
Accumulation of Phenolic Compounds and Antioxidant Capacity during Berry Development in Black ‘Isabel’ Grape (Vitis vinifera L. x Vitis labrusca L.)

 , , and
, , and
Abstract
:1. Introduction
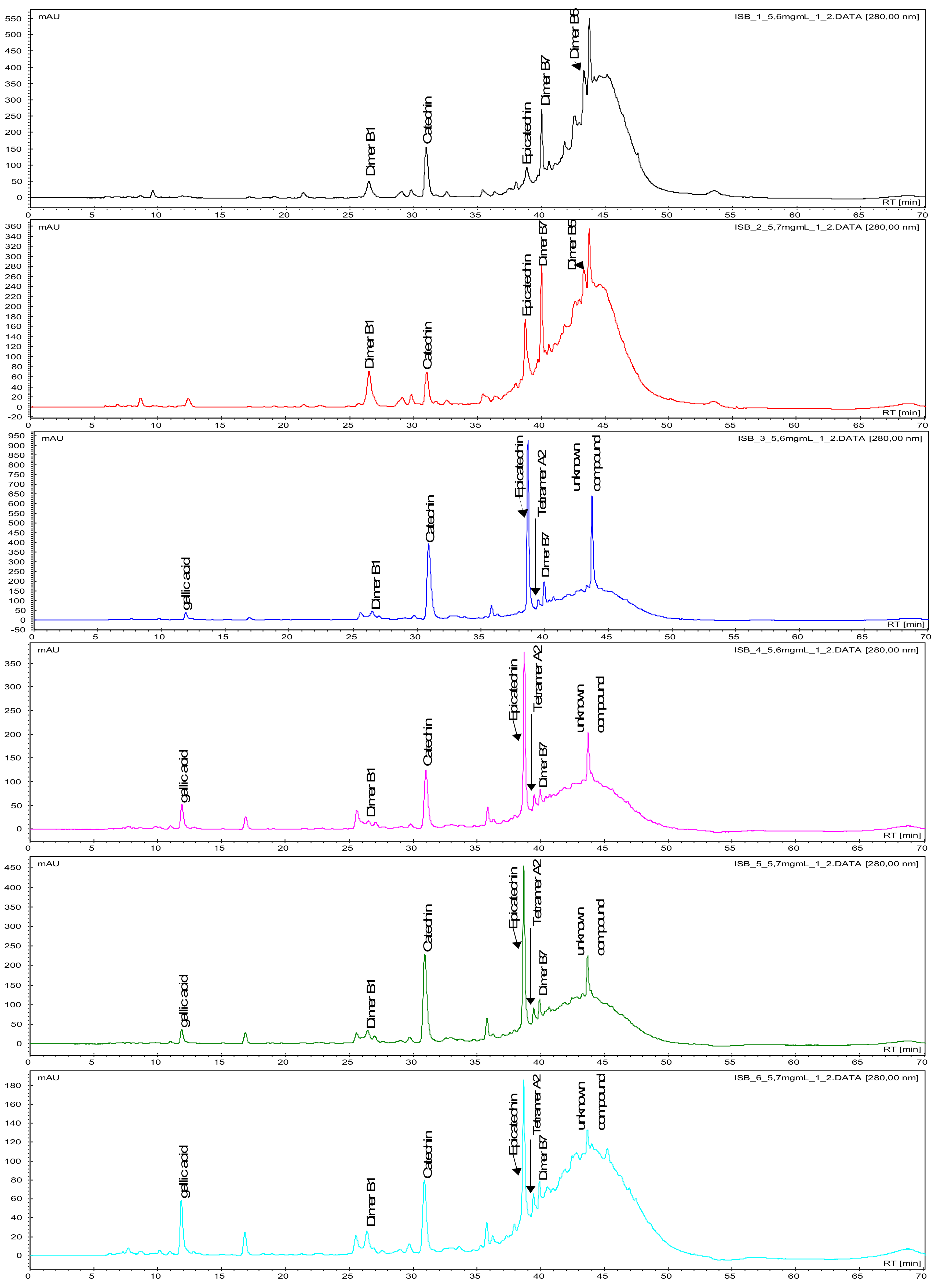
2. Results and Discussion
2.1. Phenolic Compounds Content and Antioxidant Capacity during Berry Development
2.2. Anthocyanin Composition during Berry Development
2.3. Seed Proanthocyanidins during Berry Development
2.4. Principal Component Analysis (PCA) of the Anthocyanins and Proanthocyanidins during Berry Development
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Extraction of Whole Berries for Anthocyanin Analysis by HPLC-PDA and LC-MS/MS
3.4. Extraction of Skin Anthocyanins for Analysis by HPLC-PDA and LC-MS/MS
3.5. Preparation of Seeds for Proanthocyanidin Analysis by HPLC-PDA and LC-MS/MS
3.6. Analysis of Anthocyanins in Grape Berries by HPLC-PDA Analysis
3.7. Proanthocyanidin Characterization of Grape Seeds by LC-MS/MS Analysis
3.8. HPLC-PDA Analyses of Grape Seed Proanthocyanidins on a Reversed-Phase Column
3.9. HPLC-PDA Analyses of Grape Seed Proanthocyanidins on a Diol-column
3.10. Determination of Phenolic Compounds by Spectroscopic Methods
3.11. Determination of the Antioxidant Capacity by DPPH and CUPRAC Assays
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lago-Vanzela, E.S.; Da-Silva, R.; Gomes, E.; García-Romero, E.; Hermosín-Gutiérrez, I. Phenolic composition of the Brazilian seedless table grape varieties BRS Clara and BRS Morena. J. Agric. Food Chem. 2011, 59, 8314–8323. [Google Scholar] [CrossRef] [PubMed]
- Lago-Vanzela, E.S.; Da-Silva, R.; Gomes, E.; García-Romero, E.; Hermosín-Gutiérrez, I. Phenolic composition of the edible parts (flesh and skin) of Bordô grape (Vitis labrusca) using HPLC–DAD–ESI-MS/MS. J. Agric. Food Chem. 2011, 59, 13136–13146. [Google Scholar] [CrossRef] [PubMed]
- Tomaz, I.; Huzanić, N.; Preiner, D.; Stupić, D.; Andabaka, Ž.; Maletić, E.; Kontić, J.K.; Ašperger, D. Extraction Methods of Polyphenol from Grapes: Extractions of Grape Polyphenols. In Polyphenols in Plants, 2nd ed.; Watson, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 151–167. [Google Scholar]
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Hermosín-Gutiérrez, I.; Gómez-Alonso, S.; Pérez-Navarro, J.; Kurt, A.; Colak, N.; Akpınar, E.; Hayirlioglu-Ayaz, S.; Ayaz, F.A. Vitis Vinifera Turkish grape cultivar Karaerik. Part I: Anthocyanin composition, and identification of a newly found anthocyanin. J. Sci. Food Agric. 2020, 100, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Kurt, A.; Torun, H.; Colak, N.; Seiler, G.; Hayirlioglu-Ayaz, S.; Ayaz, F.A. Nutrient profiles of the hybrid grape cultivar ‘Isabel’during berry maturation and ripening. J. Sci. Food Agric. 2017, 97, 2468–2479. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, Y.; Lu, J. Phenolic contents and compositions in skins of red wine grape cultivars among various genetic backgrounds and originations. Int. J. Mol. Sci. 2012, 13, 3492–3510. [Google Scholar] [CrossRef] [Green Version]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Nixdorf, S.L.; Hermosín-Gutiérrez, I. Brazilian red wines made from the hybrid grape cultivar Isabel: Phenolic composition and antioxidant capacity. Anal. Chim. Acta. 2010, 659, 208–215. [Google Scholar] [CrossRef]
- Yamamoto, L.Y.; Marinho de Assis, A.; Roberto, S.R.; Bovolenta, Y.R.; Nixdorf, S.L.; Garcia-Romero, E.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I. Application of abscisic acid (S-ABA) to cv. Isabel grapes (Vitis vinifera x Vitis labrusca) for color improvement: Effect on color, phenolic composition and antioxidant capacity of their grape juice. Food Res. Int. 2015, 77, 572–583. [Google Scholar] [CrossRef]
- Burin, V.M.; Ferreira-Lima, N.E.; Panceri, C.P.; Bordignon-Luiz, M.T. Bioactive compounds and antioxidant activity of Vitis vinifera and Vitis labrusca grapes: Evaluation of different extraction methods. Microchem. J. 2014, 114, 155–163. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Changes in grape seed polyphenols during fruit ripening. Phytochemistry 2000, 55, 77–85. [Google Scholar] [CrossRef]
- Balik, J.; Kumšta, M. Evaluation of colour content in grapes originating from South Moravia. Czech J. Food Sci. 2009, 26, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Duan, C.Q.; Wang, J. Anthocyanins profile of grape berries of Vitis amurensis, its hybrids and their wines. Int. J. Mol. Sci. 2010, 11, 2212–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega-Regules, A.; Romero-Cascales, I.; García, J.M.R.; Bautista-Ortín, A.B.; López-Roca, J.M.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Anthocyanins and tannins in four grape varieties (Vitis vinifera L.). Evolution of their content and extractability. OENO ONE 2008, 42, 147–156. [Google Scholar] [CrossRef]
- Azuma, A.; Yakushiji, H.; Sato, A. Postharvest light irradiation and appropriate temperature treatment increase anthocyanin accumulation in grape berry skin. Postharvest Biol. Tec. 2019, 147, 89–99. [Google Scholar] [CrossRef]
- Balík, J.; Kumšta, M.; Rop, O. Comparison of anthocyanins present in grapes of Vitis vinifera L. varieties and interspecific hybrids grown in the Czech Republic. Chem. Pap. 2013, 67, 1285–1292. [Google Scholar]
- Nile, S.H.; Kim, D.H.; Keum, Y.S. Determination of anthocyanin content and antioxidant capacity of different grape varieties. Ciênc. Téc. Vitiviníc. 2015, 30, 60–68. [Google Scholar]
- He, J.; Giusti, M.M. Anthocyanins: Natural colourants with health-promoting properties. Rev Food Sci Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef]
- Liang, Z.C.; Wu, B.H.; Fan, P.; Yang, C.; Duan, W.; Zheng, X.; Liu, C.; Li, S. Anthocyanin composition and content in grape berry skin in Vitis germplasm. Food Chem. 2008, 111, 837–844. [Google Scholar] [CrossRef]
- Castillo-Muñoz, N.; Gómez-Alonso, S.; García-Romero, E.; Gómez, M.V.; Velders, A.H.; Hermosín-Gutiérrez, I. Flavonol 3-O-glycosides series of Vitis vinifera cv. Petit Verdot red wine grapes. J. Agric. Food Chem. 2008, 57, 209–219. [Google Scholar]
- Castillo-Munoz, N.; Fernandez-Gonzalez, M.; Gomez-Alonso, S.; Garcia-Romero, E.; Hermosin Gutierrez, I. Red-color related phenolic composition of Garnacha Tintorera (Vitis vinifera L.) grapes and red wines. J. Agric. Food Chem. 2009, 57, 7883–7891. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.; Zhang, L.; Li, J.; Zoecklein, B.; Zhou, K. Antioxidant properties and bioactive components of Norton (Vitis aestivalis) and Cabernet Franc (Vitis vinifera) wine grapes. LWT-Food Sci. Technol. 2009, 42, 1269–1274. [Google Scholar] [CrossRef]
- Favre, G.; Hermosín-Gutiérrez, I.; Piccardo, D.; Gómez-Alonso, S.; González-Neves, G. Selectivity of pigments extraction from grapes and their partial retention in the pomace during red wine making. Food Chem. 2019, 277, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; He, J.; Duan, C.; Yu, X.; Zhu, L.; Hie, Z.; Zhang, C.; Xu, W.; Wang, S. Root restriction affects anthocyanin accumulation and composition in berry skin of “Kyoho” grape (Vitis vinifera L. x Vitis labrusca L.) during ripening. Sci. Hortic. 2012, 137, 20–28. [Google Scholar] [CrossRef]
- Hui, Y.H. Handbook of Fruit and Vegetable Flavors; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 1–1083. [Google Scholar]
- Shoeva, O.Y.; Glagoleva, A.Y.; Khlestkina, E.K. The factors affecting the evolution of the anthocyanin biosynthesis pathway genes in monocot and dicot plant species. BMC Plant Biol. 2017, 17, 256. [Google Scholar] [CrossRef] [PubMed]
- Miele, A. Grapevine yield components and composition of Isabel grape produced according to the organic and conventional systems. In Proceedings of the 39th World Congress of Vine and Wine, BIO Web of Conferences, Bento Gonçalves, Brazil, 24–28 October 2016; p. 01011. [Google Scholar]
- Rebello, L.P.G.; Lago-Vanzela, E.S.; Barcia, M.T.; Ramos, A.M.; Stringheta, P.C.; Da-Silva, R.; Castillo-Muñoz, N.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I. Phenolic composition of the berry parts of hybrid grape cultivar BRS Violeta (BRS Rubea × IAC 1398-21) using HPLC-DAD-ESI-MS/MS. Food Resc. Int. 2013, 54, 354–366. [Google Scholar] [CrossRef]
- Cádiz-Gurrea, M.L.; Borrás-Linares, I.; Lozano-Sánchez, J.; Joven, J.; Fernández-Arroyo, S.; Segura Carretero, A. Cocoa and grape seed byproducts as a source of antioxidant and anti-inflammatory proanthocyanidins. Int. J. Mol. Sci. 2017, 18, 376. [Google Scholar] [CrossRef]
- Pantelić, M.M.; Dabić Zagorac, D.Č.; Davidović, S.M.; Todić, S.R.; Bešlić, Z.S.; Gašić, U.M.; Tešić, Ž.L.; Natić, M.M. Identification and quantification of phenolic compounds in berry skin, pulp, and seeds in 13 grapevine varieties grown in Serbia. Food Chem. 2016, 211, 243–252. [Google Scholar]
- Katalinić, V.; Maleš, P. Compositional changes in grape polyphenols throughout maturation. J. Wine Res. 1997, 8, 169–177. [Google Scholar] [CrossRef]
- Niculescu, V.C.; Paun, N.; Ionete, R.E. The evolution of polyphenols from grapes to wines. In Grapes and Wines: Advances in Production, Processing, Analysis and Valorization; Jardao, A.M., Cosme, F., Eds.; IntechOpen: Rijeka, Croatia, 2018; pp. 119–132. [Google Scholar]
- Kyraleou, M.; Kallithraka, S.; Theodorou, N.; Teissedre, P.L.; Kotseridis, Y.; Koundouras, S. Changes in tannin composition of Syrah grape skins and seeds during fruit ripening under contrasting water conditions. Molecules 2017, 22, 1453. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Saona, L.E.; Wrolstad, R.E. Extraction, isolation, and purification of anthocyanins. Curr. Protoc. Food Anal. Chem. 2001, 1, F1-1. [Google Scholar] [CrossRef]
- Ćurko, N.; Ganić, K.K.; Gracin, L.; Ðapic, M.; Jourdes, M.; Teissedre, P.L. Characterization of seed and skin polyphenolic extracts of two red grape cultivars grown in Croatia and their sensory perception in a wine model medium. Food Chem. 2014, 145, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, S.; Lehmann, L.; Winterhalter, P. Rapid characterisation of grape seed extracts by a novel HPLC method on a diol stationary phase. J. Funct. Foods 2015, 15, 225–232. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Huang, D.J.; Chun-Der, L.I.N.; Hsien-Jung, C.; Yaw-Huei, L.I.N. Antioxidant and antiproliferative activities of sweet potato (Ipomoea batatas [L.] LamTainong 57’) constituents. Bot. Bull. Acad. Sin. 2004, 45, 179–186. [Google Scholar]
- Giusti, M.; Rodríguez-Saona, L.E.; Wrolstad, R.E. Molar absorptivity and color characteristics of acylated and non-acylated pelargonidin-based anthocyanins. J. Agric. Food Chem. 1999, 47, 4631–4637. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds of the grape are available from the authors (F.A.A). |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAFB Φ | Berry Development | TPC χ | TF δ | ACY φ | DPPH β | CUPRAC β |
|---|---|---|---|---|---|---|
| Whole berry | ||||||
| 161 | Unripe (green) | 115.3 ± 9.4 f | 12.1 ± 0.1 d | 0.03 ± 0.05 f | 0.32 ± 0.07 f | 8.5 ± 0.8 e |
| 181 | Unripe (green) | 119.1 ± 4.1 e | 13.3 ± 3.2 d | 0.12 ± 0.01 e | 0.72 ± 0.28 e | 13.6 ± 0.5 d |
| 201 | Unripe (green) | 128.2 ± 6.3 d | 18.4 ± 0.1 d | 2.8 ± 0.2 d | 0.98 ± 0.06 d | 19.3 ± 0.4 c |
| 232 | Underripe (reddish) | 235.5 ± 1.0 c | 23.1 ± 0.3 c | 12.1 ± 0.1 c | 5.87 ± 0.03 c | 21.5 ± 0.2 b |
| 263 | Ripe (red) | 312.1 ± 3.3 b | 27.2 ± 0.1 b | 31.7 ± 2 b | 13.5 ± 0.1 b | 26.3 ± 1.3 a,b |
| 293 | Overripe (dark blue black) | 334.4 ± 8.1 a | 32.6 ± 1.3 a | 42.3 ± 2 a | 16.4 ± 0.3 a | 33.5 ± 10.2 a |
| R2(r) | TPC → DPPH | 0.974 (0.987**) | TPC → CUPRAC | 0.848 (0.921**) | ||
| TF → DPPH | 0.924 (0.961**) | TF → CUPRAC | 0.968 (0.984**) | |||
| ACY →DPPH | 0.991 (0.976**) | ACY → CUPRAC | 0.927 (0.912**) | |||
| Skin | ||||||
| 161 | Unripe (green) | 203.1 ± 6.3 d | 10.2 ± 0.3 e | 0.03 ± 0.05 d | 0.75 ± 0.07 f | 16.3 ± 1.1 f |
| 181 | Unripe (green) | 212.7 ± 9.1 c | 13.1 ± 0.1 d | 0.12 ± 0.05 d | 1.42 ± 0.08 e | 21.7 ± 1.3 e |
| 201 | Unripe (green) | 218.3 ± 19.4 c | 18.6 ± 0.1 c | 4.3 ± 0.4 c | 2.37 ± 0.34 d | 26.2 ± 0.4 d |
| 232 | Underripe (reddish) | 229.4 ± 12.7 b | 49.4 ± 1.2 b | 84.2 ± 3.2 b | 11.1 ± 0.2 c | 27.3 ± 0.4 c |
| 263 | Ripe (red) | 383.8 ± 15.6 b | 84.1 ± 1.3 b | 169.5 ± 4.4 a | 18.4 ± 0.2 b | 40.1 ± 2.4 b |
| 293 | Overripe (dark blue black) | 474.2 ± 7.2 a | 93.5 ± 4.0 a | 171.2 ± 2.0 a | 26.2 ± 0.3 a | 44.6 ± 2.1 a |
| R2(r) | TPC → DPPH | 0.951 (0.972**) | TPC → CUPRAC | 0.891 (0.944**) | ||
| TF → DPPH | 0.979 (0.983**) | TF → CUPRAC | 0.915 (0.956**) | |||
| ACY → DPPH | 0.953 (0.968**) | ACY → CUPRAC | 0.876 (0.936**) | |||
| Seed | ||||||
| 201 | Unripe (green) | 218.6 ± 7.2 b | 93.3 ± 2.2 b | n.d. | 74.2 ± 3.1 d | 189 ± 8.3 d |
| 232 | Underripe (reddish) | 259.1 ± 3.1 b | 100.6 ± 6.1 b | n.d. | 92.3 ± 1.4 c | 256 ± 3.5 c |
| 263 | Ripe (red) | 383.5 ± 1.3 a | 117.4 ± 2.4 a | n.d. | 95.7 ± 2.2 b | 296 ± 7.2 b |
| 293 | Overripe (dark blue black) | 384.2 ± 4.2 a | 118.5 ± 9.0 a | n.d. | 128.7 ± 4.6 a | 364 ± 11.6 a |
| R2 (r) | TPC → DPPH | 0.840 (0.917) | TPC → CUPRAC | 0.934 (0.967*) | ||
| TF → DPPH | 0.624 (0.790) | TF → CUPRAC | 0.842 (0.917) | |||
| Compound A | Unripe (green) 161 DAFB | Unripe (green) 181 DAFB | Unripe (green) 201 DAFB | Underripe (reddish) 232 DAFB (veraison) | Ripe (red) 263 DAFB | Overripe (dark blue-black) 293 DAFB |
|---|---|---|---|---|---|---|
| Anthocyanin (mg/kg fw) | ||||||
| Skin | ||||||
| dp-3-glc | 0.12 ± 0.01 a,* | 0.21 ± 0.01 a | 1.37 ± 0.03 b | 14.3 ± 0.3 c | 20.5 ± 1.1 d | 34.5 ± 11.3 e |
| cy-3-glc | 0.31 ± 0.01 a | 0.43 ± 0.02 a | 5.4 ± 0.1 b | 32.4 ± 0.4 c | 40.3 ± 1.4 d | 58.1 ± 2.4 e |
| pt-3-glc | 0.21 ± 0.01 a | 0.23 ± 0.01 a | 2.34 ± 0.1 a | 18.1 ± 0.3 b | 24.7 ± 1.3 c | 31.4 ± 1.2 d |
| pn-3-glc | 0.98 ± 0.01 a | 1.03 ± 0.07 a | 1.98 ± 0.02 a | 154.6 ± 2.3 b | 596.3 ± 3.2 c | 799.2 ± 8.1 d |
| mv-3-glc | 1.05 ± 0.04 a | 2.01 ± 0.06 a | 3.4 ± 0.2 a | 143.2 ± 2.2 b | 1499.2 ± 10.4 c | 1729.3 ± 8.5 d |
| Whole berry | ||||||
| dp-3-glc | ND | 0.02 ± 0.0 a | 0.57 ± 0.08 b | 4.2 ± 0.2 c | 6.2 ± 0.1 d | 10.2 ± 0.4 e |
| cy-3-glc | 0.06 ± 0.01 a | 0.09 ± 0.01 a | 1.24 ± 0.12 b | 9.3 ± 0.2 c | 15.2 ± 0.4 d | 26.1 ± 1.2 e |
| pt-3-glc | 0.04 ± 0.0 a | 0.05 ± 0.01 a | 0.12 ± 0.01 a | 5.3 ± 0.2 b | 7.3 ± 0.2 c | 15.4 ± 1.1 d |
| pn-3-glc | 0.15 ± 0.01 a | 0.34 ± 0.03 a | 0.55 ± 0.06 a | 50.2 ± 1.4 b | 132.2 ± 1.6 c | 202.7 ± 3.3 d |
| mv-3-glc | 0.23 ± 0.03 a | 0.37 ± 0.05 a | 1.27 ± 0.06 a | 73.4 ± 0.1 b | 560.5 ± 14.2 c | 895.2 ± 14.5 d |
| Proanthocyanidins (PAs, %, dw) | ||||||
| Seed | ||||||
| Monomers | 1.47 ± 0.05 a | 3.1 ± 0.1 b,* | 13 ± 0.1 e | 6.7 ± 0.1 d | 4.39 ± 0.01 c | 3.06 ± 0.03 b |
| Galloylated monomers | 1.37 ± 0.04 b | 2.54 ± 0.03 e | 5.26 ± 0.03 f | 2.2 ± 0.1 d | 1.95 ± 0.04 c | 1.28 ± 0.02 a |
| Dimers | 1.43 ± 0.02 c | 2.15 ± 0.06 d | 3.01 ± 0.04 f | 2.5 ± 0.1 e | 1.61 ± 0.03 b | 1.44 ± 0.06 a |
| Galloylated dimers | 1.89 ± 0.02 f | 1.56 ± 0.02 e | 0.83 ± 0.03 d | 0.35 ± 0.01 c | 0.20 ± 0.01 a | 0.27 ± 0.02 b |
| Trimers | 0.67 ± 0.08 b | 0.85 ± 0.01 c | 0.92 ± 0.04 d | 0.72 ± 0.02 b | 0.50 ± 0.01 a | 0.55 ±0.02 a |
| Tetramers | 0.82 ± 0.03 d | 0.80 ± 0.01 c | 0.79 ± 0.03 c | 0.70 ± 0.01 b | 0.58 ± 0.02 a | 0.54 ±0.09 a |
| Pentamers | 0.73 ± 0.07 d | 0.63 ± 0.02 c | 0.64 ± 0.01 c | 0.52 ± 0.03 b | 0.45 ± 0.01 a | 0.41 ± 0.02 a |
| Polymers | 83 ± 1 e | 40 ± 0.1 d | 35 ± 0.2 b | 37 ± 1 c | 35 ± 0.4 b | 19 ± 0.2 a |
| Gallic acid | ND | ND | 0.29 ± 0.00 a | 0.63 ± 0.01 d | 0.52 ± 0.00 c | 0.35 ± 0.00 b |
| Dimer B1 | 0.1 ± 0.00 d | 0.01 ± 0.00 a | 0.03 ± 0.00 c | 0.03 ± 0.00 c | 0.03 ± 0.00 c | 0.02 ± 0.00 b |
| Catechin | 0.58 ± 0.02 a | 2.02 ± 0.03 d | 5.41 ± 0.03 f | 3.01 ± 0.02 e | 1.64 ± 0.01 c | 0.95 ± 0.02 b |
| Dimer B2 | ND | 0.02 ± 0.00 a | 0.08 ± 0.04 b | 0.10 ± 0.01 b | 0.07± 0.02 b | 0.08 ± 0.00 b |
| Epicatechin | 1.1 ± 0.1 a | 2.48 ± 0.04 c | 8.0 ± 0.1 f | 4.56 ± 0.03 e | 3.32 ± 0.02 d | 2.21 ± 0.12 b |
| Tetramer A2 | ND | ND | 0.77 ± 0.06 c | 0.76 ± 0.02 c | 0.17 ± 0.00 a | 0.56 ± 0.05 b |
| Dimer B7 | 2.18 ± 0.04 f | 2.33 ± 0.05 e | 1.72 ± 0.07 d | 1.23 ± 0.04 c | 0.89 ± 0.02 b | 0.76 ± 0.04 a |
| Dimer B5 | 2.98 ± 0.1 e | 2.00 ± 0.22 d | 0 ± 0 a | 1.64 ± 0.13 c | 1.16 ± 0.29 b | 1.29 ± 0.01 b |
| TPAs (+polymers) | 95.5 | 49.5 | 59.5 | 48.7 | 44.7 | 36.6 |
| TPAs (−polymers) | 12.5 | 9.7 | 24.6 | 11.5 | 9.8 | 17.3 |
| Variables | sk dp-3-glc | sk cy-3-glc | sk pt-3-glc | sk pn-3-glc | sk mv-3-glc | wb dp-3-glc | wb cy-3-glc | wb pt-3-glc | wb pn-3-glc | wb mv-3-glc |
|---|---|---|---|---|---|---|---|---|---|---|
| sk dp-3-glcA | - | 0.990 * | 0.981 | 0.963 | 0.916 | 0.999 | 0.999 | 0.995 | 0.984 | 0.948 |
| sk cy-3-glc | - | - | 0.997 | 0.940 | 0.890 | 0.992 | 0.982 | 0.973 | 0.960 | 0.907 |
| sk pt-3-glc | - | - | - | 0.940 | 0.896 | 0.985 | 0.972 | 0.957 | 0.955 | 0.900 |
| sk pn-3-glc | - | - | - | - | 0.990 | 0.966 | 0.971 | 0.952 | 0.995 | 0.991 |
| sk mv-3-glc | - | - | - | - | - | 0.921 | 0.929 | 0.901 | 0.971 | 0.982 |
| wb dp-3-glc | - | - | - | - | - | - | 0.998 | 0.991 | 0.984 | 0.948 |
| wb cy-3-glc | - | - | - | - | - | - | - | 0.996 | 0.990 | 0.962 |
| wb pt-3-glc | - | - | - | - | - | - | - | - | 0.978 | 0.950 |
| wb pn-3-glc | - | - | - | - | - | - | - | - | - | 0.988 |
| wb mv-3-glc | - | - | - | - | - | - | - | - | - | - |
| Variables | F1 | F2 | F3 | F4 | F5 | Observations | F1 | F2 | F3 | F4 | F5 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| sk dp-3-glc A | 0.987 * | 0.011 | 0.002 | 0.000 | 0.000 | Uripe (161 DAFB) | 0.989 | 0.009 | 0.002 | 0.000 | 0.000 |
| sk cy-3-glc | 0.959 | 0.035 | 0.005 | 0.000 | 0.000 | Unripe (181 DAFB) | 0.990 | 0.008 | 0.002 | 0.000 | 0.000 |
| sk pt-3-glc | 0.950 | 0.030 | 0.020 | 0.000 | 0.000 | Unripe (201 DAFB) | 0.999 | 0.000 | 0.000 | 0.001 | 0.000 |
| sk pn-3-glc | 0.974 | 0.025 | 0.002 | 0.000 | 0.000 | Underripe (232 DAFB) | 0.002 | 0.983 | 0.015 | 0.000 | 0.000 |
| sk mv-3-glc | 0.912 | 0.078 | 0.010 | 0.000 | 0.000 | Ripe (263 DAFB) | 0.939 | 0.030 | 0.032 | 0.000 | 0.000 |
| wb dp-3-glc | 0.990 | 0.010 | 0.000 | 0.000 | 0.000 | Overripe (293 DAFB) | 0.996 | 0.000 | 0.004 | 0.000 | 0.000 |
| wb cy-3-glc | 0.993 | 0.003 | 0.004 | 0.000 | 0.000 | Variability (%) | 96.76 | 2.54 | 0.69 | 0.01 | 0.00 |
| wb pt-3-glc | 0.972 | 0.009 | 0.019 | 0.000 | 0.000 | Cumulative (%) | 96.76 | 99.30 | 99.99 | 100.00 | 100.00 |
| wb pn-3-glc | 0.994 | 0.006 | 0.000 | 0.000 | 0.000 | ||||||
| wb mv-3-glc | 0.947 | 0.046 | 0.006 | 0.000 | 0.000 | ||||||
| Variability (%) | 96.76 | 2.54 | 0.69 | 0.01 | 0.00 | ||||||
| Cumulative (%) | 96.76 | 99.30 | 99.99 | 100.00 | 100.00 |
| Variables | M | Gall M | D | Gall D | Tri | Tet | Pent | Pol | GaA | DB1 | C | DB2 | EC | TetA2 | DB7 | DB5 | TPAs(+pol) | TPAs (−pol) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | - | 0.927 * | 0.902 | −0.301 | 0.593 | 0.192 | 0.074 | −0.346 | 0.347 | −0.302 | 0.977 | 0.581 | 0.997 | 0.742 | -0.065 | −0.845 | −0.153 | 0.745 |
| Gall M | - | - | 0.902 | 0.002 | 0.777 | 0.431 | 0.318 | −0.205 | 0.009 | −0.277 | 0.954 | 0.274 | 0.921 | 0.477 | 0.247 | −0.745 | −0.021 | 0.705 |
| D | - | - | - | −0.074 | 0.821 | 0.447 | 0.259 | −0.250 | 0.190 | −0.360 | 0.969 | 0.421 | 0.908 | 0.621 | 0.221 | −0.633 | −0.114 | 0.532 |
| Gall D | - | - | - | - | 0.470 | 0.842 | 0.906 | 0.793 | −0.918 | 0.558 | −0.188 | −0.916 | −0.338 | −0.634 | 0.945 | 0.618 | 0.781 | −0.132 |
| Tri | - | - | - | - | - | 0.802 | 0.657 | 0.095 | −0.388 | −0.139 | 0.723 | −0.124 | 0.578 | 0.210 | 0.699 | −0.244 | 0.213 | 0.426 |
| Tet | - | - | - | - | - | - | 0.965 | 0.664 | −0.646 | 0.405 | 0.323 | −0.583 | 0.163 | −0.238 | 0.943 | 0.281 | 0.708 | 0.090 |
| Pent | - | - | - | - | - | - | - | 0.801 | −0.731 | 0.599 | 0.169 | −0.705 | 0.033 | −0.352 | 0.929 | 0.377 | 0.854 | 0.099 |
| Pol | - | - | - | - | - | - | - | - | −0.567 | 0.913 | −0.314 | −0.768 | −0.386 | −0.563 | 0.662 | 0.720 | 0.963 | −0.256 |
| GaA | - | - | - | - | - | - | - | - | - | −0.369 | 0.257 | 0.904 | 0.386 | 0.658 | −0.829 | −0.480 | −0.582 | 0.007 |
| DB1 | - | - | - | - | - | - | - | - | - | - | −0.355 | −0.563 | −0.356 | −0.355 | 0.355 | 0.602 | 0.928 | −0.066 |
| C | - | - | - | - | - | - | - | - | - | - | - | 0.496 | 0.980 | 0.668 | 0.085 | −0.777 | −0.146 | 0.655 |
| DB2 | - | - | - | - | - | - | - | - | - | - | - | - | 0.608 | 0.876 | −0.779 | −0.727 | −0.694 | 0.353 |
| EC | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.735 | −0.089 | −0.858 | −0.208 | 0.699 |
| TetA2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | −0.495 | −0.697 | −0.398 | 0.652 |
| DB7 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.433 | 0.659 | −0.078 |
| DB5 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.553 | −0.697 |
| TPAs (+pol) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 0.013 |
| TPAs (-pol) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Variables | F1 | F2 | F3 | F4 | F5 | Observations | F1 | F2 | F3 | F4 | F5 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Monomers | 0.560 * | 0.402 | 0.026 | 0.000 | 0.012 | Uripe (161 DAFB) | 0.917 | 0.024 | 0.058 | 0.001 | 0.000 |
| Galloylated monomers | 0.288 | 0.658 | 0.004 | 0.009 | 0.041 | Unripe (181 DAFB) | 0.354 | 0.103 | 0.540 | 0.003 | 0.000 |
| Dimers | 0.347 | 0.575 | 0.009 | 0.067 | 0.003 | Unripe (201 DAFB) | 0.371 | 0.607 | 0.007 | 0.013 | 0.001 |
| Galloylated dimers | 0.673 | 0.295 | 0.021 | 0.006 | 0.005 | Underripe (232 DAFB) | 0.518 | 0.021 | 0.042 | 0.369 | 0.050 |
| Trimers | 0.010 | 0.855 | 0.093 | 0.007 | 0.035 | Ripe (263 DAFB) | 0.074 | 0.770 | 0.003 | 0.004 | 0.149 |
| Tetramers | 0.207 | 0.748 | 0.004 | 0.037 | 0.004 | Overripe (293 DAFB) | 0.138 | 0.723 | 0.000 | 0.104 | 0.035 |
| Pentamers | 0.354 | 0.637 | 0.007 | 0.002 | 0.000 | Variability (%) | 49.24 | 36.08 | 8.15 | 4.35 | 2.18 |
| Polymers | 0.705 | 0.108 | 0.156 | 0.022 | 0.008 | Cumulative (%) | 49.24 | 85.33 | 93.48 | 97.82 | 100.00 |
| Gallic acid | 0.547 | 0.177 | 0.099 | 0.172 | 0.004 | ||||||
| Dimer B1 | 0.483 | 0.027 | 0.489 | 0.001 | 0.001 | ||||||
| Catechin | 0.469 | 0.512 | 0.000 | 0.012 | 0.007 | ||||||
| Dimer B2 | 0.874 | 0.046 | 0.034 | 0.030 | 0.015 | ||||||
| Epicatechin | 0.600 | 0.364 | 0.013 | 0.003 | 0.019 | ||||||
| Tetramer A2 | 0.718 | 0.024 | 0.100 | 0.003 | 0.154 | ||||||
| Dimer B7 | 0.398 | 0.515 | 0.080 | 0.006 | 0.001 | ||||||
| Dimer B5 | 0.832 | 0.051 | 0.001 | 0.066 | 0.051 | ||||||
| TPAs (+pol) | 0.523 | 0.231 | 0.245 | 0.000 | 0.002 | ||||||
| TPAs (−pol) | 0.274 | 0.269 | 0.086 | 0.341 | 0.031 | ||||||
| Variability (%) | 49.24 | 36.08 | 8.15 | 4.35 | 2.18 | ||||||
| Cumulative (%) | 49.24 | 85.33 | 93.47 | 97.82 | 100.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurt-Celebi, A.; Colak, N.; Hayirlioglu-Ayaz, S.; Kostadinović Veličkovska, S.; Ilieva, F.; Esatbeyoglu, T.; Ayaz, F.A. Accumulation of Phenolic Compounds and Antioxidant Capacity during Berry Development in Black ‘Isabel’ Grape (Vitis vinifera L. x Vitis labrusca L.). Molecules 2020, 25, 3845. https://doi.org/10.3390/molecules25173845
Kurt-Celebi A, Colak N, Hayirlioglu-Ayaz S, Kostadinović Veličkovska S, Ilieva F, Esatbeyoglu T, Ayaz FA. Accumulation of Phenolic Compounds and Antioxidant Capacity during Berry Development in Black ‘Isabel’ Grape (Vitis vinifera L. x Vitis labrusca L.). Molecules. 2020; 25(17):3845. https://doi.org/10.3390/molecules25173845
Chicago/Turabian StyleKurt-Celebi, Aynur, Nesrin Colak, Sema Hayirlioglu-Ayaz, Sanja Kostadinović Veličkovska, Fidanka Ilieva, Tuba Esatbeyoglu, and Faik Ahmet Ayaz. 2020. "Accumulation of Phenolic Compounds and Antioxidant Capacity during Berry Development in Black ‘Isabel’ Grape (Vitis vinifera L. x Vitis labrusca L.)" Molecules 25, no. 17: 3845. https://doi.org/10.3390/molecules25173845