Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
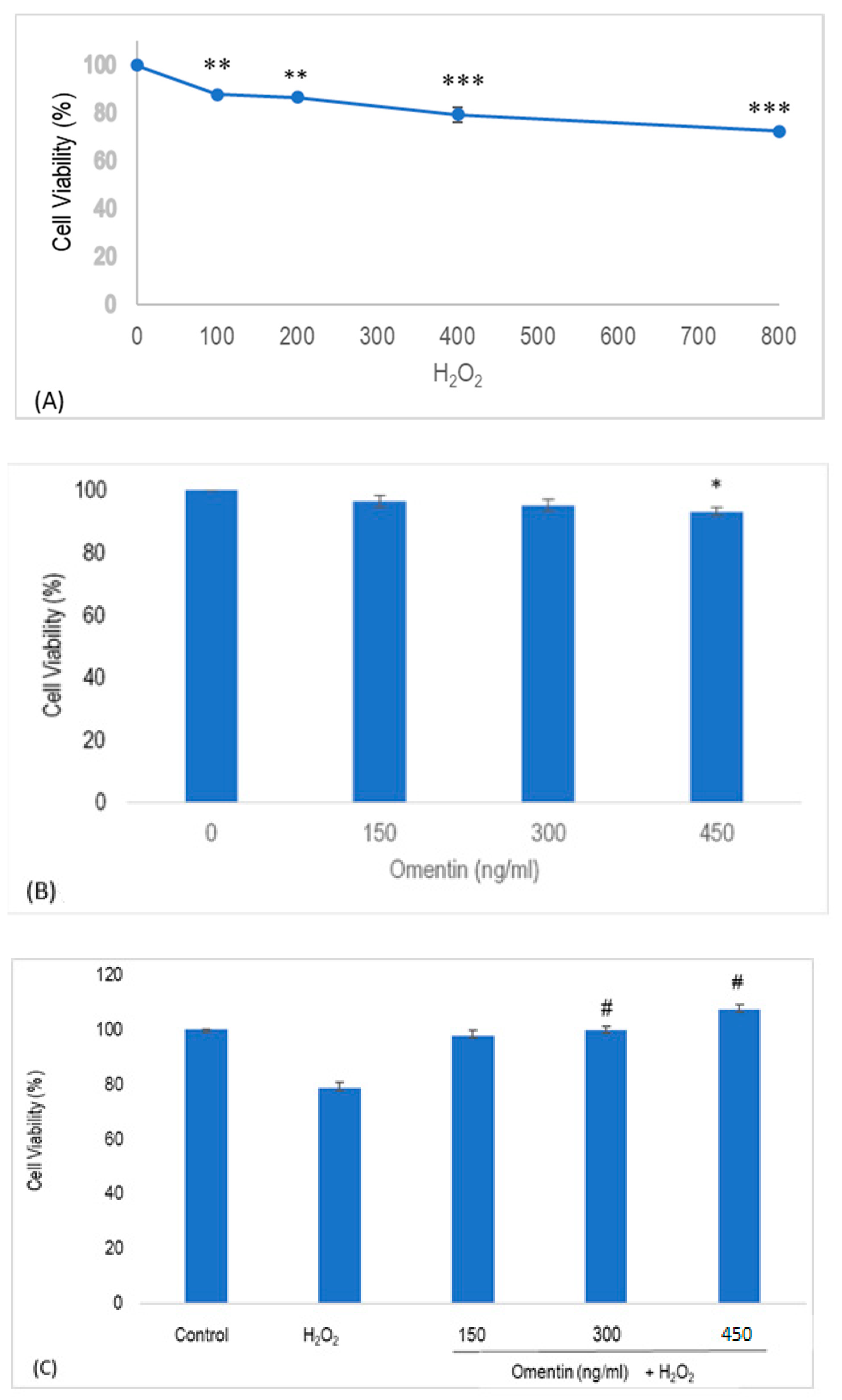
2.1. Effect of Omentin and H2O2 in the Viability of Human Umbilical Vein Endothelial Cells (HUVECs)
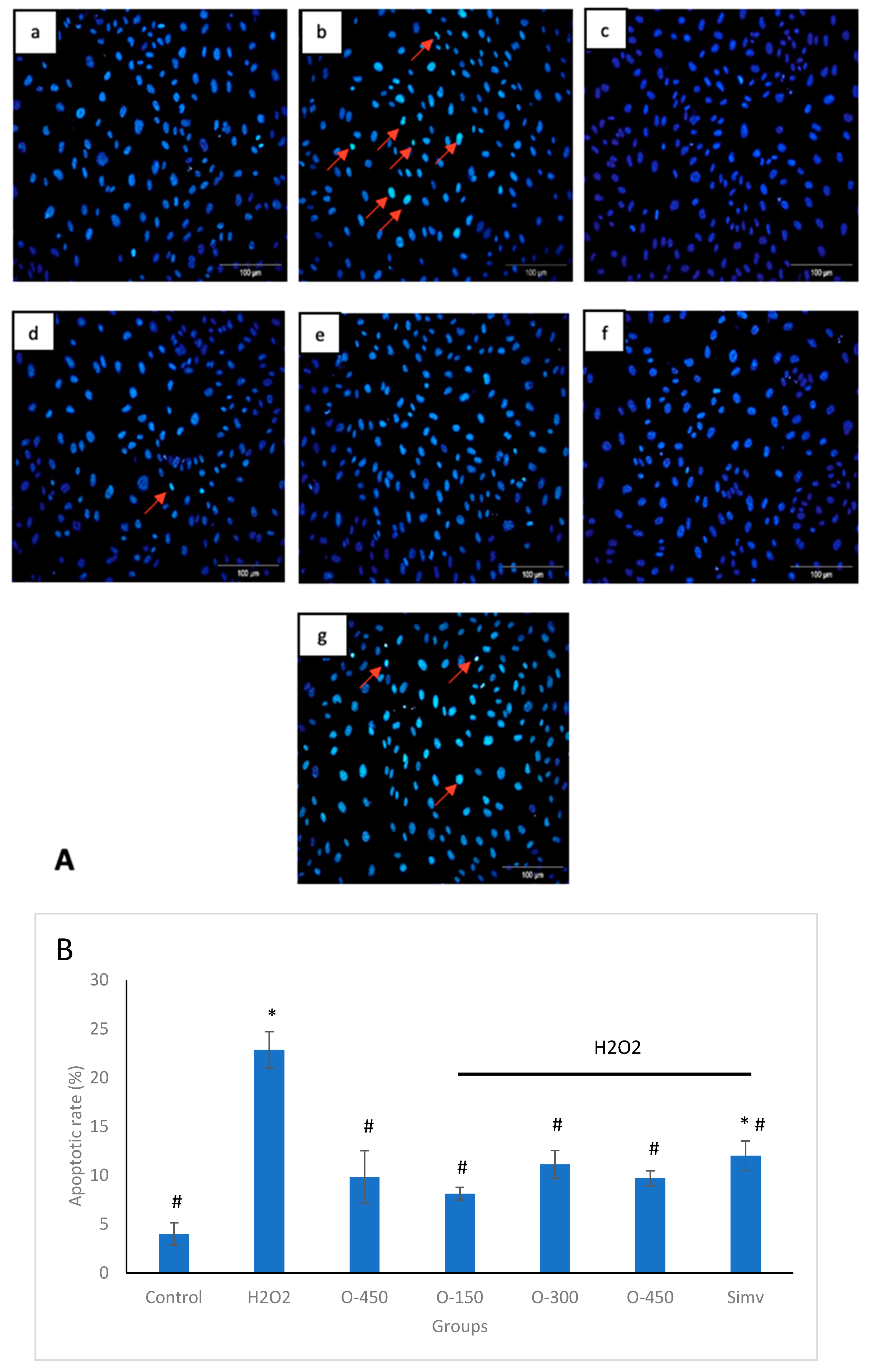
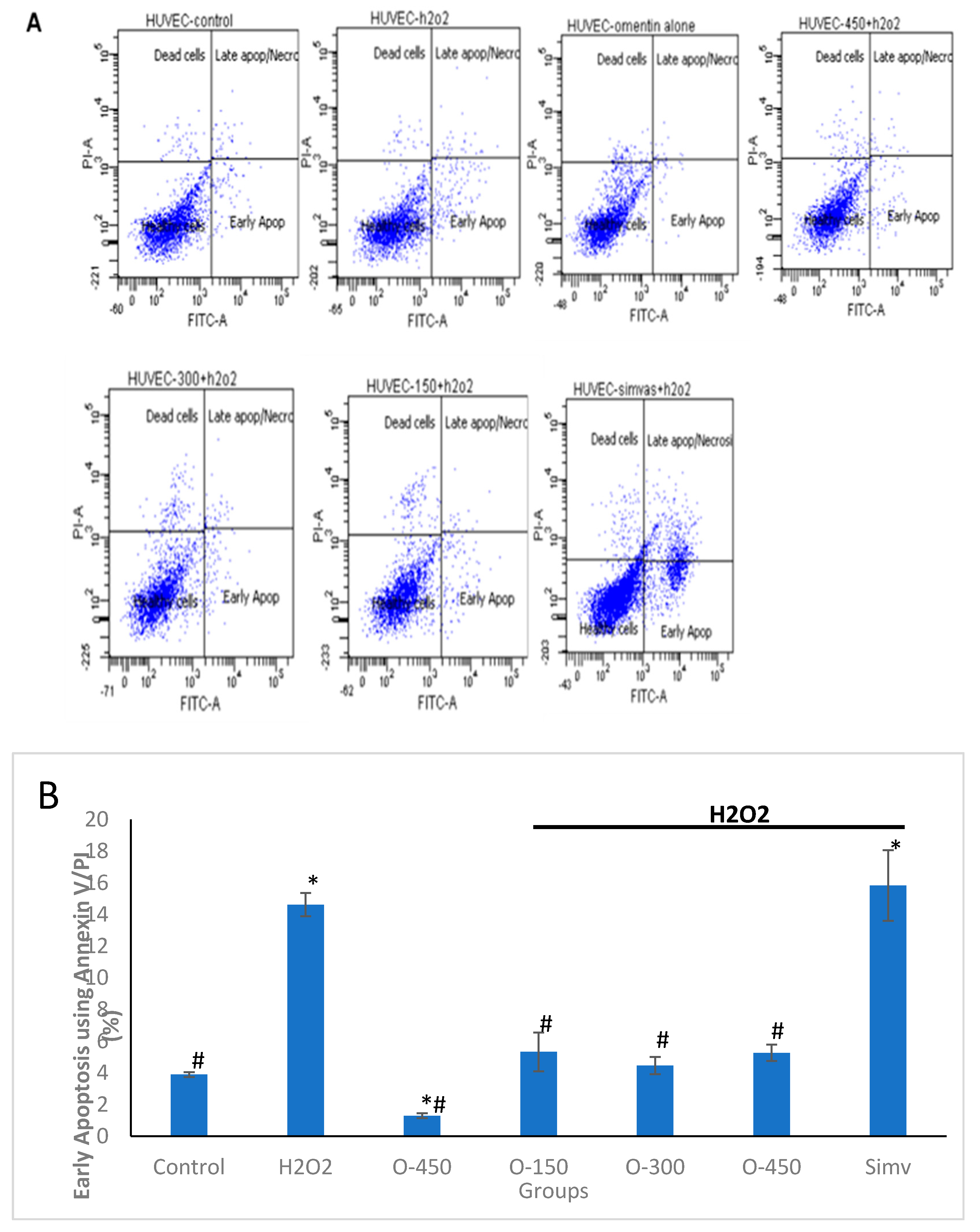
2.2. Effect of Omentin on H2O2-Induced Apoptosis in HUVECs
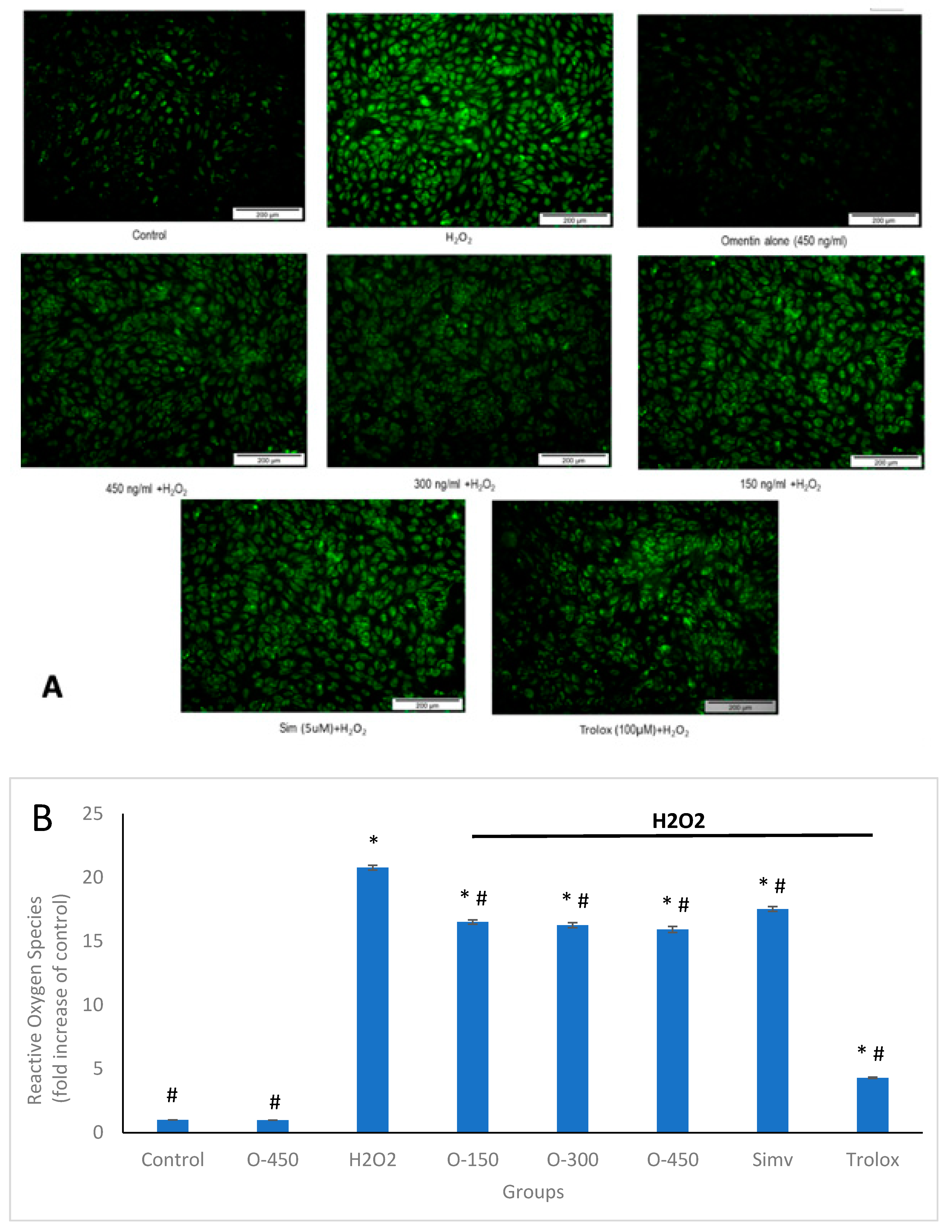
2.3. Antioxidant Effect of Omentin in H2O2-Induced ROS Production in HUVECs
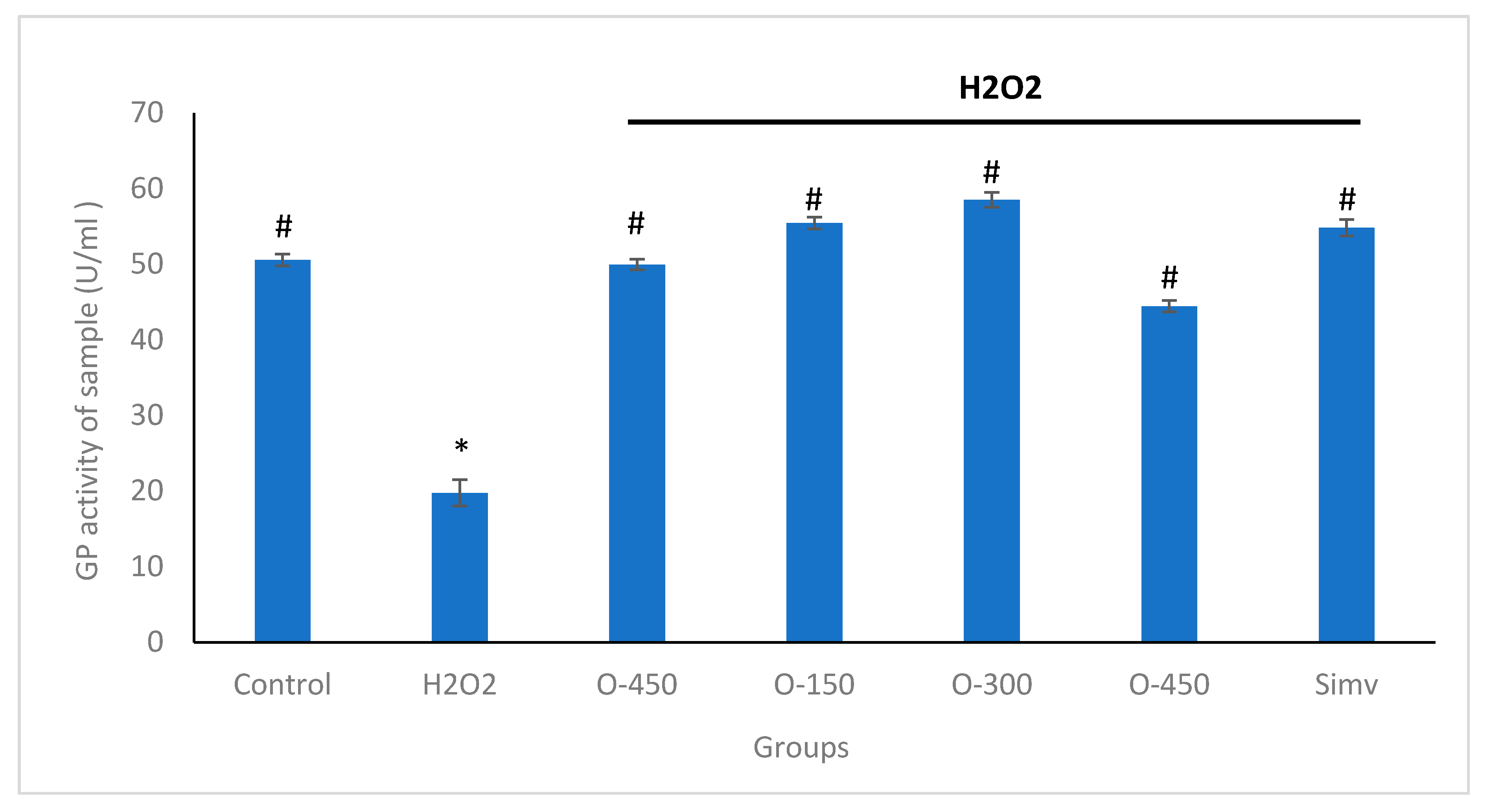
2.4. Effect of Omentin in H2O2-Induced Decreased GPx Activity in HUVECs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. HUVECs Culture
4.3. Treatment Protocols
4.4. Cell Viability Assay
4.5. Hoechst 33358 Fluorescence Staining
4.6. Flow Cytometric Analysis of Apoptosis
4.7. Measurement of Intracellular Reactive Oxygen Species (ROS)
4.8. Measurement of the Intracellular GPx
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daiber, A.; Steven, S.; Weber, A.; Shuvaev, V.V.; Muzykantov, V.R.; Laher, I.; Li, H.; Lamas, S.; Münzel, T. Targeting vascular (endothelial) dysfunction. Br. J. Pharmacol. 2017, 174, 1591–1619. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative stress in atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic-Mirski, K.; Helmstädter, J.; Kröller-Schön, S.; Münzel, T.; et al. Vascular inflammation and oxidative stress: Major triggers for cardiovascular disease. Oxid. Med. Cell Longev. 2019, 2019, 7092151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vascul. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef]
- Jiang, B.; Liang, P.; Zhang, B.; Song, J.; Huang, X.; Xiao, X. Role of PPAR-beta in hydrogen peroxide-induced apoptosis in human umbilical vein endothelial cells. Atherosclerosis. 2009, 204, 353–358. [Google Scholar] [CrossRef]
- Jia, Y.; Ji, L.; Zhang, S.; Xu, L.; Yin, L.; Li, L.; Zhao, Y.; Peng, J. Total flavonoids from Rosa Laevigata Michx fruit attenuates hydrogen peroxide induced injury in human umbilical vein endothelial cells. Food Chem. Toxicol. 2012, 50, 3133–3141. [Google Scholar] [CrossRef]
- Yang, B.; Oo, T.N.; Rizzo, V.; Yang, B.; Oo, T.N.; Rizzo, V. Lipid rafts mediate H2O2 prosurvival effects in cultured endothelial cells. Faseb J. 2006, 20, 1501–1503. [Google Scholar] [CrossRef] [Green Version]
- Tian, R.; Yang, Z.; Lu, N.; Peng, Y.Y. Quercetin, but not rutin, attenuated hydrogen peroxide-induced cell damage via heme oxygenase-1 induction in endothelial cells. Arch. Biochem. Biophys. 2019, 676, 108157. [Google Scholar] [CrossRef]
- Li, Y.; Meng, R. MicroRNA-154 targets the Wnt/β-Catenin signaling pathway following injury to human vascular endothelial cells by hydrogen peroxide. Med. Sci. Monit. 2019, 25, 5648–5656. [Google Scholar] [CrossRef]
- Huang, C.Y.; Ting, W.J.; Huang, C.Y.; Yang, J.Y.; Lin, W.T. Resveratrol attenuated hydrogen peroxide-induced myocardial apoptosis by autophagic flux. Foos Nutr. Res. 2016, 60, 30511. [Google Scholar] [CrossRef] [Green Version]
- Pastori, D.; Nocella, C.; Pignatelli, P.; Novo, M.; Cammisotto, V.; Violi, F.; Carnevale, R. Assessment of blood hydrogen peroxide break-down activity (HBA) in healthy subjects and in patients with atrial fibrillation: Relation to cardiovascular events. Atherosclerosis 2018, 275, 252–253. [Google Scholar] [CrossRef]
- Ballatori, N.; Krance, S.M.; Notenboom, S.; Shi, S.; Tieu, K.; Hammond, C.L. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem. 2009, 390, 191–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza Batista, C.M.; Yang, R.Z.; Lee, M.J.; Glynn, N.M.; Yu, D.Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin plasma levels and gene expression are decreased in obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herder, C.; Carstensen, M.; Ouwens, D.M. Anti-inflammatory cytokines and risk of type 2 diabetes. Diabetes Obes. Metab. 2013, 15, 39–50. [Google Scholar] [CrossRef]
- Yan, P.; Li, L.; Yang, M.; Liu, D.; Liu, H.; Boden, G.; Yang, G. Effects of the long-acting human glucagon-like peptide-1 analog liraglutide on plasma omentin-1 levels in patients with type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2011, 92, 368–374. [Google Scholar] [CrossRef]
- Yamawaki, H.; Tsubaki, N.; Mukohda, M.; Okada, M.; Hara, Y. Omentin, a novel adipokine, induces vasodilation in rat isolated blood vessels. Biochem. Biophys. Res. Commun. 2010, 393, 668–672. [Google Scholar] [CrossRef]
- Yamawaki, H.; Kuramoto, J.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. Omentin, a novel adipocytokine inhibits TNF-induced vascular inflammation in human endothelial cells. Biochem. Biophys. Res. Commun. 2011, 408, 339–343. [Google Scholar] [CrossRef]
- Maruyama, S.; Shibata, R.; Kikuchi, R.; Izumiya, Y.; Rokutanda, T.; Araki, S.; Kataoka, Y.; Ohashi, K.; Daida, H.; Kihara, S.; et al. Fat-derived factor omentin stimulates endothelial cell function and ischemia-induced revascularization via endothelial nitric oxide synthase-dependent mechanism. J. Biol. Chem. 2012, 287, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, Y.; Shibata, R.; Ohashi, K.; Kambara, T.; Enomoto, T.; Uemura, Y.; Ogura, Y.; Yuasa, D.; Matsuo, K.; Nagata, T.; et al. Omentin prevents myocardial ischemic injury through AMP-activated protein kinase-and Akt-dependent mechanisms. J. Am. Coll. Cardiol. 2014, 63, 2722–2733. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.H.; Sun, W.; Li, W.; Hu, H.Z.; Zhou, L.; Jiang, H.H.; Xu, J.X. Calycosin-7-O-β-D-glucoside promotes oxidative stress-induced cytoskeleton reorganization through integrin-linked kinase signaling pathway in vascular endothelial cells. Bmc Complement. Altern. Med. 2015, 15, 315. [Google Scholar] [CrossRef] [Green Version]
- Fico, A.; Paglialunga, F.; Cigliano, L.; Abrescia, P.; Verde, P.; Martini, G.; Iaccarino, I.; Filosa, S. Glucose-6-phosphate dehydrogenase plays a crucial role in protection from redox-stress-induced apoptosis. Cell Death Differ. 2004, 11, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Kazama, K.; Usui, T.; Okada, M.; Hara, Y.; Yamawaki, H. Omentin plays an anti-inflammatory role through inhibition of TNF-α-induced superoxide production in vascular smooth muscle cells. Eur. J. Pharmacol. 2012, 686, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.R. and Yang, S. Hydrogen peroxide: A signaling messenger. Antioxid. Redox Signal. 2006, 8, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Zhu, J.; Xiao, Q.; Zhang, L. Reactive oxygen species: Key regulators in vascular health and diseases. Br. J. Pharmacol. 2018, 175, 1279–1292. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Parajuli, K. Reactive oxygen species: A key hallmark of cardiovascular disease. Adv. Med. 2016, 2016, 9152732. [Google Scholar] [CrossRef] [Green Version]
- Milkovic, L.; Cipak Gasparovic, A.; Zarkovic, N. Overview on major lipid peroxidation bioactive factor 4-hydroxynonenal as pluripotent growth-regulating factor. Free Radic. Res. 2015, 49, 850–860. [Google Scholar] [CrossRef]
- Chapple, S.J.; Cheng, X.; Mann, G.E. Effect of 4-hydroxynonenal on vascular endothelial and smooth muscle cell redox signalling and function in health and disease. Redox Biol. 2013, 1, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Cesar, V.; Jozić, I.; Begović, L.; Vuković, T.; Mlinarić, S.; Lepeduš, H.; Borović Šunjić, S.; Žarković, N. Cell-type-specific modulation of hydrogen peroxide cytotoxicity and 4-hydroxynonenal binding to human cellular proteins in vitro by antioxidant Aloe vera extract. Antioxidant. 2018, 7, 125. [Google Scholar] [CrossRef] [Green Version]
- Coyle, C.H.; Martinez, L.J.; Coleman, M.C.; Spitz, D.R.; Weintraub, N.L.; Kader, K.N. Mechanism of H2O2-induced oxidative stress in endothelial cells. Free Radic. Biol. Med. 2006, 40, 2206–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devasvaran, K.; Tan, J.J.; Ng, C.T.; Fong, L.Y.; Yong, Y.K. malaysian tualang honey inhibits hydrogen peroxide-induced endothelial hyperpermeability. Oxid. Med. Cell Longev. 2019, 2019, 1202676. [Google Scholar] [CrossRef]
- Manoharan, G.; Jaiswal, S.R.; Singh, J. Effect of α, β momorcharin on viability, caspase activity, cytochrome c release and on cytosolic calcium levels in different cancer cell lines. Mol. Cell Biochem. 2014, 388, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Guo, H.; DaSilva, N.A.; Li, D.; Zhang, K.; Wan, Y.; Gao, X.H.; Chen, H.D.; Seeram, N.P.; Ma, H. Pomegranate (Punica granatum) phenolics ameliorate hydrogen peroxide-induced oxidative stress and cytotoxicity in human keratinocytes. J. Funct. Foods. 2019, 54, 559–567. [Google Scholar] [CrossRef]
Sample availability: Samples of the compound were purchased from Biovendor (Candler, NC, USA). |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binti Kamaruddin, N.A.; Fong, L.Y.; Tan, J.J.; Abdullah, M.N.H.; Singh Cheema, M.; Bin Yakop, F.; Yong, Y.K. Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury. Molecules 2020, 25, 2534. https://doi.org/10.3390/molecules25112534
Binti Kamaruddin NA, Fong LY, Tan JJ, Abdullah MNH, Singh Cheema M, Bin Yakop F, Yong YK. Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury. Molecules. 2020; 25(11):2534. https://doi.org/10.3390/molecules25112534
Chicago/Turabian StyleBinti Kamaruddin, Nur Aqilah, Lai Yen Fong, Jun Jie Tan, Muhammad Nazrul Hakim Abdullah, Manraj Singh Cheema, Fahmi Bin Yakop, and Yoke Keong Yong. 2020. "Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury" Molecules 25, no. 11: 2534. https://doi.org/10.3390/molecules25112534
APA StyleBinti Kamaruddin, N. A., Fong, L. Y., Tan, J. J., Abdullah, M. N. H., Singh Cheema, M., Bin Yakop, F., & Yong, Y. K. (2020). Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury. Molecules, 25(11), 2534. https://doi.org/10.3390/molecules25112534