
In-Vivo Antidiabetic Activity and In-Silico Mode of Action of LC/MS-MS Identified Flavonoids in Oleaster Leaves

 , , , , , , , , ,
, , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material
2.3. Animals
2.4. Preparation of Flavonoids Extracts
2.5. Evaluation of Antidiabetic Activity
2.5.1. Induction of Experimental Diabetes
2.5.2. The Experimental Model of Fasting Glucose Measurement
2.5.3. Evaluation of the Antihyperglycemic Activity
2.6. Oleaster Flavonoids Qualitative Analysis
2.7. Molecular Docking
2.7.1. Preparation of Ligands
2.7.2. Preparation of Receptors
2.7.3. Docking Simulations
3. Results and Discussion
3.1. Yield of Extraction
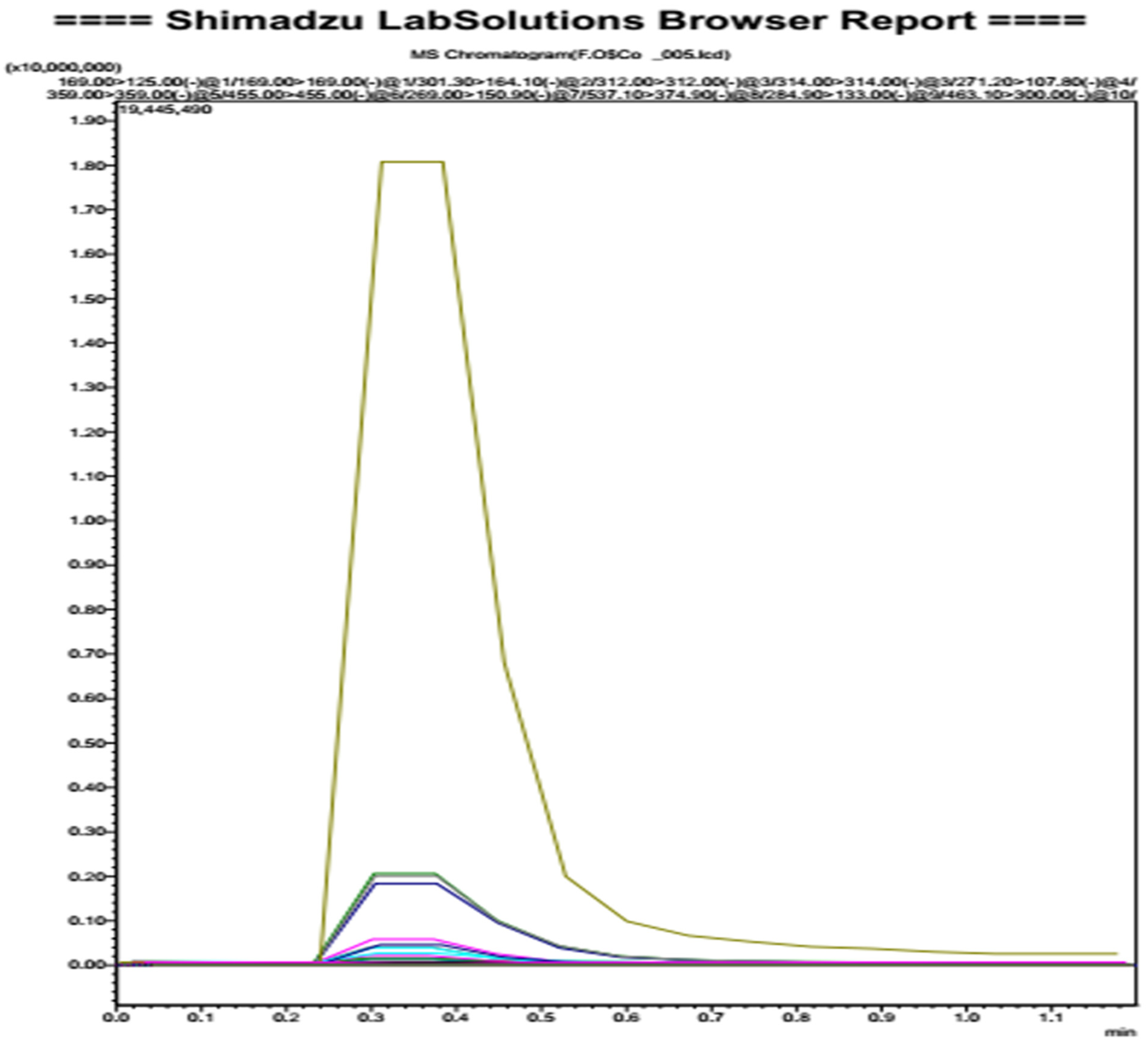
3.2. Oleaster Flavonoids Qualitative Analysis
3.3. Antidiabetic Activity
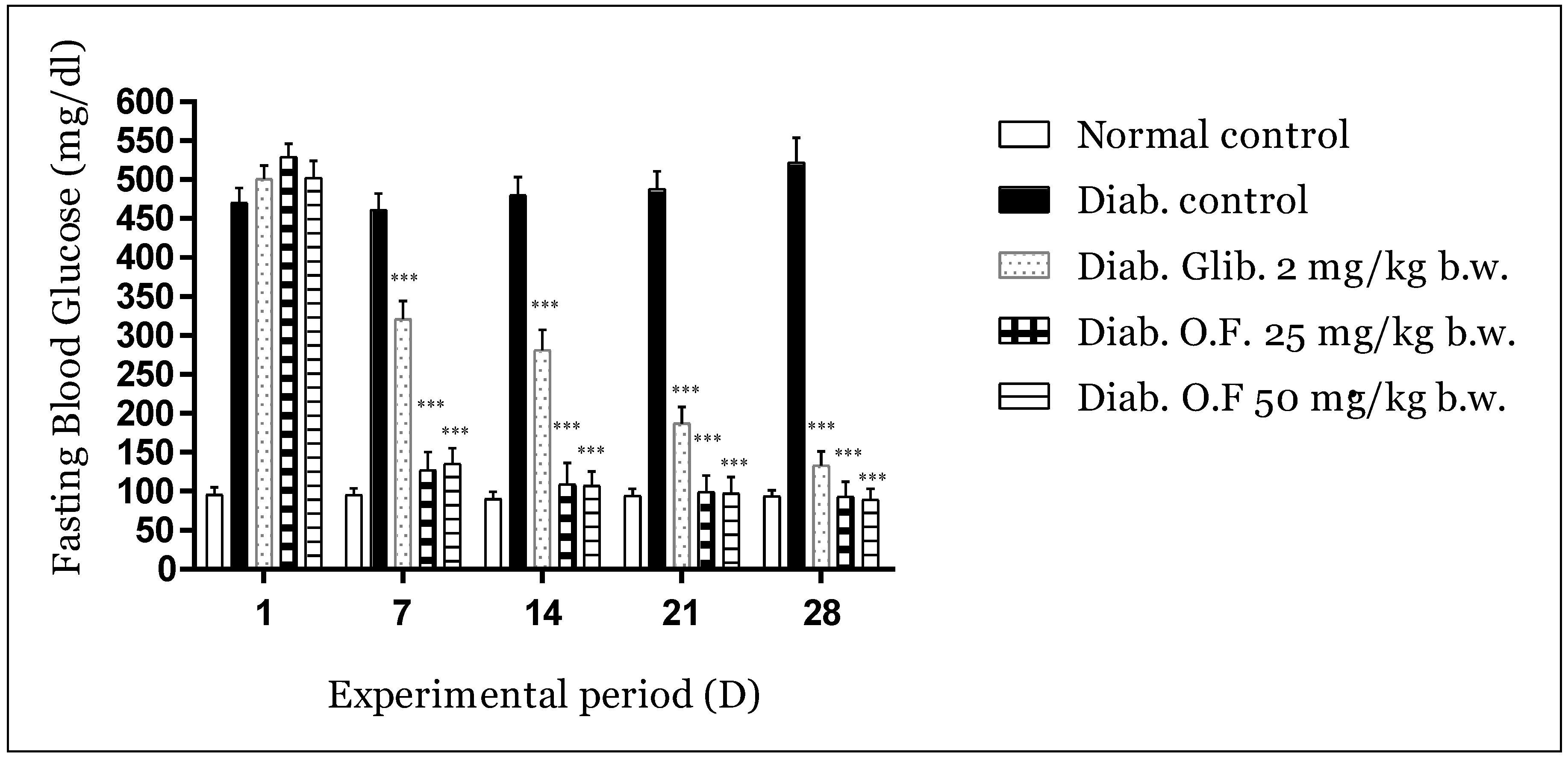
3.3.1. Effect of Oleaster Flavonoids Extract on Glycemia in Diabetic Mice
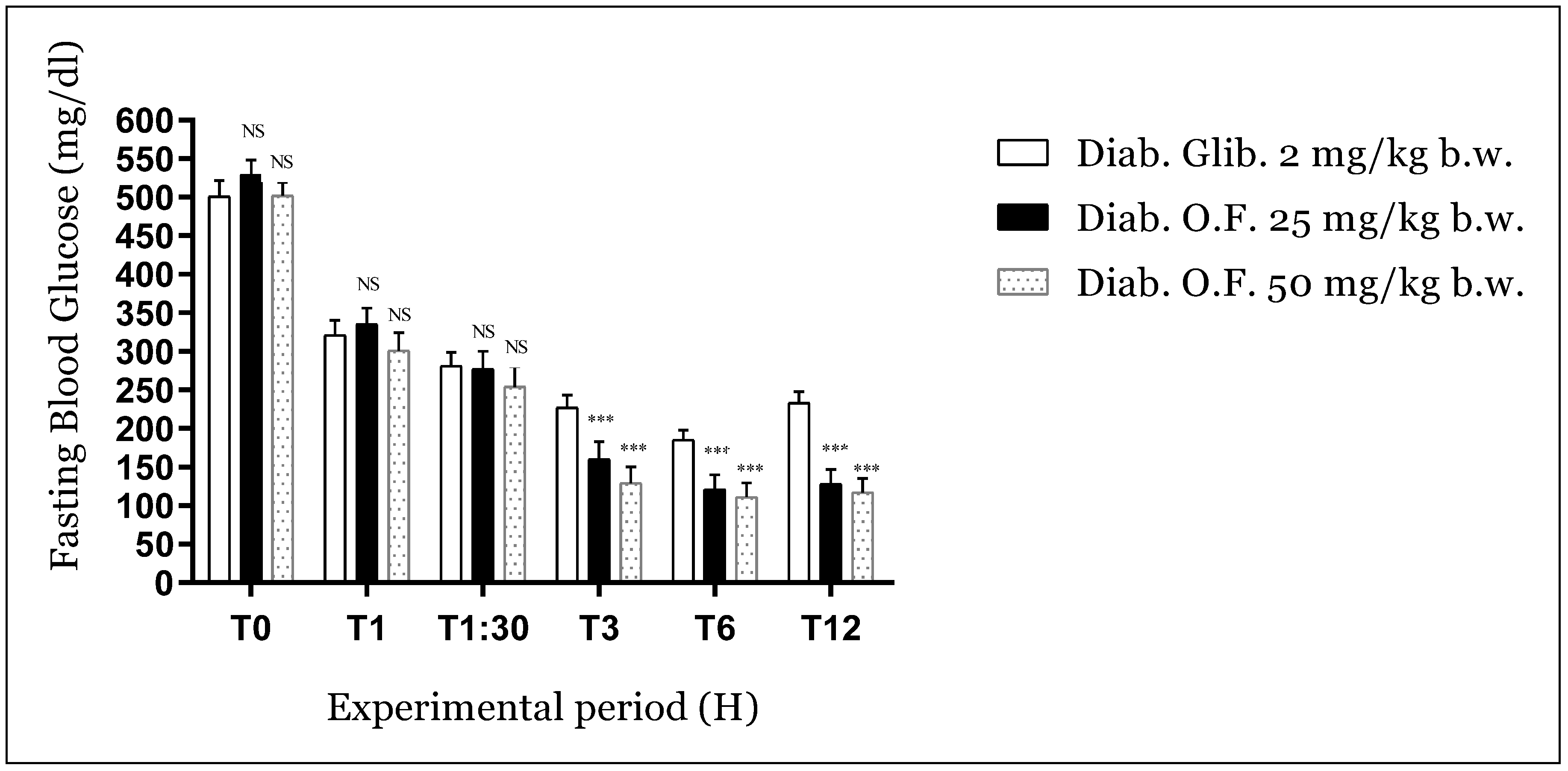
3.3.2. Antihyperglycemic Activity of Oleaster Flavonoids on Diabetic Mice
4. Molecular Docking
4.1. PTP1B
4.2. DPP4
4.3. FFAR1
4.4. α-Amylase
4.5. PPARγ
4.6. α-Glucosidase
4.7. Aldose Reductase
4.8. Glycogen Phosphorylase
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BGL | Blood glucose level |
| GLIB | Glibenclamide |
| DW | distilled water |
| PTP1B | Protein tyrosine phosphatase 1B |
| DPP4 | Dipeptidyl-peptidase 4 |
| AAM | α-Amylase |
| AldR | Aldose reductase |
| GP | Glycogen phosphorylase |
| FFAR1 | Free fatty acid receptor 1 |
| ROS | Reactive oxygen species |
References
- Soumyanath, A. Review of Plants with Anti-Diabetes Mellitus. J. Nat. Prod. 2018, 81, 1916. [Google Scholar] [CrossRef]
- Mekala, K.C.; Bertoni, A.G. Epidemiology of diabetes mellitus. In Transplantation, Bioengineering, and Regeneration of the Endocrine Pancreas; Elsevier BV: Amsterdam, The Netherlands, 2020; pp. 49–58. [Google Scholar]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-C.; Li, L.; Sanborn, T.M.; Shieh, B.; Lenhart, P.; Ammar, D.; LaBarbera, D.V.; Petrash, J.M. Characterization of Emodin as a Therapeutic Agent for Diabetic Cataract. J. Nat. Prod. 2016, 79, 1439–1444. [Google Scholar] [CrossRef] [Green Version]
- Mechchate, H.; Es-Safi, I.; Haddad, H.; Bekkari, H.; Grafov, A.; Bousta, D. Combination of Catechin, Epicatechin, and Rutin:optimization of a novel complete antidiabetic formulation using a mixture design approach. J. Nutr. Biochem. 2020, 108520. [Google Scholar] [CrossRef]
- Es-Safi, I.; Mechchate, H.; Amaghnouje, A.; El Moussaoui, A.; Cerruti, P.; Avella, M.; Grafov, A.; Bousta, D. Marketing and legal status of phytomedicines and food supplements in Morocco. J. Complement. Integr. Med. 2020. [Google Scholar] [CrossRef]
- Besnard, G.; De Casas, R.R.; Christin, P.-A.; Vargas, P. Phylogenetics of Olea (Oleaceae) based on plastid and nuclear ribosomal DNA sequences: Tertiary climatic shifts and lineage differentiation times. Ann. Bot. 2009, 104, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Zaouani, M.; Yahiaoui, F.; Bey, N.N.; Ben-Mahdi, M.H. Antidiarrhoeal activity of aqueous leaf extract of Olea europaea var. sylvestris in albino Wistar rats. J. Biol. Res. 2018, 91. [Google Scholar] [CrossRef]
- Mechchate, H.; Es-Safi, I.; Jawhari, F.Z.; Bari, A.; Grafov, A.; Bousta, D. Ethnobotanical survey about the management of diabetes with medicinal plants used by diabetic patients in Region of Fez-Meknes, Morocco. Ethnobot. Res. Appl. 2020, 19, 1–28. [Google Scholar] [CrossRef] [Green Version]
- National Research Council, Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011. Available online: https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdf (accessed on 17 July 2020).
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The Protein Data Bank. Acta Crystallogr. D 2002, 58, 899–907. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2008, 32, S62–S67. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, G. Insulin and Insulin Resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar]
- Lucchesi, A.N.; Cassettari, L.L.; Spadella, C.T. Alloxan-Induced Diabetes Causes Morphological and Ultrastructural Changes in Rat Liver that Resemble the Natural History of Chronic Fatty Liver Disease in Humans. J. Diabetes Res. 2015, 2015, 494578. [Google Scholar] [CrossRef]
- Gowda, S.; Desai, P.B.; Kulkarni, S.S.; Hull, V.V.; Math, A.A.K.; Vernekar, S.N. Markers of renal function tests. N. Am. J. Med Sci. 2010, 2, 170–173. [Google Scholar] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Elchebly, M. Increased Insulin Sensitivity and Obesity Resistance in Mice Lacking the Protein Tyrosine Phosphatase-1B Gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef]
- Klaman, L.D.; Boss, O.; Peroni, O.D.; Kim, J.K.; Martino, J.L.; Zabolotny, J.M.; Moghal, N.; Lubkin, M.; Kim, Y.-B.; Sharpe, A.H.; et al. Increased Energy Expenditure, Decreased Adiposity, and Tissue-Specific Insulin Sensitivity in Protein-Tyrosine Phosphatase 1B-Deficient Mice. Mol. Cell. Biol. 2000, 20, 5479–5489. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.O.; Ermolieff, J.; Jirousek, M.R. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002, 1, 696–709. [Google Scholar] [CrossRef]
- Wiesmann, C.; Barr, K.J.; Kung, J.; Zhu, J.; Erlanson, D.A.; Shen, W.; Fahr, B.J.; Zhong, M.; Taylor, L.; Randal, M.; et al. Allosteric inhibition of protein tyrosine phosphatase 1B. Nat. Struct. Mol. Biol. 2004, 11, 730–737. [Google Scholar] [CrossRef]
- Jia, Z.; Barford, D.; Flint, A.J.; Tonks, N.K. Structural basis for phosphotyrosine peptide recognition by protein tyrosine phosphatase 1B. Science 1995, 268, 1754–1758. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B. Protein-Tyrosine Phosphatase 1B (PTP1B): A Novel Therapeutic Target for Type 2 Diabetes Mellitus, Obesity and Related States of Insulin Resistance. Curr. Drug Targets Immune. Endocr. Metab. Disord. 2001, 1, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Eröhrborn, D.; Ewronkowitz, N.; Eckel, J. DPP4 in diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [CrossRef] [Green Version]
- Bjelke, J.R.; Christensen, J.; Branner, S.; Wagtmann, N.; Olsen, C.; Kanstrup, A.B.; Rasmussen, H.B. Tyrosine 547 Constitutes an Essential Part of the Catalytic Mechanism of Dipeptidyl Peptidase IV. J. Biol. Chem. 2004, 279, 34691–34697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, C.-H.; Tsai, C.-H.; Lin, C.-H.; Chou, A.C.-Y.; Chen, X. Identification of Hydrophobic Residues Critical for DPP-IV Dimerization. Biochemistry 2006, 45, 7006–7012. [Google Scholar] [CrossRef]
- Srivastava, A.; Yano, J.; Hirozane, Y.; Kefala, G.; Gruswitz, F.; Snell, G.; Lane, W.; Ivetac, A.; Aertgeerts, K.; Nguyen, J.; et al. High-resolution structure of the human GPR40 receptor bound to allosteric agonist TAK-875. Nat. Cell Biol. 2014, 513, 124–127. [Google Scholar] [CrossRef]
- Ren, X.-M.; Cao, L.-Y.; Zhang, J.; Qin, W.-P.; Yang, Y.; Wan, B.; Guo, L.-H. Investigation of the Binding Interaction of Fatty Acids with Human G Protein-Coupled Receptor 40 Using a Site-Specific Fluorescence Probe by Flow Cytometry. Biochemistry 2016, 55, 1989–1996. [Google Scholar] [CrossRef]
- Morgan, N.G.; Dhayal, S. G-protein coupled receptors mediating long chain fatty acid signalling in the pancreatic beta-cell. Biochem. Pharmacol. 2009, 78, 1419–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sum, C.S.; Tikhonova, I.G.; Costanzi, S.; Gershengorn, M.C. Two Arginine-Glutamate Ionic Locks near the Extracellular Surface of FFAR1 Gate Receptor Activation. J. Biol. Chem. 2009, 284, 3529–3536. [Google Scholar] [CrossRef] [Green Version]
- Sum, C.S.; Tikhonova, I.G.; Neumann, S.; Engel, S.; Raaka, B.M.; Costanzi, S.; Gershengorn, M.C. Identification of Residues Important for Agonist Recognition and Activation in GPR40. J. Biol. Chem. 2007, 282, 29248–29255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suvd, D.; Fujimoto, Z.; Takase, K.; Matsumura, M.; Mizuno, H. Crystal Structure of Bacillus stearothermophilus a-Amylase: Possible Factors Determining the Thermostability. J. Biochem. 2001, 129, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Aghajari, N.; Haser, R.; Feller, G.; Gerday, C. Crystal structures of the psychrophilic α-amylase from Alteromonas haloplanctis in its native form and complexed with an inhibitor. Protein Sci. 1998, 7, 564–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiu, J.; Fischer, E.H.; Stein, E.A. Alpha-Amylases as Calcium-Metalloenzymes. II. Calcium and the Catalytic Activity. Biochemistry 1964, 3, 61–66. [Google Scholar] [CrossRef]
- Ragunath, C.; Manuel, S.G.; Venkataraman, V.; Sait, H.B.; Kasinathan, C.; Ramasubbu, N. Probing the Role of Aromatic Residues at the Secondary Saccharide-Binding Sites of Human Salivary α-Amylase in Substrate Hydrolysis and Bacterial Binding. J. Mol. Biol. 2008, 384, 1232–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragunath, C.; Manuel, S.G.A.; Kasinathan, C.; Ramasubbu, N. Structure-function relationships in human salivary α-amylase: Role of aromatic residues in a secondary binding site. Biologia 2008, 63, 47–56. [Google Scholar] [CrossRef]
- Ramasubbu, N.; Ragunath, C.; Mishra, P.J.; Thomas, L.M.; Gyemant, G.; Kandra, L. Human salivary alpha-amylase Trp58 situated at subsite-2 is critical for enzyme activity. JBIC J. Biol. Inorg. Chem. 2004, 271, 2517–2529. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Pochetti, G.; Godio, C.; Mitro, N.; Caruso, D.; Galmozzi, A.; Scurati, S.; Loiodice, F.; Fracchiolla, G.; Tortorella, P.; Laghezza, A.; et al. Insights into the Mechanism of Partial Agonism. J. Biol. Chem. 2007, 282, 17314–17324. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.J. Glucosidase. RCSB Protein Data Bank 2019. [Google Scholar] [CrossRef]
- Alqahtani, A.; Hidayathulla, S.; Rehman, T.; El-Gamal, A.A.; Al-Massarani, S.; Naumovski, V.; Alqahtani, M.S.; El Dib, R.A.; Alajmi, M.F. Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia. Biomolecules 2019, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Hermans, M.M.; Kroos, M.A.; Van Beeumen, J.; Oostra, B.A.; Reuser, A.J. Human lysosomal alpha-glucosidase. Characterization of the catalytic site. J. Biol. Chem. 1991, 266, 13507–13512. [Google Scholar]
- Tang, W.H.; Martin, K.A.; Hwa, J. Aldose Reductase, Oxidative Stress, and Diabetic Mellitus. Front. Pharmacol. 2012, 3, 87. [Google Scholar] [CrossRef] [Green Version]
- Heather, L.C.; Clarke, K. Metabolism, hypoxia and the diabetic heart. J. Mol. Cell. Cardiol. 2011, 50, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.K.; Bohren, K.M.; Gabbay, K.H.; Quiocho, F.A. An unlikely sugar substrate site in the 1.65 A structure of the human aldose reductase holoenzyme implicated in diabetic complications. Science 1992, 257, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Livanova, N.B.; Chebotareva, N.A.; Eronina, T.B.; Kurganov, B.I. Pyridoxal 5″-Phosphate as a Catalytic and Conformational Cofactor of Muscle Glycogen Phosphorylase b. Biochemistry (Moscow) 2002, 67, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Barford, D.; Johnson, L.N. The allosteric transition of glycogen phosphorylase. Nat. Cell Biol. 1989, 340, 609–616. [Google Scholar] [CrossRef]
- Goldsmith, E.J.; Sprang, S.R.; Hamlin, R.; Xuong, N.H.; Fletterick, R.J. Domain separation in the activation of glycogen phosphorylase a. Science 1989, 245, 528–532. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Molecule | Analyzed Fragment | Area under Curve |
|---|---|---|
| Amentoflavone | 537.10 > 374.90 | 88,078 |
| Quercetin-3-O-glucoside | 463.10 > 300.00 | 840,585 |
| Quercetin-3-O-hexose-deoxyhexose | 609.10 > 300.00 | 1,426,485 |
| luteolin-7-O-glucoside | 447.10 > 285.00 | 21,618,792 |
| Oleuropein | 539.00 > 539.00 | 195,134,071 |
| Rutin | 609.00 > 301.00 | 1,000,877 |
| Treatments | Bodyweight Development | Biochemical Parameters | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Day 1 | Day 7 | Day 14 | Day 21 | Day 28 | ASAT (UI/L) | ALAT (UI/L) | UREA (g/L) | CREATININE (mg/L) | |
| Normal Control DW 2 mL/day | 23.4 ± 1.8 ns | 24.7 ± 1.5 * | 25.2 ± 1.4 *** | 26.9 ± 1.3 *** | 27.2 ± 1.5 *** | 253 ± 23.7 *** | 45 ± 7.5 *** | 0.24 ± 0.04 *** | 3.2 ± 0.4 *** |
| Diab. Control DW 2 mL/day | 23.8 ± 1.6 | 21.7 ± 2.2 | 20.1 ± 2.7 | 19.2 ± 2.5 | 17.7 ± 2.4 | 502 ± 38.8 | 134 ± 11.2 | 0.63 ± 0.06 | 5.8 ± 0.8 |
| Diab. Glib. 2 mg/kg/day | 24.3 ± 1.6 ns | 23.1 ± 1.5 ns | 24.2 ± 1.7 ** | 25.8 ± 1.8 *** | 25.9 ± 1.7 *** | 298 ± 24.6 *** | 77 ± 8.8 *** | 0.28 ± 0.04 *** | 4.2 ± 0.4 ** |
| Diab. O.F. 25 mg/kg/day | 23.7 ± 1.2 ns | 24.1 ± 1.3 ns | 25.8 ± 1.1 *** | 26.6 ± 1.2 *** | 28.4 ± 1.1 *** | 223 ± 20.1 *** | 51 ± 7.0 *** | 0.35 ± 0.03 *** | 3.6 ± 0.5 ** |
| Diab. OF. 50 mg/kg/day | 23.5 ± 1.9 ns | 24.5 ± 2.3 ns | 26.1 ± 1.9 *** | 27.6 ± 2.2 *** | 28.9 ± 2.4 *** | 286 ± 18.7 *** | 44 ± 5.5 *** | 0.30 ± 0.03 *** | 3.8 ± 0.4 ** |
| Affinity (kcal/mol) | ||||||||
|---|---|---|---|---|---|---|---|---|
| PTP1B | DPP4 | FFAR1 | Alpha Amylase | PPAR Gamma | Alpha Glucosidase | Aldose Reductase | Glycogen Phosphorylase | |
| Amentoflavone | −8.8 | −10.5 | None | −11.3 | None | −9.5 | −10.0 | −10.7 |
| Quercetin | −6.9 | −8.7 | None | −8.4 | None | −7.3 | −9.6 | −9.8 |
| Rutin | −7.3 | −9.2 | None | −8.4 | None | −8.1 | −8.3 | −8.4 |
| Luteolin-7-O-glucoside | −8.9 | −9.1 | −8.2 | −10.0 | None | −8.5 | −8.8 | −8.2 |
| Oleuropein | −6.6 | −7.3 | −6.2 | −6.9 | None | −7.2 | −9.1 | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mechchate, H.; Es-Safi, I.; Bourhia, M.; Kyrylchuk, A.; El Moussaoui, A.; Conte, R.; Ullah, R.; Ezzeldin, E.; Mostafa, G.A.; Grafov, A.; et al. In-Vivo Antidiabetic Activity and In-Silico Mode of Action of LC/MS-MS Identified Flavonoids in Oleaster Leaves. Molecules 2020, 25, 5073. https://doi.org/10.3390/molecules25215073
Mechchate H, Es-Safi I, Bourhia M, Kyrylchuk A, El Moussaoui A, Conte R, Ullah R, Ezzeldin E, Mostafa GA, Grafov A, et al. In-Vivo Antidiabetic Activity and In-Silico Mode of Action of LC/MS-MS Identified Flavonoids in Oleaster Leaves. Molecules. 2020; 25(21):5073. https://doi.org/10.3390/molecules25215073
Chicago/Turabian StyleMechchate, Hamza, Imane Es-Safi, Mohammed Bourhia, Andrii Kyrylchuk, Abdelfattah El Moussaoui, Raffaele Conte, Riaz Ullah, Essam Ezzeldin, Gamal A. Mostafa, Andriy Grafov, and et al. 2020. "In-Vivo Antidiabetic Activity and In-Silico Mode of Action of LC/MS-MS Identified Flavonoids in Oleaster Leaves" Molecules 25, no. 21: 5073. https://doi.org/10.3390/molecules25215073
APA StyleMechchate, H., Es-Safi, I., Bourhia, M., Kyrylchuk, A., El Moussaoui, A., Conte, R., Ullah, R., Ezzeldin, E., Mostafa, G. A., Grafov, A., Bekkari, H., & Bousta, D. (2020). In-Vivo Antidiabetic Activity and In-Silico Mode of Action of LC/MS-MS Identified Flavonoids in Oleaster Leaves. Molecules, 25(21), 5073. https://doi.org/10.3390/molecules25215073