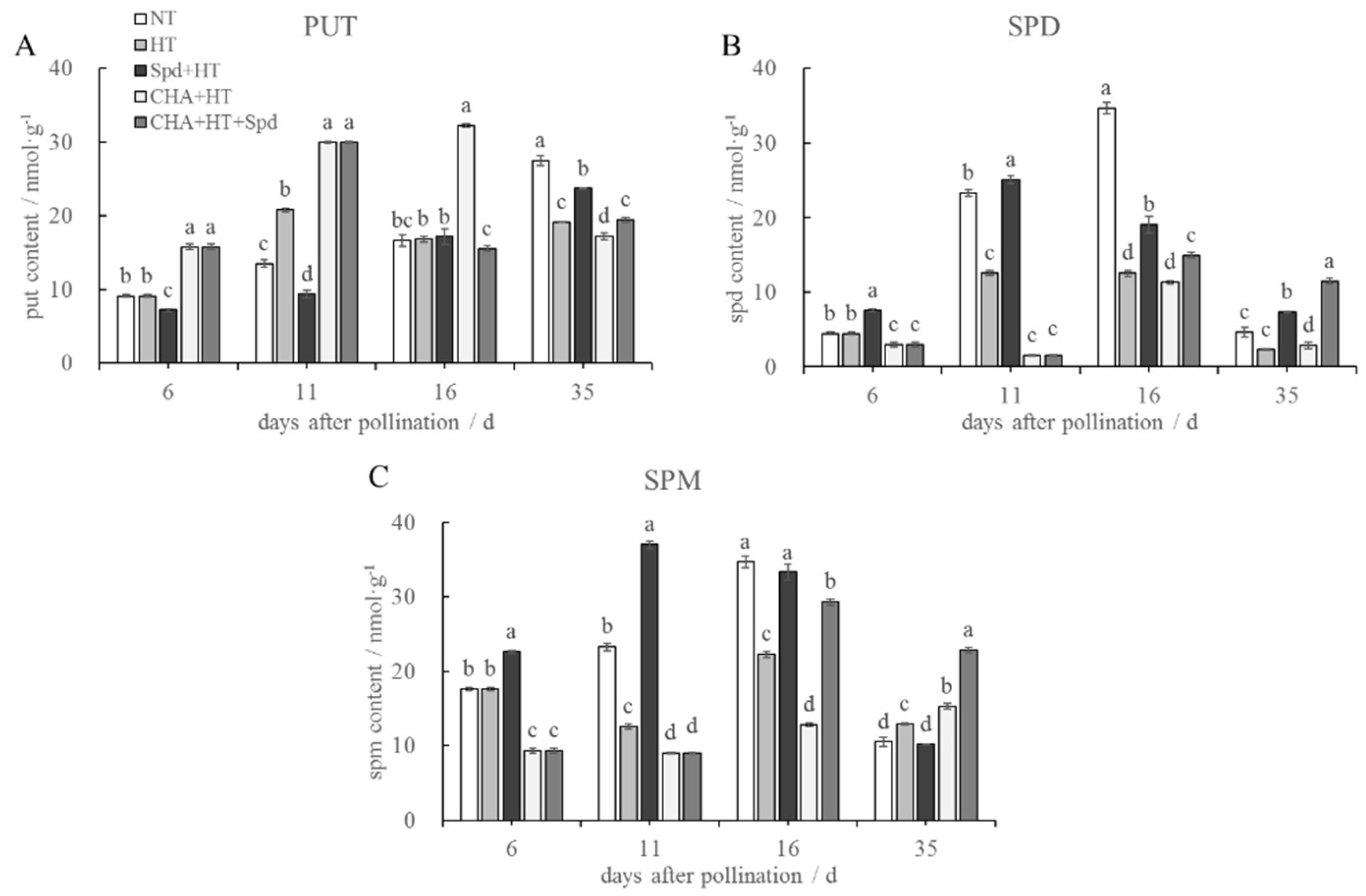
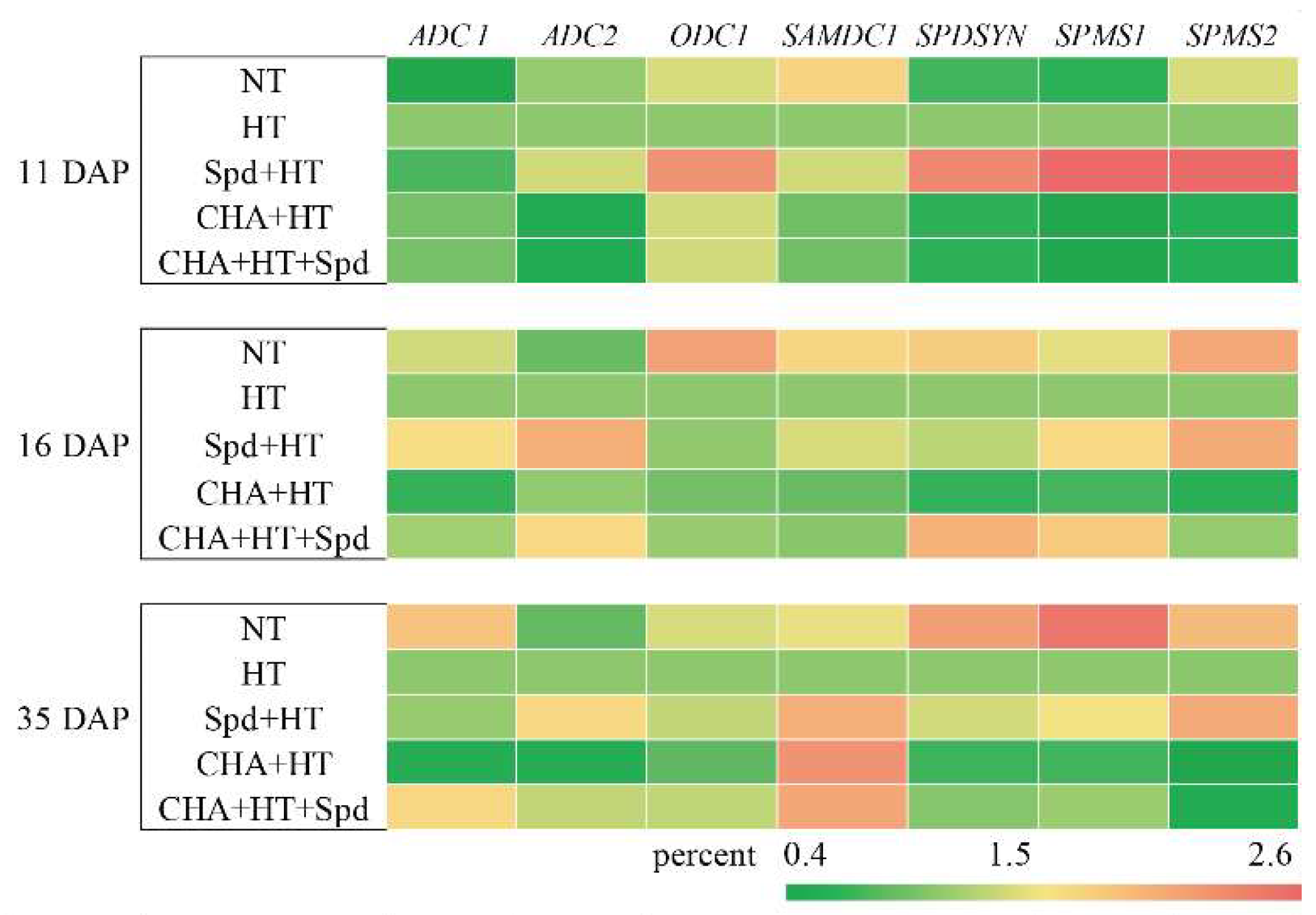
2.4. Exogenous Spd Treatment Had a Positive Effect on the Levels of Endogenous Spd and Spm under High-Temperature Stress
2.5. Exogenous Spd Could Promote the peroxidase (POD) Activity and Decrease MDA Content under High Temperature Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Cultivation Conditions
4.2. Measurement of Morphological Index of Rice Grain
4.3. Germination Test
4.4. Measurement of Physiological Parameters
4.5. Measurement of Starch Content
4.6. Measurement of Endogenous Polyamine Content
4.7. Real-time Fluorescence Quantitative PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carmo-Silva, A.E.; Gore, M.A.; Andrade-Sanchez, P.; French, A.N.; Hunsaker, D.J.; Salvucci, M.E. Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field. J. Exp. Bot. 2012, 83, 1–11. [Google Scholar]
- Siddique, K.H.M.J. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148–1167. [Google Scholar]
- Deeba, F.; Pandey, A.K.; Ranjan, S.; Mishra, A.; Singh, R.; Sharma, Y.K.; Shirke, P.A. Physiological and proteomic responses of cotton (Gossypium herbaceum L.) to drought stress. Plant Physiol. Biochem. 2012, 53, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, M.; An, N.; Liang, L.; Wan, Y.; Jin, W.J. A novel reference plasmid for the qualitative detection of genetically modified rice in food and feed. BioMed Res. Int. 2015, 2015, 948297. [Google Scholar] [CrossRef]
- Asaoka, M.; Okuno, K.; Sugimoto, Y.; Kawakami, J.; Fuwa, H.J. Effect of environmental temperature during development of rice plants on some properties of endosperm starch. Starch-Starke 2010, 36, 189–193. [Google Scholar]
- Inouchi, N.; Ando, H.; Asaoka, M.; Okuno, K.; Fuwa, H.J. The effect of environmental temperature on distribution of unit chains of rice amylopectin. Starch-Starke 2000, 52, 8–12. [Google Scholar] [CrossRef]
- Krishnan, P.; Ramakrishnan, B.J. Chapter three—High-temperature effects on rice growth, yield, and grain quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar]
- Peter, G.J. Regulation of starch biosynthesis in response to a fluctuating environment. Plant Physiol. 2011, 155, 1566–1577. [Google Scholar]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. J. Plant Physiol. 2001, 127, 459. [Google Scholar] [CrossRef]
- Tanaka, N.; Fujita, N.; Nishi, A.; Satoh, H.; Hosaka, Y.; Ugaki, M.; Kawasaki, S.; Nakamura, Y.J. The structure of starch can be manipulated by changing the expression levels of starch branching enzyme IIb in rice endosperm. Plant Biol. J. 2010, 2, 507–516. [Google Scholar] [CrossRef]
- Tang, D.; Wang, Y.; Cai, J.; Zhao, R.J. Effects of exogenous application of plant growth regulators on the development of ovule and subsequent embryo rescue of stenospermic grape (Vitis vinifera L.). Sci. Hortic. 2009, 120, 51–57. [Google Scholar] [CrossRef]
- Zapata, P.J.; Serrano, M.A.; Pretel, M.T.; Amorós, A.; Botella, M.Á.J. Polyamines and ethylene changes during germination of different plant species under salinity. Plant Sci. 2004, 167, 781–788. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Bogatek, R.; Regulation, A.G. Polyamines and nitric oxide link in regulation of dormancy removal and germination of apple (Malus domestica borkh.) embryos. J. Plant Growth Regul. 2014, 33, 590–601. [Google Scholar] [CrossRef]
- Yiu, J.C.; Liu, C.W.; Fang, Y.T.; Lai, Y.S.J. Waterlogging tolerance of welsh onion (Allium fistulosum L.) Enhanced by exogenous spermidine and spermine. Plant Physiol. Biochem. 2009, 47, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lin, C.; He, F.; Li, Z.; Guan, Y.; Hu, Q.; Hu, J. Exogenous spermidine improves seed germination of sweet corn via involvement in phytohormone interactions, H2O2 and relevant gene expression. Bmc Plant Biol. 2017, 17, 1. [Google Scholar] [CrossRef]
- Sagor, G.H.M.; Berberich, T.; Takahashi, Y.; Niitsu, M.; Kusano, T.J. The polyamine spermine protects Arabidopsis from heat stress-induced damage by increasing expression of heat shock-related genes. Transgenic Res. 2013, 22, 595–605. [Google Scholar] [CrossRef]
- Alcázar, R.; Cuevas, J.C.; Planas, J.; Zarza, X.; Bortolotti, C.; Carrasco, P.; Salinas, J.; Tiburcio, A.F.; Science, T.A. Integration of polyamines in the cold acclimation response. Plant Sci. 2011, 180, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.H.; Nakajima, I.; Moriguchi, T. Effects of salt and osmotic stresses on free polyamine content and expression of polyamine biosynthetic genes in Vitis vinifera. Biol. Plant. 2011, 55, 340–344. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Yoshida, N.; Fujita, M.J. Spermidine pretreatment enhances heat tolerance in rice seedlings through modulating antioxidative and glyoxalase systems. Plant Growth Regul. 2014, 73, 31–44. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, T.; Amombo, E.; Wang, G.; Xie, Y.; Fu, J. The alleviation of heat damage to photosystem ii and enzymatic antioxidants by exogenous spermidine in tall fescue. Front. Plant Sci. 2017, 8, 1747. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xu, Y.; Wang, J.; Wang, Z.; Yang, J.; Zhang, J. Polyamines and ethylene interact in rice grains in response to soil drying during grain filling. J. Exp. Bot. 2013, 64, 2523. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Chen, Y.H.; Chen, M.X.; Wang, Z.Q.; Wu, C.F.; Bian, X.C.; Yang, J.C.; Zhang, J.H. Growth characteristics and endosperm structure of superior and inferior spikelets of indica rice under high-temperature stress. Biol. Plant. 2016, 60, 1–11. [Google Scholar] [CrossRef]
- Saleethong, P.; Sanitchon, J.; Kongngern, K.; Theerakulpisut, P. Effects of exogenous spermidine (Spd) on yield, yield-related parameters and mineral composition of rice (Oryza sativa L. ssp. indica) grains under salt stress. Aust. J. Crop Sci. 2013, 7, 1293–1301. [Google Scholar]
- Duan, H.; Yang, J.C. Research advances in the effect of high temperature on rice and its mechanism. Chin. J. Rice Sci. 2012, 26, 393–400. [Google Scholar]
- Liu, Q.; Wu, X.; Ma, J.; Li, T.; Zhou, X.; Guo, T. Effects of high air temperature on rice grain quality and yield under field condition. Agron. J. 2013, 105, 446. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef] [PubMed]
- Morita, S.; Yonemaru, J.; Takanashi, J. Grain Growth and Endosperm Cell Size Under High Night Temperatures in Rice (Oryza sativa L.). Ann. Bot. 2005, 95, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.; Zhang, H.; Li, L.; Liu, X.; Chen, L.; Chen, W.; Ding, Y. Exogenous spermidine enhances the photosynthetic and antioxidant capacity of rice under heat stress during early grain-filling period. Funct Plant Biol. 2018, 45, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, F.; Shahzad, M.; Basra, H. Seed priming with polyamines improves the germination and early seedling growth in fine rice. J. New Seeds 2008, 9, 145–155. [Google Scholar]
- Cui, K.; Wang, H.Y.; Li, K.; Liao, S.X.; Li, L.; Zhang, C. Physiological and biochemical effects of ultra-dry storage on Barbados nut seeds. Crop Sci. 2014, 54, 1748. [Google Scholar] [CrossRef]
- Diao, Q.; Song, Y.; Qi, H. Exogenous spermidine enhances chilling tolerance of tomato (Solanum lycopersicum L.) seedlings via involvement in polyamines metabolism and physiological parameter levels. Acta Physiol. Plant. 2015, 37, 1–15. [Google Scholar] [CrossRef]
- Aryadeep, R.; Supratim, B.; Sengupta, D.N. Amelioration of salinity stress by exogenously applied spermidine or spermine in three varieties of indica rice differing in their level of salt tolerance. J. Plant Physiol. 2011, 168, 317–328. [Google Scholar]
- Huang, H.; Villanueva, V.R. Inhibition of polyamine biosynthesis and seed germination in Picea abies. Phytochemistry 1992, 31, 3353–3356. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Ghosh, B. Protein synthesis and loss of viability of rice seeds: Effect of polyamines on in vitro translation. Physiol. Plant. 2010, 68, 441–445. [Google Scholar] [CrossRef]
- Bueno, M.; Garrido, D.; Matilla, A. Gene expression induced by spermine in isolated embryonic axes of chickpea seeds. Physiol. Plant. 2010, 87, 381–388. [Google Scholar] [CrossRef]
- Cao, D.D.; Hu, J.; Zhu, S.J.; Hu, W.M.; Knapp, A. Relationship between changes in endogenous polyamines and seed quality during development of sh2 sweet corn (Zea mays L.) seed. Sci. Hortic. 2010, 123, 301–307. [Google Scholar] [CrossRef]
- Duan, J.; Li, J.; Guo, S.; Kang, Y. Exogenous spermidine affects polyamine metabolism in salinity-stressed Cucumis sativus roots and enhances short-term salinity tolerance. J. Plant Physiol. 2008, 165, 1620–1635. [Google Scholar] [CrossRef]
- Jimenez-Bremont, J.; Ruiz, O.; Rodriguez-Kessler, M. Modulation of spermidine and spermine levels in maize seedlings subjected to long-term salt stress. Plant Physiol. Biochem. 2007, 45, 812–821. [Google Scholar] [CrossRef]
- Yoshihisa, K.; Lixiong, H.; Kazuyoshi, N.; Shuhei, M.; Izumi, I.; Shoji, T.J.P. Overexpression of spermidine synthase enhances tolerance to multiple environmental stresses and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 712. [Google Scholar]
- Sarvajeet Singh, G.; Narendra, T. Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [Green Version]
- Yang, W.B.; Yong, L.I.; Yin, Y.P.; Jiang, W.W.; Peng, D.L.; Cui, Z.Y.; Yang, D.Q.; Wang, Z.L. Ethylene and spermidine in wheat grains in relation to starch content and granule size distribution under water deifcit. J. Agric. Sci. 2014, 13, 2141–2153. [Google Scholar]
- Wang, Z.; Xu, Y.; Wang, J.; Yang, J. Polyamine and ethylene interactions in grain filling of superior and inferior spikelets of rice. Plant Growth Regul. 2012, 66, 215–228. [Google Scholar] [CrossRef]
- Zhang, X. Preliminary Study on the Effects of Starch, Protein and Fat Content on the Vigor of Hybrid Rice Seeds. Master’s Thesis, Zhejiang Agriculture and Forestry University, Hangzhou, China, 2014. [Google Scholar]
- Fan, L. Studies on low seed vigor of sh2 sweet corn and seed treatments for improving it’s field seedling emergence. Acta Agron. Sin. 1998, 1, 103–109. [Google Scholar]
- Nicholas, D.; Chua, H.P.; Rosniyana, A.; Hazila, K.K. Effects of aging on physico-chemical properties, nutritional compositions and cooking characteristics of Bario rice. J. Trop. For. Sci. 2013, 41, 239–248. [Google Scholar]
- Hu, Q.; Fu, Y.; Guan, Y.; Lin, C.; Cao, D.; Hu, W.; Sheteiwy, M. Inhibitory effect of chemical combinations on seed germination and pre-harvest sprouting in hybrid rice. J. Plant Growth Regul. 2016, 80, 281–289. [Google Scholar] [CrossRef]
- Zhu, L.W.; Cao, D.D.; Hu, Q.J.; Guan, Y.J.; Hu, W.M.; Nawaz, A.; Hu, J. Soluble protein, sugar, GA3 and sHSPs genes related with seed germination during maturation of hybrid rice seed. J. Sci. Food Agric. 2015, 96, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q. Determination of amylose and amylopectin in micro-pore starch of grain by dual-wavelength spectrophotometry. Chin. J. Spectrosc. Lab. 2013, 30, 1569–1572. [Google Scholar]
Sample Availability: Not available. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | DAP | SH (cm) | SDW (g) | GP | GI | VI |
|---|---|---|---|---|---|---|
| NT | 11 | 5.04 ± 0.03b | 0.028 ± 0.001b | 0.42 ± 0.012b | 2.63 ± 0.27b | 0.07 ± 0.007c |
| HT | 5.91 ± 0.05a | 0.036 ± 0.001a | 0.50 ± 0.012a | 3.40 ± 0.17a | 0.12 ± 0.007a | |
| Spd + HT | 4.43 ± 0.05c | 0.032 ± 0.001b | 0.42 ± 0.012a | 2.91 ± 0.03ab | 0.09 ± 0.002b | |
| CHA + HT | 2.83 ± 0.04d | 0.019 ± 0.001c | 0.34 ± 0.010c | 1.75 ± 0.15c | 0.03 ± 0.003d | |
| CHA + HT + Spd | 2.83 ± 0.04d | 0.019 ± 0.001c | 0.34 ± 0.010c | 1.82 ± 0.11c | 0.03 ± 0.003d | |
| NT | 16 | 8.12 ± 0.06a | 0.053 ± 0.001a | 0.67 ± 0.093ab | 4.52 ± 0.58a | 0.24 ± 0.028a |
| HT | 5.70 ± 0.08d | 0.040 ± 0.001c | 0.62 ± 0.012b | 3.64 ± 0.06bc | 0.15 ± 0.002b | |
| Spd + HT | 7.25 ± 0.06b | 0.049 ± 0.001b | 0.76 ± 0.012a | 4.21 ± 0.05ab | 0.21 ± 0.003a | |
| CHA + HT | 4.57 ± 0.06e | 0.031 ± 0.001e | 0.55 ± 0.017b | 2.54 ± 0.16d | 0.08 ± 0.004c | |
| CHA + HT + Spd | 6.60 ± 0.15c | 0.046 ± 0.001d | 0.65 ± 0.007ab | 3.27 ± 0.05cd | 0.15 ± 0.004b | |
| NT | 35 | 8.89 ± 0.08a | 0.100 ± 0.001a | 0.85 ± 0.018a | 6.45 ± 0.32a | 0.64 ± 0.031a |
| HT | 5.90 ± 0.01c | 0.052 ± 0.002c | 0.59 ± 0.007c | 3.82 ± 0.09c | 0.20 ± 0.011c | |
| Spd + HT | 7.93 ± 0.04b | 0.063 ± 0.001b | 0.87 ± 0.007a | 6.06 ± 0.12a | 0.38 ± 0.006b | |
| CHA + HT | 5.48 ± 0.33c | 0.044 ± 0.001d | 0.43 ± 0.007d | 2.48 ± 0.15d | 0.11 ± 0.008d | |
| CHA + HT + Spd | 7.72 ± 0.10b | 0.054 ± 0.001c | 0.63 ± 0.013b | 3.85 ± 0.10b | 0.21 ± 0.006c |
| DAP | 3–5 (Spd or CHA Sprayed) | 6 | 7–11 (High Temperature Treatment) | 11 | 12–15 (Spd Sprayed Based on CHA + HT) | 16 | 28 | 35 | |
|---|---|---|---|---|---|---|---|---|---|
| Treatment | |||||||||
| NT | -- | √ | -- | √ | -- | √ | √ | √ | |
| HT | -- | -- | -- | √ | -- | √ | √ | √ | |
| Spd + HT | -- | √ | -- | √ | -- | √ | √ | √ | |
| CHA + HT | -- | √ | -- | √ | -- | √ | √ | √ | |
| CHA + HT + Spd | -- | -- | -- | -- | -- | √ | √ | √ | |
| Locus ID | Gene | Primer Sequence | |
|---|---|---|---|
| LOC_Os06g04070 | ADC1 | FW | CGTCATCGACGTTGGTGGA |
| RW | CCAAGCTGTATGCCACGGAC | ||
| LOC_Os04g01690 | ADC2 | FW | AGAAGGTTGCGACGGAGAATG |
| RW | TGGTCAGCCCTTTCTTCATCA | ||
| LOC_Os09g37120 | ODC1 | FW | CGGCTGGCTCCAACTTCAA |
| RW | TGGAGTATGCCAGGTGGATCTT | ||
| LOC_Os04g42095 | SAMDC1 | FW | GTCTTTGCTGACCCTGATGG |
| RW | CGTGCAAGATCCAGAACAGAG | ||
| LOC_Os07g22600 | SPDSYN | FW | GGTGTTTCAGTCCTCCACGTA |
| RW | TCCCTCTCAGTGACCTGAATC | ||
| LOC_Os06g33710 | SPMS1 | FW | CCTGGTGGAGTTCTATGC |
| RW | CACTGCTGGACCTTCTTT | ||
| LOC Os02g15550 | SPMS2 | FW | AGAGCATGTGGTTGCATACGC |
| RW | AACCCTTGAATGTCTCACGGC | ||
| LOC_Os06g04200 | GBSSI | FW | ACCTGACACTGGAGTTGATTAC |
| RW | GTATGGGTTGTTGTTGAGGTTTAG | ||
| LOC_Os06g06560 | SSI | FW | GTCTTGTGCCAGTCCTTCTT |
| RW | CACACCCTGATGTGCTAGATTAT | ||
| LOC_Os02g51070 | SSII-2 | FW | CAGGGCCAAATGTGATGAATG |
| RW | CTCTTCTTGCCAGAGCCTTAG | ||
| LOC_Os06g12450 | SSII-3 | FW | CTGCACTCCTGCCTGTTTAT |
| RW | GCCCTGGTAAGCGATATTATGT | ||
| LOC_Os06g51084 | Be | FW | AATGGGCATGCATCGACATC |
| RW | CTGGTTCTTGCCCTTCCCTA | ||
| LOC_Os02g0528200 | BEIIb | FW | GGCATGCTAGAGTTTGACCG |
| RW | TCCACCAAAGAGTCCAGCAT | ||
| LOC_Os04g0164900 | PUL | FW | GGTCTGTTCTTGGAGCCTAAT |
| RW | CAGCTAGTCCGATCTGTATGTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Gu, Q.; Dong, Q.; Zhang, Z.; Lin, C.; Hu, W.; Pan, R.; Guan, Y.; Hu, J. Spermidine Enhances Heat Tolerance of Rice Seeds by Modulating Endogenous Starch and Polyamine Metabolism. Molecules 2019, 24, 1395. https://doi.org/10.3390/molecules24071395
Fu Y, Gu Q, Dong Q, Zhang Z, Lin C, Hu W, Pan R, Guan Y, Hu J. Spermidine Enhances Heat Tolerance of Rice Seeds by Modulating Endogenous Starch and Polyamine Metabolism. Molecules. 2019; 24(7):1395. https://doi.org/10.3390/molecules24071395
Chicago/Turabian StyleFu, Yuying, Qingqing Gu, Qian Dong, Zhihao Zhang, Cheng Lin, Weimin Hu, Ronghui Pan, Yajing Guan, and Jin Hu. 2019. "Spermidine Enhances Heat Tolerance of Rice Seeds by Modulating Endogenous Starch and Polyamine Metabolism" Molecules 24, no. 7: 1395. https://doi.org/10.3390/molecules24071395
APA StyleFu, Y., Gu, Q., Dong, Q., Zhang, Z., Lin, C., Hu, W., Pan, R., Guan, Y., & Hu, J. (2019). Spermidine Enhances Heat Tolerance of Rice Seeds by Modulating Endogenous Starch and Polyamine Metabolism. Molecules, 24(7), 1395. https://doi.org/10.3390/molecules24071395