
Development of One Pot Strategy for Hyper Production and In Vivo Evaluation of Lovastatin

 ,
,
Abstract
:
1. Introduction
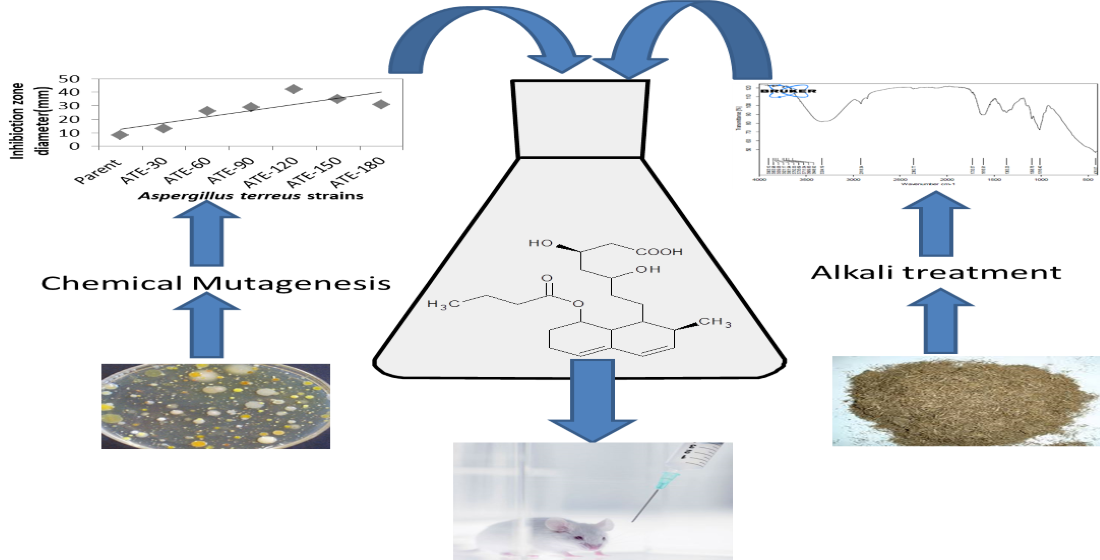
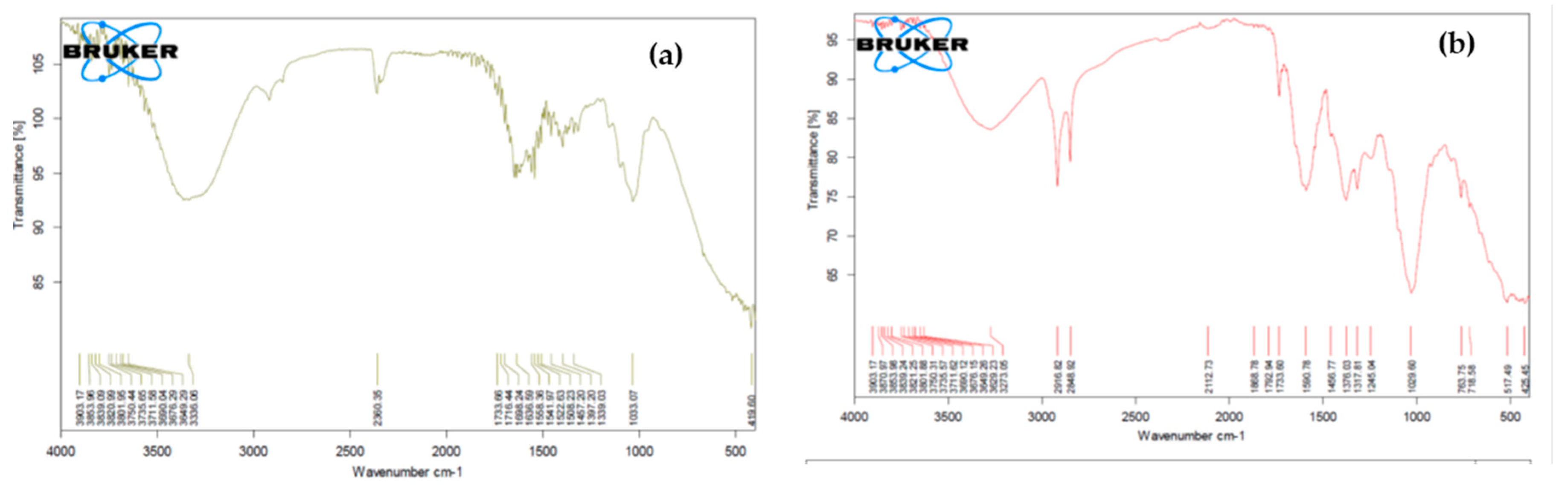
2. Results and Discussion
3. Material and Methods
3.1. Pretreatment of Substrate
3.2. Mutagenic Treatments
3.3. Selection of Best Mutant
3.4. Lovastatin Production
3.5. Extraction of Lovastatin
3.6. Analysis of Lovastatin
3.7. In Vivo Application of Lovastatin
- Group 1 contains normal rats on normal diet
- Group 2 contains rats with high cholesterol diet (HCD)
- Group 3 HCD+ fermented lovastatin derived from parent Aspergillus terreus strain (PL)
- Group 4 HCD+ fermented lovastatin derived from mutant Aspergillus terreus strain (ML)
- Group 5 contains rats which are on synthetic statin
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seenivasan, A.; Sankar, P.; Gummadi, S.N.; Panda, T.; Ch, V.; Eswari, J.S.; Venkateswarlu, C. Metabolic pathway analysis and dynamic macroscopic model development for lovastatin production by Monascus purpureus using metabolic footprinting concept. Biochem. Eng. J. 2020, 154, 107437. [Google Scholar] [CrossRef]
- Lakshmanan, D.; Radha, K. An effective quantitative estimation of lovastatin from Pleurotus ostreatus using UV and HPLC. Int. J. Pharm. Pharm. Sci. 2012, 4, 462–464. [Google Scholar]
- Zhang, Y.; Chen, Z.; Wen, Q.; Xiong, Z.; Cao, X.; Zheng, Z.; Zhang, Y.; Huang, Z. An overview on the biosynthesis and metabolic regulation of monacolin K/lovastatin. Food Funct. 2020. [Google Scholar] [CrossRef]
- Stancu, C.; Sima, A. Statins: Mechanism of action and effects. J. Cell. Mol. Med. 2007, 5, 378–387. [Google Scholar] [CrossRef]
- Groll, M.; Huber, R.; Potts, B.C.M. Crystal Structures of Salinosporamide A (NPI-0052) and B (NPI-0047) in Complex with the 20S Proteasome Reveal Important Consequences of β-Lactone Ring Opening and a Mechanism for Irreversible Binding. J. Am. Chem. Soc. 2006, 128, 5136–5141. [Google Scholar] [CrossRef] [PubMed]
- Thurman, A.J.; Potter, L.A.; Kim, K.; Tassone, F.; Banasik, A.; Potter, S.N.; Bullard, L.; Nguyen, V.; McDuffie, A.; Hagerman, R.; et al. Controlled trial of lovastatin combined with an open-label treatment of a parent-implemented language intervention in youth with fragile X syndrome. J. Neurodev. Disord. 2020, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Namazi, M.R. Statins: Novel additions to the dermatologic arsenal? Exp. Dermatol. 2004, 13, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Parekh, S.; Vinci, V.A.; Strobel, R.J. Improvement of microbial strains and fermentation processes. Appl. Microbiol. Biotechnol. 2000, 54, 287–301. [Google Scholar] [CrossRef]
- Awan, M.S.; Tabbasam, N.; Ayub, N.; Babar, M.E.; Rahman, M.-U.; Rana, S.M.; Rajoka, M.I. Gamma radiation induced mutagenesis in Aspergillus niger to enhance its microbial fermentation activity for industrial enzyme production. Mol. Biol. Rep. 2010, 38, 1367–1374. [Google Scholar] [CrossRef]
- Javed, S.; Bukhari, S.; Zovia, I.; Meraj, M. Screening of Indigenously Isolated Fungi for Lovastatin Production and Its in vivo Evaluation. Curr. Pharm. Biotechnol. 2014, 15, 422–427. [Google Scholar] [CrossRef]
- Chandel, A.K.; Antunes, F.A.; Anjos, V.; Bell, M.J.V.; Rodrigues, L.N.; Polikarpov, I.; De Azevedo, E.R.; Bernardinelli, O.D.; Rosa, C.A.; Pagnocca, F.C.; et al. Multi-scale structural and chemical analysis of sugarcane bagasse in the process of sequential acid–base pretreatment and ethanol production by Scheffersomyces shehatae and Saccharomyces cerevisiae. Biotechnol. Biofuels 2014, 7, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, S.; Bukhari, S.A.; Ali, M.; Rehman, S.-U. Estimation of Antifungal Activity of Mevastatin Produced by Aspergillus terreus GCBL-03 on pretreated substrate in solid state fermentation. Curr. Pharm. Biotechnol. 2016, 17, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.J.D.M.; Martin, C.; Soares, I.B.; Maior, A.M.S.; Baudel, H.M.; De Abreu, C.A.M. Dilute mixed-acid pretreatment of sugarcane bagasse for ethanol production. Biomass Bioenergy 2011, 35, 663–670. [Google Scholar] [CrossRef]
- Rabelo, S.; Carrere, H.; Filho, R.M.; Da Costa, A.C. Production of bioethanol, methane and heat from sugarcane bagasse in a biorefinery concept. Bioresour. Technol. 2011, 102, 7887–7895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, V.S.; Holtzapple, M.T. Fundamental factors affecting enzymatic reactivity. App. Biochem. Biotechnol. 2000, 84, 5–37. [Google Scholar] [CrossRef]
- Ko, J.K.; Bak, J.S.; Jung, M.W.; Lee, H.J.; Choi, I.-G.; Kim, T.H.; Kim, K.H.; Jung, Y.H. Ethanol production from rice straw using optimized aqueous-ammonia soaking pretreatment and simultaneous saccharification and fermentation processes. Bioresour. Technol. 2009, 100, 4374–4380. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.; Pitman, A. FTIR studies of the changes in wood chemistry following decay by brown-rot and white-rot fungi. Int. Biodeterior. Biodegrad. 2003, 52, 151–160. [Google Scholar] [CrossRef]
- Fajula, X.C.; Carrillo, F.; Nogués, F.; Garriga, P. Structural analysis of photodegraded wood by means of FTIR spectroscopy. Polym. Degrad. Stab. 2003, 80, 543–549. [Google Scholar] [CrossRef]
- Pandey, K.K. Study of the effect of photo-irradiation on the surface chemistry of wood. Polym. Degrad. Stab. 2005, 90, 9–20. [Google Scholar] [CrossRef]
- Hinterstoisser, B.; Salmén, L. Two-Dimensional step-scan FTIR: A tool to unravel the OH-valency-range of the spectrum of Cellulose I. Cellulose 1999, 6, 251–263. [Google Scholar] [CrossRef]
- Chisti, Y.; Ferrón, M.A.V.; López, J.C.; Pérez, J.S.; Fernández-Sevilla, J.M. Rapid screening of Aspergillus terreus mutants for overproduction of lovastatin. World J. Microbiol. Biotechnol. 2005, 21, 123–125. [Google Scholar] [CrossRef]
- Samiee, S.M.; Moazami, N.; Haghighi, S.; Mohseni, F.A.; Mirdamadi, S.; Bakhtiari, M.A. Screening of lovastatin production by filamentous fungi. Iran. Biomed. J. 2003, 7, 29–33. [Google Scholar]
- Mangunwardoyo, W.; Rafliyanti, Y.; Kusmana, D. Bioprospect of lovastatin in Aspergillus spp. from University of Indonesia Culture Collection (UICC). World Appl. Sci. J. 2012, 16, 183–188. [Google Scholar]
- Hamden, K.; Jaouadi, B.; Carreau, S.; Bejar, S.; Elfeki, A. Inhibitory effect of fenugreek galactomannan on digestive enzymes related to diabetes, hyperlipidemia, and liver-kidney dysfunctions. Biotechnol. Bioprocess Eng. 2010, 15, 407–413. [Google Scholar] [CrossRef]
- Shepherd, J. Lipids in health and disease. Biochem. Soc. Trans. 2004, 32, 1051–1056. [Google Scholar] [CrossRef]
- Kamal, A.A.; Thanaa, M.A. Oxidative markers, nitric oxide and Homocysteine alteration in hypocholesterolemic rats: Role of atorvastatin and cinnamon. Int. J. Clin. Exp. Med. 2009, 2, 254–265. [Google Scholar]
- López, J.C.; Pérez, J.S.; Fernández-Sevilla, J.M.; Fernández, F.G.A.; Grima, E.M.; Chisti, Y. Production of lovastatin by Aspergillus terreus: Effects of the C:N ratio and the principal nutrients on growth and metabolite production. Enzym. Microb. Technol. 2003, 33, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Steel, R.G.D. Principles and Procedure of Statistics. In A Biomaterial Approach, 3rd ed.; McGraw Hill: New York, NY, USA, 1997. [Google Scholar]
Sample Availability: Samples of the compounds mutant Aspergillus terreus strain, fermented lovastatin and all chemicals used in this study are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Native | 1% NaOH | 2% NaOH | 3% NaOH | ||||
|---|---|---|---|---|---|---|---|---|
| Lovastatin (mg/L) | Biomass (g) | Lovastatin (mg/L) | Biomass (g) | Lovastatin (mg/L) | Biomass (g) | Lovastatin (mg/L) | Biomass (mg/L) | |
| Parent | 8.35 ± 0.56 | 4.16 ± 0.11 | 18.31 ± 0.96 | 3.28 ± 0.11 | 20.28 ± 0.69 | 3.81 ± 0.67 | 34.54 ± 0.61 | 4.22 ± 0.59 |
| ATE-30 | 11.28 ± 0.56 | 4.28 ± 0.11 | 33.43 ± 1.17 | 3.28 ± 0.14 | 34.17 ± 0.33 | 3.27 ± 0.17 | 37.16 ± 1.11 | 4.33 ± 0.23 |
| ATE-60 | 15.72 ± 5.15 | 3.66 ± 0.21 | 21.33 ± 1.01 | 4.21 ± 0.33 | 46.33 ± 0.56 | 4.31 ± 0.41 | 39.11 ± 1.34 | 3.58 ± 0.27 |
| ATE-90 | 11.67 ± 1.01 | 4.67 ± 0.33 | 48.51 ± 1.22 | 4.61 ± 0.51 | 51.37 ± 0.81 | 4.56 ± 0.61 | 45.65 ± 1.30 | 4.13 ± 0.32 |
| ATE-120 | 16.32 ± 0.97 | 3.84 ± 0.55 | 52.14 ± 1.31 | 4.88 ± 0.41 | 91 ± 1.77 | 4.49 ± 0.27 | 51.12. ± 1.61 | 4.66 ± 0.31 |
| ATE-150 | 12.31 ± 1.21 | 3.54 ± 0.43 | 44.45 ± 1.12 | 3.91 ± 0.31 | 74.76 ± 1.22 | 3.64 ± 0.34 | 53.31 ± 1.51 | 4.31 ± 0.25 |
| ATE-180 | 13.32 ± 0.79 | 4.20 ± 0.34 | 39.18 ± 0.97 | 3.87 ± 0.51 | 64.10 ± 1.22 | 4.12 ± 0.53 | 49.28 ± 1.32 | 4.56 ± 0.35 |
| Groups | Days | Body Weight (g) | Blood Glucose Level (mg/dL) | Serum Cholesterol Level (mg/dL) |
|---|---|---|---|---|
| Control | 20 | 230 ± 10 | 160 ± 12 | 63 ± 5 |
| 40 | 265 ± 13 | 186 ± 11 | 61 ± 3 | |
| 60 | 281 ± 14 | 180 ± 15 | 58 ± 5 | |
| HCD | 20 | 221 ± 11 | 470 ± 12 | 75 ± 4 |
| 40 | 245 ± 15 | 510 ± 13 | 70 ± 7 | |
| 60 | 274 ± 18 | 540 ± 11 | 63 ± 6 | |
| HCD-PL | 20 | 215 ± 15 | 430 ± 10 | 65 ± 10 |
| 40 | 232 ± 10 | 390 ± 9 | 61 ± 3 | |
| 60 | 250 ± 13 | 425 ± 14 | 68 ± 4 | |
| HCD-ML | 20 | 213 ± 9 | 410 ± 15 | 55 ± 5 |
| 40 | 235 ± 13 | 360 ± 13 | 62 ± 3 | |
| 60 | 250 ± 16 | 350 ± 11 | 58 ± 5 | |
| HCD-SA | 20 | 220 ± 17 | 445 ± 14 | 60 ± 6 |
| 40 | 231 ± 12 | 418 ± 12 | 65 ± 7 | |
| 60 | 234 ± 14 | 390 ± 10 | 67 ± 4 |
| Groups | Days | AST (mmol/L) | ALT (mmol/L) | HDL-C (mmol/L) | LDL-C (mmol/L) | TG (mmol/L) | TC (mmol/L) | %TC/HDL-C |
|---|---|---|---|---|---|---|---|---|
| Control | 20 | 188 ± 8.44 | 62 ± 4.44 | 0.71 ± 0.03 | 0.50 ± 0.02 | 1.3 ± 0.01 | 1.82 ± 0.04 | 2.5 |
| 40 | 182 ± 10.11 | 60 ± 3.11 | 0.78 ± 0.05 | 0.48 ± 0.03 | 1.24 ± 0.03 | 1.75 ± 0.03 | 2.2 | |
| 60 | 183 ± 7.90 | 61 ± 8.16 | 0.75 ± 0.09 | 0.50 ± 0.05 | 1.4 ± 0.04 | 1.8 ± 0.00 | 2.4 | |
| HCD | 20 | 240 ± 11.21 | 58 ± 7.32 | 0.38 ± 0.04 | 7.45 ± 0.91 | 1.5 ± 0.02 | 7.3 ± 0.12 | 19.2 |
| 40 | 236 ± 9.38 | 57 ± 5.87 | 0.36 ± 0.01 | 7.98 ± 0.24 | 1.57 ± 0.05 | 6.88 ± 0.08 | 19.1 | |
| 60 | 235 ± 8.47 | 59 ± 5.44 | 0.35 ± 0.03 | 8.84 ± 0.17 | 1.6 ± 0.06 | 6.2 ± 0.14 | 17.7 | |
| HCD-PL | 20 | 209 ± 10.11 | 61 ± 6.10 | 0.50 ± 0.01 | 6.54 ± 0.65 | 1.75 ± 0.04 | 4.2 ± 0.11 | 8.4 |
| 40 | 203 ± 6.38 | 62 ± 6.88 | 0.51 ± 0.05 | 5.63 ± 0.41 | 1.6 ± 0.05 | 4.8 ± 0.08 | 9.4 | |
| 60 | 204 ± 9.19 | 59 ± 6.54 | 0.53 ± 0.09 | 5.87 ± 0.33 | 1.7 ± 0.04 | 4.5 ± 0.09 | 8.5 | |
| HCD-ML | 20 | 195 ± 11.32 | 64 ± 6.21 | 0.61 ± 0.07 | 4.32 ± 0.90 | 1.77 ± 0.08 | 3.45 ± 0.07 | 5.6 |
| 40 | 190 ± 9.67 | 65 ± 8.44 | 0.62 ± 0.06 | 4.18 ± 0.46 | 1.8 ± 0.06 | 3.1 ± 0.01 | 5 | |
| 60 | 191 ± 12.21 | 64 ± 6.11 | 0.65 ± 0.05 | 3.87 ± 0.16 | 1.64 ± 0.01 | 3.87 ± 0.12 | 5.9 | |
| HCD-SS | 20 | 208 ± 12.80 | 53 ± 5.19 | 0.50 ± 0.02 | 6.81 ± 0.80 | 0.9 ± 0.06 | 5.4 ± 0.90 | 10.8 |
| 40 | 206 ± 9.73 | 55 ± 2.08 | 0.49 ± 0.07 | 6.13 ± 0.73 | 0.8 ± 0.07 | 5.2 ± 0.13 | 10.6 | |
| 60 | 207 ± 11.61 | 51 ± 4.17 | 0.47 ± 0.05 | 5.81 ± 0.65 | 1.1 ± 0.05 | 5 ± 0.15 | 10.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azeem, M.; Arshad, M.; Mahmood, S.; Abrar, S.; Zahoor, A.F.; Javed, S.; Tariq, B.; Hayyat, K. Development of One Pot Strategy for Hyper Production and In Vivo Evaluation of Lovastatin. Molecules 2020, 25, 4380. https://doi.org/10.3390/molecules25194380
Azeem M, Arshad M, Mahmood S, Abrar S, Zahoor AF, Javed S, Tariq B, Hayyat K. Development of One Pot Strategy for Hyper Production and In Vivo Evaluation of Lovastatin. Molecules. 2020; 25(19):4380. https://doi.org/10.3390/molecules25194380
Chicago/Turabian StyleAzeem, Muhammad, Muhammad Arshad, Saqib Mahmood, Shazia Abrar, Ameer Fawad Zahoor, Sadia Javed, Bisma Tariq, and Khizar Hayyat. 2020. "Development of One Pot Strategy for Hyper Production and In Vivo Evaluation of Lovastatin" Molecules 25, no. 19: 4380. https://doi.org/10.3390/molecules25194380
APA StyleAzeem, M., Arshad, M., Mahmood, S., Abrar, S., Zahoor, A. F., Javed, S., Tariq, B., & Hayyat, K. (2020). Development of One Pot Strategy for Hyper Production and In Vivo Evaluation of Lovastatin. Molecules, 25(19), 4380. https://doi.org/10.3390/molecules25194380