Small Molecule Soluble Epoxide Hydrolase Inhibitors in Multitarget and Combination Therapies for Inflammation and Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
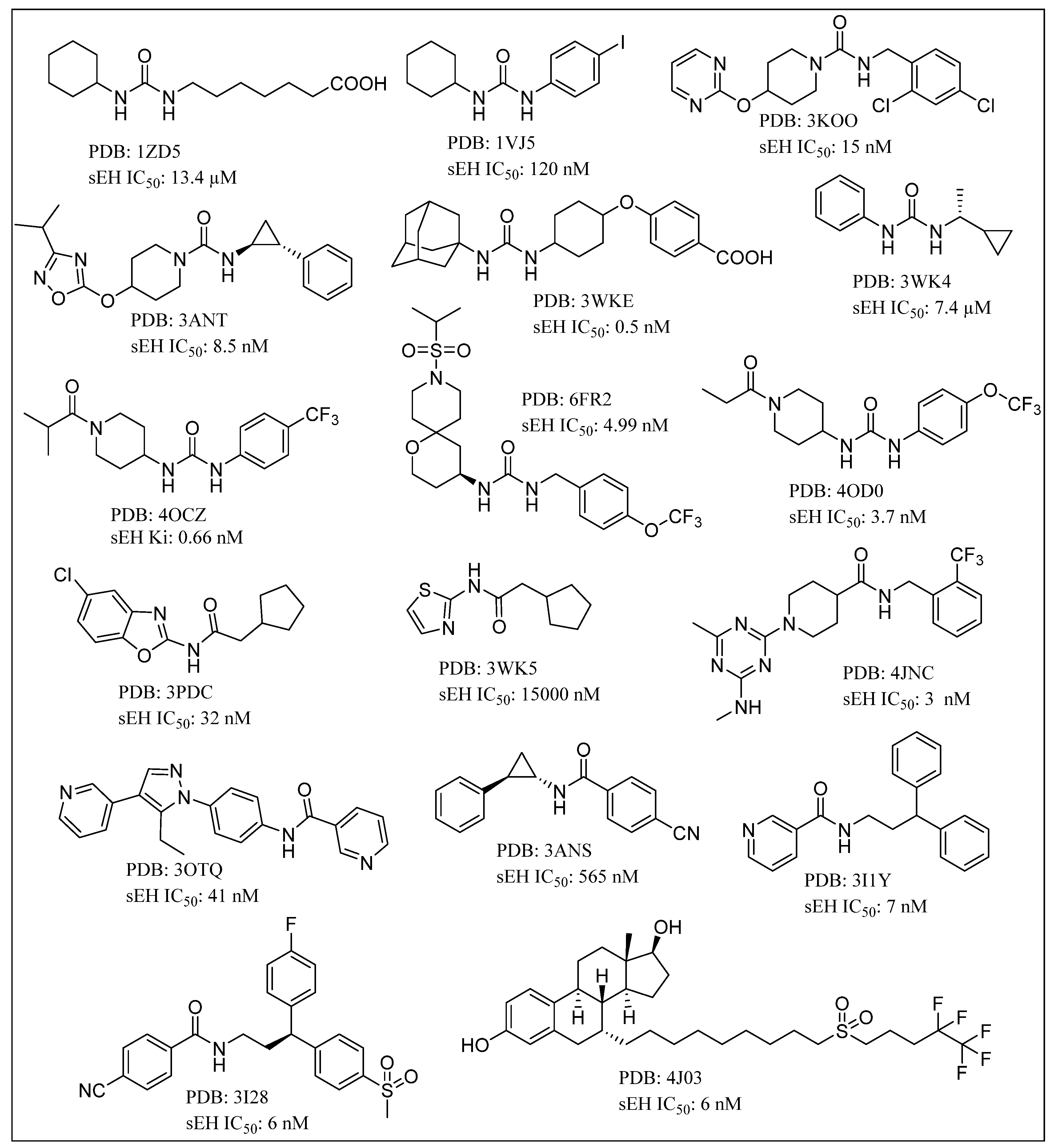{kind=link}
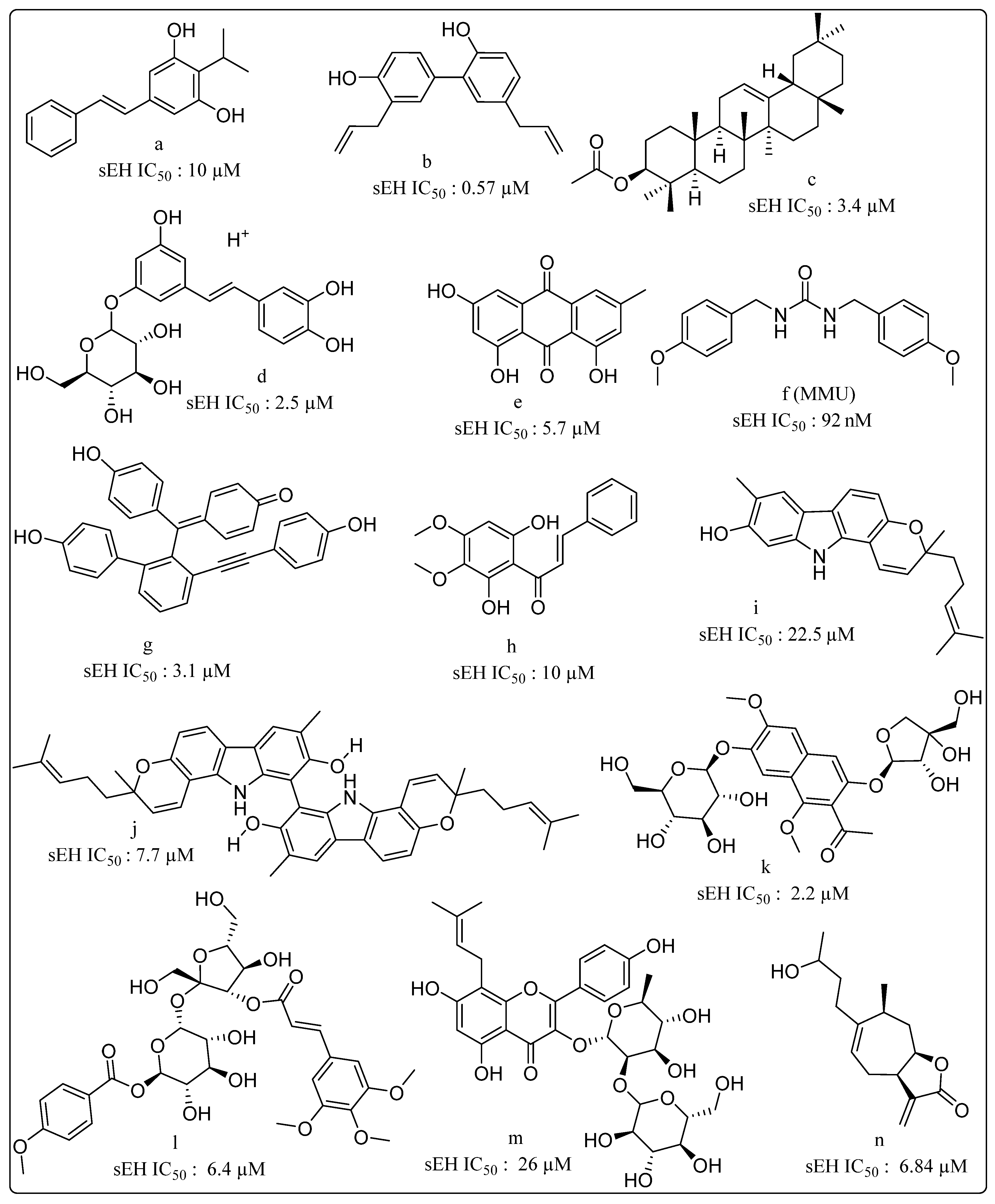{kind=link}
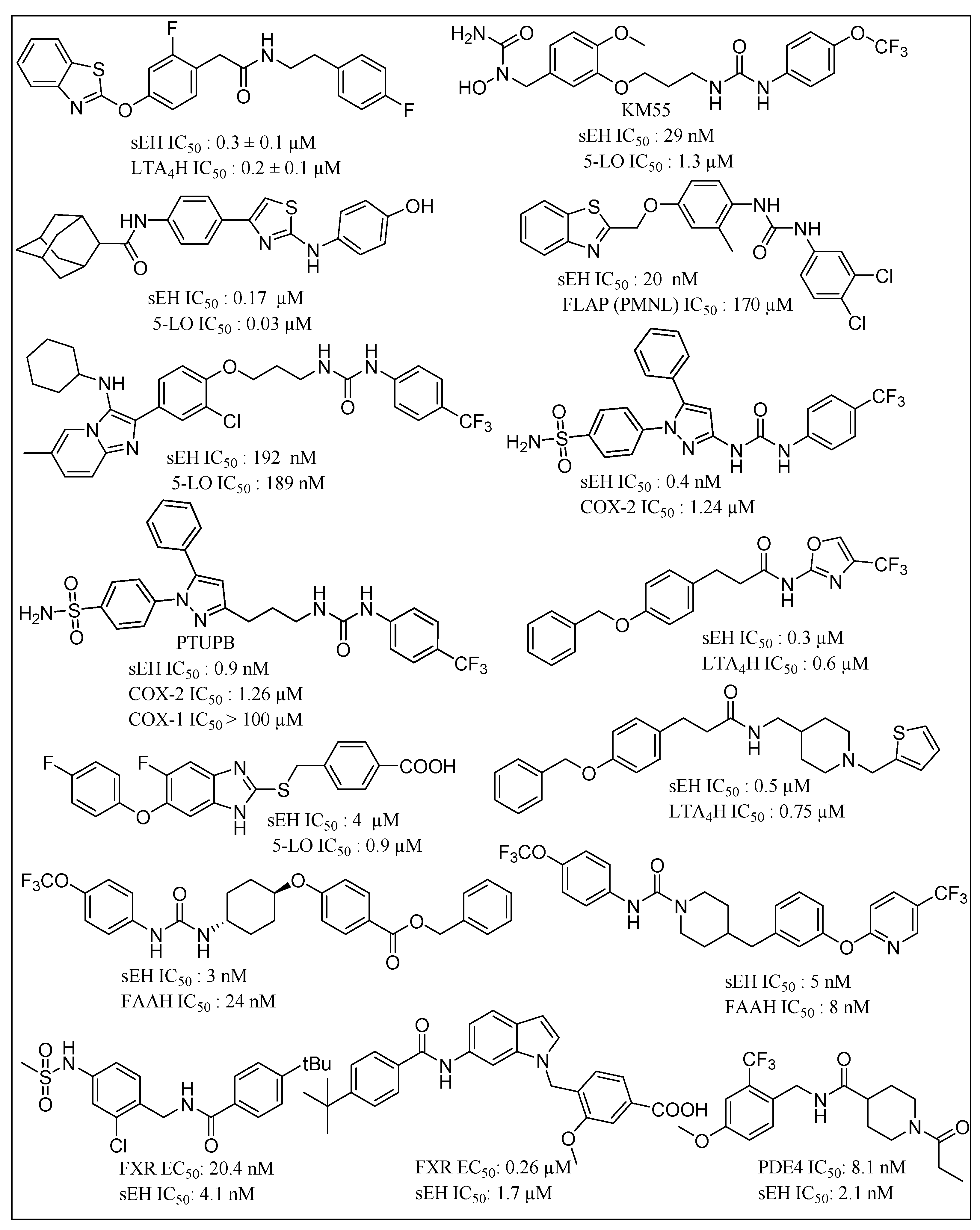{kind=link}
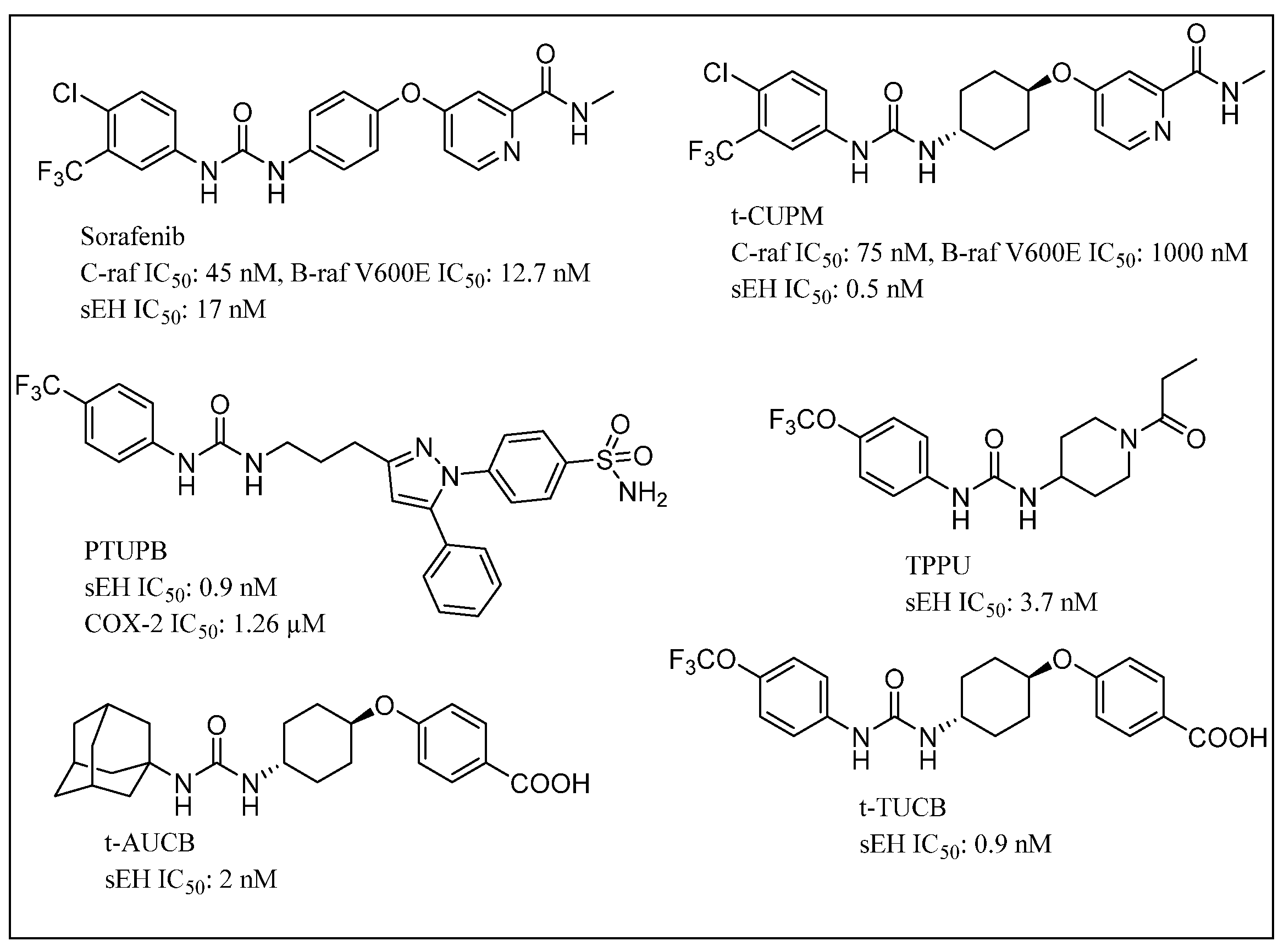{kind=link}
Abstract
1. Arachidonic Acid Pathway and Epoxyeicosatrienoic Acids (EETs)
2. Soluble Epoxide Hydrolase (sEH)
3. Role of EETs in Angiogenesis and Inflammation
4. sEH in Inflammation-Driven Carcinogenesis
5. Soluble Epoxide Hydrolase Inhibitors (sEHIs)
6. Dual Inhibition/Modulation of sEH as Part of Anti-Inflammatory Therapeutics
7. Dual Inhibition of sEH in Combination Chemotherapy
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| sEH | soluble epoxy hydrolase |
| EET | epoxyeicosatrienoic acid |
| ARA | arachidonic acid |
| COX | cyclooxygenase |
| LOX | lipoxygenase |
| 5-LO | 5-lipoxygenase |
| CYP | cytochrome P450 |
| HETEs | hydroxyeicosatetraenoic acids |
| LTA4 | leukotriene A4 |
| hsEH | human soluble epoxy hydrolase |
| DHETs | dihydroxyeicosatrienoic acid |
| Sphk1 | Sphingosine kinase 1 |
| FLAP | 5-lipoxygenase-activating protein |
| KRAS | Ki-ras2 Kirsten rat sarcoma viral oncogene homolog |
| sEHIs | soluble epoxy hydrolase inhibitors |
| DCU | 1,3-Dicyclohexylurea |
| CDU | 1-cyclohexyl-3-dodecyl-urea |
| AUDA | 12-(3-adamantan-1-yl-ureido) dodecanoic acid |
| AEPU | 1-adamantanyl-3-(5-(2-(2-ethoxyethoxy)ethoxy)pentyl)urea |
| APAU | 1-(1-Acetypiperidin-4-yl)-3-adamantanylurea |
| TPPU | 1-(1-propanoylpiperidin-4-yl)-3-[4-(trifluoromethoxy)phenyl]urea |
| PTUPB | 4-(5-phenyl-3-(3-[3-(4-trifluoromethyl-phenyl)-ureido]-propyl)S-pyrazol-1-yl) benzenesulfonamide |
| t-AUCB | trans-4-[4-(3-Adamantan-1-yl-ureido)-cyclohexyloxy]-benzoic acid |
| t-TUCB | trans-4-[4-[3-(4-Trifluoromethoxy-phenyl)-ureido]-cyclohexyloxy]-benzoic acid |
| CPTU | 1-(1-(cyclopropanecarbonyl)piperidin-4-yl)-3-(4 (trifluoromethoxy)phenyl)urea |
| TPAU | (1-trifluoromethoxyphenyl-3-(1-acetylpiperidin-4-yl)urea |
| ACU | 1-adamantyl-3-cyclohexylurea |
| AMAU | N-((1-acetylpiperidin-4-yl)methyl)-N’-(adamant-1-yl)urea |
| CUPM | 5-[4-[3-(4-chloro-3-trifluoromethyl-phenyl)-ureido]-cyclohexyloxy]-pyridine-2-carboxylic acid methylamide |
References
- Needleman, P.; Truk, J.; Jakschik, B.A.; Morrison, A.R.; Lefkowith, J.B. Arachidonic acid metabolism. Annu. Rev. Biochem. 1986, 55, 69–102. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D. Epoxides and soluble epoxide hydrolase in cardiovascular physiology. Physiol. Rev. 2012, 92, 101–130. [Google Scholar] [CrossRef] [PubMed]
- Kulmatycki, K.M.; Jamali, F. Drug disease interactions: Role of inflammatory mediators in disease and variability in drug response. Inflammation 2005, 23, 24. [Google Scholar]
- Zhang, W.; Liao, J.; Li, H.; Dong, H.; Bai, H.; Yang, A.; Hammock, B.D.; Yang, G.Y. Reduction of inflammatory bowel disease-induced tumor development in IL-10 knockout mice with soluble epoxide hydrolase gene deficiency. Mol. Carcinog. 2013, 52, 726–738. [Google Scholar] [CrossRef]
- Norwood, S.; Liao, J.; Hammock, B.D.; Yang, G.Y. Epoxyeicosatrienoic acids and soluble epoxide hydrolase: Potential therapeutic targets for inflammation and its induced carcinogenesis. Am. J. Transl. Res. 2010, 2, 447. [Google Scholar]
- Spector, A.A.; Fang, X.; Snyder, G.D.; Weintraub, N.L. Epoxyeicosatrienoic acids (EETs): Metabolism and biochemical function. Prog. Lipid Res. 2004, 43, 55–90. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, D.; Dou, X.; Niu, N.; Huang, W.; Bai, J.; Zhang, G. Elevated 14, 15-epoxyeicosatrienoic acid by increasing of cytochrome P450 2C8, 2C9 and 2J2 and decreasing of soluble epoxide hydrolase associated with aggressiveness of human breast cancer. BMC Cancer 2014, 14, 841. [Google Scholar] [CrossRef]
- Panigrahy, D.; Edin, M.L.; Lee, C.R.; Huang, S.; Bielenberg, D.R.; Butterfield, C.E.; Barnes, C.M.; Mammoto, A.; Mammoto, T.; Luria, A.; et al. Epoxyeicosanoids stimulate multiorgan metastasis and tumor dormancy escape in mice. J. Clin. Investig. 2012, 122, 178–191. [Google Scholar] [CrossRef]
- Yang, S.; Lin, L.; Chen, J.-X.; Lee, C.R.; Seubert, J.M.; Wang, Y.; Wang, H.; Chao, Z.-R.; Tao, D.-D.; Gong, J.-P.; et al. Cytochrome P-450 epoxygenases protect endothelial cells from apoptosis induced by tumor necrosis factor-alpha via MAPK and PI3K/Akt signaling pathways. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H142–H151. [Google Scholar] [CrossRef]
- Jiang, J.-G.; Chen, C.-L.; Card, J.W.; Yang, S.; Chen, J.-X.; Fu, X.-N.; Ning, Y.-G.; Xiao, X.; Zeldin, D.C.; Wang, D.W. Cytochrome P450 2J2 promotes the neoplastic phenotype of carcinoma cells and is up-regulated in human tumors. Cancer Res. 2005, 65, 4707–4715. [Google Scholar] [CrossRef]
- Newman, J.W.; Morisseau, C.; Hammock, B.D. Epoxide hydrolases: Their roles and interactions with lipid metabolism. Prog. Lipid Res. 2005, 44, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Hammock, B.D.; Grant, D.; Storms, D. Epoxide hydrolases. Compr. Toxicol. 1997, 3, 283–305. [Google Scholar]
- Fretland, A.J.; Omiecinski, C.J. Epoxide hydrolases: Biochemistry and molecular biology. Chem. Biol. Interact. 2000, 129, 41–59. [Google Scholar] [CrossRef]
- Shen, H.C.; Hammock, B.D. Discovery of inhibitors of soluble epoxide hydrolase: A target with multiple potential therapeutic indications. J. Med. Chem. 2012, 55, 1789–1808. [Google Scholar] [CrossRef]
- Gomez, G.A.; Morisseau, C.; Hammock, B.D.; Christianson, D.W. Structure of human epoxide hydrolase reveals inferences on bifunctional catalysis in epoxide and phosphate ester hydrolysis. Biochemistry 2004, 43, 4716–4723. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Xu, F.; Huse, L.M.; Morisseau, C.; Draper, A.J.; Newman, J.W.; Parker, C.; Graham, L.; Engler, M.M.; Hammock, B.D.; et al. Soluble epoxide hydrolase regulates hydrolysis of vasoactive epoxyeicosatrienoic acids. Circ. Res. 2000, 87, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Borhan, B.; Jones, A.D.; Pinot, F.; Grant, D.F.; Kurth, M.J.; Hammock, B.D. Mechanism of Soluble Epoxide Hydrolase formation of an α-hydroxy ester-enzyme intermediate through Asp-333. J. Biol. Chem. 1995, 270, 26923–26930. [Google Scholar] [CrossRef]
- Argiriadi, M.A.; Morisseau, C.; Hammock, B.D.; Christianson, D.W. Detoxification of environmental mutagens and carcinogens: Structure, mechanism, and evolution of liver epoxide hydrolase. Proc. Natl. Acad. Sci. USA 1999, 96, 10637–10642. [Google Scholar] [CrossRef]
- Yamada, T.; Morisseau, C.; Maxwell, J.E.; Argiriadi, M.A.; Christianson, D.W.; Hammock, B.D. Biochemical evidence for the involvement of tyrosine in epoxide activation during the catalytic cycle of epoxide hydrolase. J. Biol. Chem. 2000, 275, 23082–23088. [Google Scholar] [CrossRef]
- Morisseau, C.; Hammock, B.D. Epoxide hydrolases: Mechanisms, inhibitor designs, and biological roles. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 311–333. [Google Scholar] [CrossRef]
- Schiøtt, B.; Bruice, T.C. Reaction mechanism of soluble epoxide hydrolase: Insights from molecular dynamics simulations. J. Am. Chem. Soc. 2002, 124, 14558–14570. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.W.; Subrahmanyam, R.M.; Summers, S.A.; Xiao, X.; Alkayed, N.J. Soluble epoxide hydrolase dimerization is required for hydrolase activity. J. Biol. Chem. 2013, 288, 7697–7703. [Google Scholar] [CrossRef] [PubMed]
- Node, K.; Huo, Y.; Ruan, X.; Yang, B.; Spiecker, M.; Ley, K.; Zeldin, D.C.; Liao, J.K. Anti-inflammatory properties of cytochrome P450 epoxygenase-derived eicosanoids. Science 1999, 285, 1276–1279. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Norris, A.W. Action of epoxyeicosatrienoic acids on cellular function. Am. J. Physiol. Cell Physiol. 2007, 292, C996–C1012. [Google Scholar] [CrossRef] [PubMed]
- Panigrahy, D.; Greene, E.R.; Pozzi, A.; Wang, D.W.; Zeldin, D.C. EET signaling in cancer. Cancer Metastasis Rev. 2011, 30, 525–540. [Google Scholar] [CrossRef]
- Fleming, I.; Busse, R. Endothelium-derived epoxyeicosatrienoic acids and vascular function. Hypertension 2006, 47, 629–633. [Google Scholar] [CrossRef]
- Lu, T.; Hoshi, T.; Weintraub, N.L.; Spector, A.A.; Lee, H.C. Activation of ATP-sensitive K+ channels by epoxyeicosatrienoic acids in rat cardiac ventricular myocytes. J. Physiol. 2001, 53, 811–827. [Google Scholar] [CrossRef]
- Michaelis, U.R.; Fisslthaler, B.; Barbosa-Sicard, E.; Falck, J.R.; Fleming, I.; Busse, R. Cytochrome P450 epoxygenases 2C8 and 2C9 are implicated in hypoxia-induced endothelial cell migration and angiogenesis. J. Cell Sci. 2005, 118, 5489–5498. [Google Scholar] [CrossRef]
- Michaelis, U.R.; Fleming, I. From endothelium-derived hyperpolarizing factor (EDHF) to angiogenesis: Epoxyeicosatrienoic acids (EETs) and cell signaling. Pharmacol. Ther. 2006, 111, 584–595. [Google Scholar] [CrossRef]
- Michaelis, U.R.; Falck, J.R.; Schmidt, R.; Busse, R.; Fleming, I. Cytochrome P4502C9-derived epoxyeicosatrienoic acids induce the expression of cyclooxygenase-2 in endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 321–326. [Google Scholar] [CrossRef]
- Pozzi, A.; Macias-Perez, I.; Abair, T.; Wei, S.; Su, Y.; Zent, R.; Falck, J.R.; Capdevila, J.H. Characterization of 5, 6-and 8, 9-epoxyeicosatrienoic acids (5, 6-and 8, 9-EET) as potent in vivo angiogenic lipids. J. Biol. Chem. 2005, 280, 27138–27146. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Liu, Y.; Xie, L.; Wu, X.; Qiu, L.; Di, W. Sphingosine kinase 1/sphingosine-1-phosphate (S1P)/S1P receptor axis is involved in ovarian cancer angiogenesis. Oncotarget 2017, 8, 74947. [Google Scholar] [CrossRef] [PubMed]
- Pyne, N.J.; Pyne, S. Sphingosine-1-phosphate and cancer. Nat. Rev. Cancer 2010, 10, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Chen, S.; You, B.; Sun, J. Activation of sphingosine kinase-1 mediates induction of endothelial cell proliferation and angiogenesis by epoxyeicosatrienoic acids. Cardiovasc. Res. 2008, 78, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Su, K.H.; Lee, K.I.; Shyue, S.K.; Chen, H.Y.; Wei, J.; Lee, T.S. Implication of transient receptor potential vanilloid type 1 in 14, 15-epoxyeicosatrienoic acid-induced angiogenesis. Int. J. Biol. Sci. 2014, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- Lou, C.; Zhang, L.; Wang, X.; Ma, X.; Qin, C.; Li, W.; Jia, T.; Nan, Q.; Qiang, R. Soluble epoxide hydrolase: Potential target for inflammation and inflammation-driven cancer. J. Carcinog. Mutagen. 2017, 8, 1000294. [Google Scholar] [CrossRef]
- Imig, J.D.; Zhao, X.; Capdevila, J.H.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase inhibition lowers arterial blood pressure in angiotensin II hypertension. Hypertension 2002, 39, 690–694. [Google Scholar] [CrossRef]
- Chiamvimonvat, N.; Ho, C.M.; Tsai, H.J.; Hammock, B.D. The soluble epoxide hydrolase as a pharmaceutical target for hypertension. J. Cardiovasc. Pharmacol. 2007, 50, 225–237. [Google Scholar] [CrossRef]
- Li, J.; Carroll, M.A.; Chander, P.N.; Falck, J.R.; Sangras, B.; Stier, C.T. Soluble epoxide hydrolase inhibitor, AUDA, prevents early salt-sensitive hypertension. Front. Biosci. 2008, 13, 3480–3487. [Google Scholar] [CrossRef][Green Version]
- Liu, J.Y.; Park, S.H.; Morisseau, C.; Hwang, S.H.; Hammock, B.D.; Weiss, R.H. Sorafenib has soluble epoxide hydrolase inhibitory activity, which contributes to its effect profile In Vivo. Mol. Cancer Ther. 2009, 8, 2193–2203. [Google Scholar] [CrossRef]
- Zhang, L.N.; Vincelette, J.; Cheng, Y.; Mehra, U.; Chen, D.; Anandan, S.K.; Gless, R.; Webb, H.K.; Wang, Y.X. Inhibition of soluble epoxide hydrolase attenuated atherosclerosis, abdominal aortic aneurysm formation, and dyslipidemia. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Inceoglu, B.; Jinks, S.L.; Schmelzer, K.R.; Waite, T.; Kim, I.H.; Hammock, B.D. Inhibition of soluble epoxide hydrolase reduces LPS-induced thermal hyperalgesia and mechanical allodynia in a rat model of inflammatory pain. Life Sci. 2006, 79, 2311–2319. [Google Scholar] [CrossRef] [PubMed]
- Inceoglu, B.; Jinks, S.L.; Ulu, A.; Hegedus, C.M.; Georgi, K.; Schmelzer, K.R.; Wagner, K.; Jones, P.D.; Morisseau, C.; Hammock, B.D. Soluble epoxide hydrolase and epoxyeicosatrienoic acids modulate two distinct analgesic pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 18901–18906. [Google Scholar] [CrossRef] [PubMed]
- Rose, T.E.; Morisseau, C.; Liu, J.Y.; Inceoglu, B.; Jones, P.D.; Sanborn, J.R.; Hammock, B.D. 1-Aryl-3-(1-acylpiperidin-4-yl) urea inhibitors of human and murine soluble epoxide hydrolase: Structure-Activity relationships, pharmacokinetics, and reduction of inflammatory pain. J. Med. Chem. 2010, 53, 7067–7075. [Google Scholar] [CrossRef]
- Wagner, K.M.; McReynolds, C.B.; Schmidt, W.K.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for pain, inflammatory and neurodegenerative diseases. Pharmacol. Ther. 2017, 180, 62–76. [Google Scholar] [CrossRef]
- Wagner, K.; Inceoglu, B.; Gill, S.S.; Hammock, B.D. Epoxygenated fatty acids and soluble epoxide hydrolase inhibition: Novel mediators of pain reduction. J. Agric. Food Chem. 2011, 59, 2816–2824. [Google Scholar] [CrossRef][Green Version]
- Ma, M.; Ren, Q.; Yang, J.; Zhang, K.; Xiong, Z.; Ishima, T.; Pu, Y.; Hwang, S.H.; Toyoshima, M.; Iwayama, Y.; et al. Key role of soluble epoxide hydrolase in the neurodevelopmental disorders of offspring after maternal immune activation. Proc. Natl. Acad. Sci. USA 2019, 116, 7083–7088. [Google Scholar] [CrossRef]
- Ren, Q.; Ma, M.; Yang, J.; Nonaka, R.; Yamaguchi, A.; Ishikawa, K.I.; Kobayashi, K.; Murayama, S.; Hwang, S.H.; Saiki, S.; et al. Soluble epoxide hydrolase plays a key role in the pathogenesis of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2018, 115, E5815–E5823. [Google Scholar] [CrossRef]
- Hashimoto, K. Role of soluble epoxide hydrolase in metabolism of PUFAs in psychiatric and neurological disorders. Front. Pharmacol. 2019, 10, 36. [Google Scholar] [CrossRef]
- Zarriello, S.; Tuazon, J.P.; Corey, S.; Schimmel, S.; Rajani, M.; Gorsky, A.; Incontri, D.; Hammock, B.D.; Borlongan, C.V. Humble beginnings with big goals: Small molecule soluble epoxide hydrolase inhibitors for treating CNS disorders. Prog. Neurobiol. 2019, 172, 23–39. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, Y.; Fan, Z.; Chen, Q.; Chen, J.; Sun, Y.; Jiang, X.; Xiao, Q. Soluble epoxide hydrolase inhibitor protects against blood-brain barrier dysfunction in a mouse model of type 2 diabetes via the AMPK/HO-1 pathway. Biochem. Biophys. Res. Commun. 2020, 524. [Google Scholar] [CrossRef] [PubMed]
- Savina, Y.; Duflot, T.; Bounoure, F.; Kotzki, S.; Thiébaut, P.A.; Serreau, P.A.; Skiba, M.; Picquenot, J.M.; Cornic, M.; Morisseau, C.; et al. Impact of the acute local inhibition of soluble epoxide hydrolase on diabetic skin microcirculatory dysfunction. Diabetes Vasc. Dis. Res. 2019, 16, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Minaz, N.; Razdan, R.; Hammock, B.D.; Mujwar, S.; Goswami, S.K. Impact of diabetes on male sexual function in streptozotocin-induced diabetic rats: Protective role of soluble epoxide hydrolase inhibitor. Biomed. Pharmacother. 2019, 115, 108897. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Dziumbla, S.; Lin, J.; Bibli, S.I.; Zukunft, S.; De Mos, J.; Awwad, K.; Frömel, T.; Jungmann, A.; Devraj, K.; et al. Inhibition of soluble epoxide hydrolase prevents diabetic retinopathy. Nature 2017, 552, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Park, B.; Corson, T.W. Soluble epoxide hydrolase inhibition for ocular diseases: Vision for the future. Front. Pharmacol. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, R.S.; Park, B.; Sheik Pran Babu, S.P.; Si, Y.; Kharwadkar, R.; Mitter, S.K.; Lee, B.; Sun, W.; Qi, X.; Boulton, M.E.; et al. Chemical proteomics reveals soluble epoxide hydrolase as a therapeutic target for ocular neovascularization. ACS Chem. Biol. 2018, 13, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.H.; Park, Y.K.; Hammock, B.D.; Nishi, K. Structure-Activity relationships of cycloalkylamide derivatives as inhibitors of the soluble epoxide hydrolase. J. Med. Chem. 2011, 54, 1752–1761. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gomez, G.A.; Morisseau, C.; Hammock, B.D.; Christianson, D.W. Human soluble epoxide hydrolase: Structural basis of inhibition by 4-(3-cyclohexylureido)-carboxylic acids. Protein Sci. 2006, 15, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Eldrup, A.B.; Soleymanzadeh, F.; Taylor, S.J.; Muegge, I.; Farrow, N.A.; Joseph, D.; McKellop, K.; Man, C.C.; Kukulka, A.; De Lombaert, S. Structure-Based optimization of arylamides as inhibitors of soluble epoxide hydrolase. J. Med. Chem. 2009, 52, 5880–5895. [Google Scholar] [CrossRef] [PubMed]
- Argiriadi, M.A.; Morisseau, C.; Goodrow, M.H.; Dowdy, D.L.; Hammock, B.D.; Christianson, D.W. Binding of alkylurea inhibitors to epoxide hydrolase implicates active site tyrosines in substrate activation. J. Biol. Chem. 2000, 275, 15265–15270. [Google Scholar] [CrossRef]
- Xing, L.; McDonald, J.J.; Kolodziej, S.A.; Kurumbail, R.G.; Williams, J.M.; Warren, C.J.; O’Neal, J.M.; Skepner, J.E.; Roberds, S.L. Discovery of potent inhibitors of soluble epoxide hydrolase by combinatorial library design and structure-based virtual screening. J. Med. Chem. 2011, 54, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Morisseau, C.; Pakhomova, S.; Hwang, S.H.; Newcomer, M.E.; Hammock, B.D. Inhibition of soluble epoxide hydrolase by fulvestrant and sulfoxides. Bioorg. Med. Chem. Lett. 2013, 23, 3818–3821. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dietze, E.C.; Kuwano, E.; Casas, J.; Hammock, B.D. Inhibition of cytosolic epoxide hydrolase by trans-3-phenylglycidols. Biochem. Pharmacol. 1991, 42, 1163–1175. [Google Scholar] [CrossRef]
- Morisseau, C.; Du, G.; Newman, J.W.; Hammock, B.D. Mechanism of mammalian soluble epoxide hydrolase inhibition by chalcone oxide derivatives. Arch. Biochem. Biophys. 1998, 356, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Asokkumar, K.; Prathyusha, L.T.; Umamaheshwari, M.; Sivashanmugam, T.; Subhadradevi, V.; Jagannath, P.; Madeswaran, A.; Salesheir, F. Design, ADMET and docking studies on some novel chalcone derivatives as soluble epoxide hydrolase enzyme inhibitors. J. Chil. Chem. Soc. 2012, 57, 1442–1446. [Google Scholar] [CrossRef][Green Version]
- Huang, S.-X.; Cao, B.; Morisseau, C.; Jin, Y.; Hammock, B.D.; Long, Y.-Q. Structure-based optimization of the piperazino-containing 1,3-disubstituted ureas affording sub-nanomolar inhibitors of soluble epoxide hydrolase. Med. Chem. Commun. 2012, 3, 379–384. [Google Scholar] [CrossRef][Green Version]
- Jones, P.D.; Hsing-Ju, T.; Zung, N.D.; Morisseau, C.; Hammock, B.D. Synthesis and SAR of conformationally restricted inhibitors of soluble epoxide hydrolase. Bioorg. Med. Chem. Lett. 2006, 16, 5212–5216. [Google Scholar] [CrossRef][Green Version]
- Jones, P.D.; Wolf, N.M.; Morisseau, C.; Whetstone, P.; Hock, B.; Hammock, B.D. Fluorescent substrates for soluble epoxide hydrolase and application to inhibition studies. Anal. Biochem. 2005, 343, 66–75. [Google Scholar] [CrossRef]
- Morisseau, C.; Goodrow, M.H.; Newman, J.W.; Wheelock, C.E.; Dowdy, D.L.; Hammock, B.D. Structural refinement of inhibitors of urea-based soluble epoxide hydrolases. Biochem. Pharmacol. 2002, 63, 1599–1608. [Google Scholar] [CrossRef]
- Ulu, A.; Appt, S.; Morisseau, C.; Hwang, S.H.; Jones, P.D.; Rose, T.E.; Dong, H.; Lango, J.; Yang, J.; Tsai, H.J.; et al. Pharmacokinetics and in vivo potency of soluble epoxide hydrolase inhibitors in cynomolgus monkeys. Br. J. Pharmacol. 2012, 165, 1401–1412. [Google Scholar] [CrossRef]
- Watanabe, T.; Morisseau, C.; Newman, J.W.; Hammock, B.D. In Vitro metabolism of the mammalian soluble epoxide hydrolase inhibitor, 1-cyclohexyl-3-dodecyl-urea. Drug Metab. Dispos. 2003, 31, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Olearczyk, J.J.; Field, M.B.; Kim, I.H.; Morisseau, C.; Hammock, B.D.; Imig, J.D. Substituted adamantyl-urea inhibitors of the soluble epoxide hydrolase dilate mesenteric resistance vessels. J. Pharmacol. Exp. Ther. 2006, 318, 1307–1314. [Google Scholar] [CrossRef]
- Kim, I.H.; Morisseau, C.; Watanabe, T.; Hammock, B.D. Design, synthesis, and biological activity of 1, 3-disubstituted ureas as potent inhibitors of the soluble epoxide hydrolase of increased water solubility. J. Med. Chem. 2004, 47, 2110–2122. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.J.; Hwang, S.H.; Morisseau, C.; Yang, J.; Jones, P.D.; Kasagami, T.; Kim, I.H.; Hammock, B.D. Pharmacokinetic screening of soluble epoxide hydrolase inhibitors in dogs. Eur. J. Pharm. Sci. 2010, 40, 222–238. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wan, D.; Yang, J.; McReynolds, C.B.; Barnych, B.; Wagner, K.M.; Morisseau, C.; Hwang, S.H.; Sun, J.; Blöcher, R.; Hammock, B.D. In vitro and in vivo metabolism of a potent inhibitor of soluble epoxide hydrolase, 1-(1-propionylpiperidin-4-yl)-3-(4-(trifluoromethoxy) phenyl) urea. Front. Pharmacol. 2019, 10, 464. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Whitcomb, R.; MacIntyre, E.; Tran, V.; Do, Z.N.; Sabry, J.; Patel, D.V.; Anandan, S.K.; Gless, R.; Webb, H.K. Pharmacokinetics and pharmacodynamics of AR9281, an inhibitor of soluble epoxide hydrolase, in single- and multiple-dose studies in healthy human subjects. J. Clin. Pharmacol. 2012, 52, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Sun, G.Y.; Liu, T.; Duan, J.X.; Zhou, H.F.; Lee, K.S.; Hammock, B.D.; Fang, X.; Jiang, J.X.; Guan, C.X. Soluble epoxide hydrolase inhibitor 1-trifluoromethoxyphenyl-3-(1-propionylpiperidin-4-yl) urea attenuates bleomycin-induced pulmonary fibrosis in mice. Cell Tissue Res. 2016, 363, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Eldrup, A.B.; Soleymanzadeh, F.; Farrow, N.A.; Kukulka, A.; De Lombaert, S. Optimization of piperidyl-ureas as inhibitors of soluble epoxide hydrolase. Bioorg. Med. Chem. Lett. 2010, 20, 571–575. [Google Scholar] [CrossRef]
- Shen, H.C.; Ding, F.X.; Wang, S.; Deng, Q.; Zhang, X.; Chen, Y.; Zhou, G.; Xu, S.; Chen, H.S.; Tong, X.; et al. Discovery of a highly potent, selective, and bioavailable soluble epoxide hydrolase inhibitor with excellent ex vivo target engagement. J. Med. Chem. 2009, 52, 5009–5012. [Google Scholar] [CrossRef]
- Shen, H.C.; Ding, F.X.; Wang, S.; Xu, S.; Chen, H.S.; Tong, X.; Tong, V.; Mitra, K.; Kumar, S.; Zhang, X.; et al. Discovery of spirocyclic secondary amine-derived tertiary ureas as highly potent, selective and bioavailable soluble epoxide hydrolase inhibitors. Bioorg. Med. Chem. Lett. 2009, 19, 3398–3404. [Google Scholar] [CrossRef]
- Burmistrov, V.V.; Butov, G.M. Synthesis and properties of N-[R-adamantan-1 (2)-yl]-N′-(2-fluorophenyl) ureas—Target-Oriented soluble epoxide hydrolase inhibitors. Russ. J. Org. Chem. 2018, 54, 1307–1312. [Google Scholar] [CrossRef]
- Burmistrov, V.V.; Morisseau, C.; Karlov, D.; Pitushkin, D.; Vernigora, A.; Rasskazova, E.; Butov, G.M.; Hammock, B.D. Bioisosteric substitution of adamantane with bicyclic lipophilic groups improves water solubility of human soluble epoxide hydrolase inhibitors. Bioorg. Med. Chem. Lett. 2020, 30, 127430. [Google Scholar] [CrossRef] [PubMed]
- Codony, S.; Pujol, E.; Pizarro, J.; Feixas, F.; Valverde, E.; Loza, M.I.; Brea, J.M.; Saez, E.; Oyarzabal, J.; Pineda-Lucena, A.; et al. 2-Oxaadamant-1-yl Ureas as soluble epoxide hydrolase inhibitors: In Vivo evaluation in a murine model of acute pancreatitis. J. Med. Chem. 2020, 63, 9237–9257. [Google Scholar] [CrossRef] [PubMed]
- Waltenberger, B.; Garscha, U.; Temml, V.; Liers, J.; Werz, O.; Schuster, D.; Stuppner, H. Discovery of potent soluble epoxide hydrolase (sEH) inhibitors by pharmacophore-based virtual screening. J. Chem. Inf. Model. 2016, 56, 747–762. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, E.; Hedayati, M.; Rad, L.H.; Shahhosseini, S.; Faizi, M.; Tabatabai, S.A. Novel soluble epoxide hydrolase inhibitors with a dihydropyrimidinone scaffold: Design, synthesis and biological evaluation. Med. Chem. Commun. 2016, 7, 2128–2135. [Google Scholar] [CrossRef]
- Lazaar, A.L.; Yang, L.; Boardley, R.L.; Goyal, N.S.; Robertson, J.; Baldwin, S.J.; Newby, D.E.; Wilkinson, I.B.; Tal-Singer, R.; Mayer, R.J.; et al. Pharmacokinetics, pharmacodynamics and adverse event profile of GSK2256294, a novel soluble epoxide hydrolase inhibitor. Br. J. Clin. Pharmacol. 2016, 81, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Paliwal, S.; Sharma, S.; Verma, K.; Gururani, R.; Tiwari, A.; Verma, A.; Chauhan, M.; Singh, A.; Kumar, D.; et al. Discovery of novel soluble epoxide hydrolase inhibitors as potent vasodilators. Sci. Rep. 2018, 8, 14604. [Google Scholar] [CrossRef]
- Batchu, S.N.; Lee, S.B.; Samokhvalov, V.; Chaudhary, K.R.; El-Sikhry, H.; Weldon, S.M.; Seubert, J.M. Novel soluble epoxide hydrolase inhibitor protects mitochondrial function following stress. Can. J. Physiol. Pharmacol. 2012, 90, 811–823. [Google Scholar] [CrossRef]
- Kim, I.H.; Heirtzler, F.R.; Morisseau, C.; Nishi, K.; Tsai, H.J.; Hammock, B.D. Optimization of amide-based inhibitors of soluble epoxide hydrolase with improved water solubility. J. Med. Chem. 2005, 48, 3621–3629. [Google Scholar] [CrossRef]
- Anandan, S.K.; Webb, H.K.; Do, Z.N.; Gless, R.D. Unsymmetrical non-adamantyl N, N′-diaryl urea and amide inhibitors of soluble expoxide hydrolase. Bioorg. Med. Chem. Lett. 2009, 19, 4259–4263. [Google Scholar] [CrossRef]
- Rezaee, E.; Amrolah, S.M.; Nazari, M.; Tabatabi, S.A. Novel amide derivatives of 3-phenylglutaric acid as potent soluble epoxide hydrolase inhibitors. Mol. Divers. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pecic, S.; Deng, S.-X.; Morisseau, C.; Hammock, B.D.; Landry, D.W. Design, synthesis and evaluation of non-urea inhibitors of soluble epoxide hydrolase. Bioorg. Med. Chem. Lett. 2012, 22, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, Y.; Gong, G.; Smith, D.H.; Yan, F.; Rinderspacher, A.; Feng, Y.; Zhu, Z.; Li, X.; Deng, S.X.; et al. Discovery of potent non-urea inhibitors of soluble epoxide hydrolase. Bioorg. Med. Chem. Lett. 2009, 19, 2354–2359. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pecic, S.; Pakhomova, S.; Newcomer, M.E.; Morisseau, C.; Hammock, B.D.; Zhu, Z.; Rinderspacher, A.; Deng, S.-X. Synthesis and structure-activity relationship of piperidine-derived non-urea soluble epoxide hydrolase inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.C.; Ding, F.X.; Deng, Q.; Xu, S.; Tong, X.; Zhang, X.; Chen, Y.; Zhou, G.; Pai, L.Y.; Alonso-Galicia, M.; et al. A strategy of employing aminoheterocycles as amide mimics to identify novel, potent and bioavailable soluble epoxide hydrolase inhibitors. Bioorg. Med. Chem. Lett. 2009, 19, 5716–5721. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Ma, W.H.; Ma, Y.L.; Ban, S.R.; Feng, X.E.; Li, Q.S. Synthesis and biological activity of 4-substituted benzoxazolone derivatives as a new class of sEH inhibitors with high anti-inflammatory activity in vivo. Bioorg. Med. Chem. Lett. 2013, 23, 2380–2383. [Google Scholar] [CrossRef]
- Schmelzer, K.R.; Kubala, L.; Newman, J.W.; Kim, I.H.; Eiserich, J.P.; Hammock, B.D. Soluble epoxide hydrolase is a therapeutic target for acute inflammation. Proc. Natl. Acad. Sci. USA 2005, 102, 9772–9777. [Google Scholar] [CrossRef]
- Burmistrov, V.V.; Morisseau, C.; Pitushkin, D.; Karlov, D.; Fayzullin, R.R.; Butov, G.M.; Hammock, B.D. Adamantyl thioureas as soluble epoxide hydrolase inhibitors. Bioorg. Med. Chem. Lett. 2018, 28, 2302–2313. [Google Scholar] [CrossRef]
- Bhagwati, S.; Siddiqui, M.I. Identification of potential soluble epoxide hydrolase (sEH) inhibitors by ligand-based pharmacophore model and biological evaluation. J. Biomol. Struct. Dyn. 2019, 38, 4956–4966. [Google Scholar] [CrossRef]
- Tanaka, D.; Tsuda, Y.; Shiyama, T.; Nishimura, T.; Chiyo, N.; Tominaga, Y.; Sawada, N.; Mimoto, T.; Kusunose, N. A practical use of ligand efficiency indices out of the fragment-based approach: Ligand efficiency-guided lead identification of soluble epoxide hydrolase inhibitors. J. Med. Chem. 2011, 54, 851–857. [Google Scholar] [CrossRef]
- Amano, Y.; Yamaguchi, T.; Tanabe, E. Structural insights into binding of inhibitors to soluble epoxide hydrolase gained by fragment screening and X-ray crystallography. Bioorg. Med. Chem. 2014, 22, 2427–2434. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.S.; Liu, J.-Y.; Wagner, K.M.; Pakhomova, S.; Dong, H.; Morisseau, C.; Fu, S.H.; Yang, J.; Wang, P.; Ulu, A.; et al. Optimized Inhibitors of soluble epoxide hydrolase improve in vitro target residence time and in vivo efficacy. J. Med. Chem. 2014, 57, 7016–7030. [Google Scholar] [CrossRef] [PubMed]
- Lukin, A.; Kramer, J.; Hartmann, M.; Weizel, L.; Hernandez-Olmos, V.; Falahati, K.; Burghardt, I.; Kalinchenkova, N.; Bagnyukova, D.; Zhurilo, N.; et al. Discovery of polar spirocyclic orally bioavailable urea inhibitors of soluble epoxide hydrolase. Bioorg. Chem. 2018, 80, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Thalji, R.K.; McAtee, J.J.; Belyanskaya, S.; Brandt, M.; Brown, G.D.; Costell, M.H.; Ding, Y.; Dodson, J.W.; Eisennagel, S.H.; Fries, R.E.; et al. Discovery of 1-(1, 3, 5-triazin-2-yl) piperidine-4-carboxamides as inhibitors of soluble epoxide hydrolase. Bioorg. Med. Chem. Lett. 2013, 23, 3584–3588. [Google Scholar] [CrossRef]
- Lo, H.Y.; Man, C.C.; Fleck, R.W.; Farrow, N.A.; Ingraham, R.H.; Kukulka, A.; Proudfoot, J.R.; Betageri, R.; Kirrane, T.; Patel, U.; et al. Substituted pyrazoles as novel sEH antagonist: Investigation of key binding interactions within the catalytic domain. Bioorg. Med. Chem. Lett. 2010, 20, 6379–6383. [Google Scholar] [CrossRef]
- Scholz, M.S.; Wingen, L.M.; Brunst, S.; Wittman, S.; Cardoso, I.L.A.; Weizel, L.; Proschak, E. Soluble epoxide hydrolase inhibitors with carboranes as non-natural 3-D pharmacophores. Eur. J. Med. Chem. 2020, 185, 111766. [Google Scholar] [CrossRef]
- Vinh, L.B.; Kim, J.H.; Lee, J.S.; Nguyet, N.T.M.; Yang, S.Y.; Ma, J.Y.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of phenolic glycosides from Polygala tenuifolia and in silico approach. Med. Chem. Res. 2018, 27, 726–734. [Google Scholar] [CrossRef]
- Liu, J.Y.; Morisseau, C.; Huang, H.; Hammock, B.D. Screening of soluble epoxide hydrolase inhibitory ingredients from traditional Chinese medicines for anti-inflammatory use. J. Ethnopharmacol. 2016, 194, 475–482. [Google Scholar] [CrossRef]
- Kitamura, S.; Morisseau, C.; Inceoglu, B.; Kamita, S.G.; De Nicola, G.R.; Nyegue, M.; Hammock, B.D. Potent natural soluble epoxide hydrolase inhibitors from Pentadiplandra brazzeana Baillon: Synthesis, quantification, and measurement of biological activities in vitro and in vivo. PLoS ONE 2015, 10, e0117438. [Google Scholar] [CrossRef]
- Thao, N.P.; Kim, J.H.; Luyen, B.T.T.; Dat, N.T.; Kim, Y.H. In silico investigation of cycloartane triterpene derivatives from Cimicifuga dahurica (Turcz.) Maxim. roots for the development of potent soluble epoxide hydrolase inhibitors. Int. J. Biol. Macromol. 2017, 98, 526–534. [Google Scholar] [CrossRef]
- Buscató, E.L.; Büttner, D.; Brüggerhoff, A.; Klingler, F.M.; Weber, J.; Scholz, B.; Živković, A.; Marschalek, R.; Stark, H.; Steinhilber, D.; et al. From a multipotent stilbene to soluble epoxide hydrolase inhibitors with antiproliferative properties. ChemMedChem 2013, 8, 919–923. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, I.S.; Ryu, J.; Lee, J.S.; Kang, J.S.; Kang, S.Y.; Kim, Y.H. In vitro and in silico investigation of anthocyanin derivatives as soluble epoxide hydrolase inhibitors. Int. J. Biol. Macromol. 2018, 112, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Oh, S.J.; Lee, S.Y.; Lee, Y.-J.; Ma, J.Y.; Kim, Y.H.; Kim, S.K. Discovery of soluble epoxide hydrolase inhibitors from natural products. Food Chem. Toxicol. 2014, 64, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Kim, J.H.; Choi, S.-K.; Kim, Y.H. Constituents of the seeds of Cassia tora with inhibitory activity on soluble epoxide hydrolase. Bioorg. Med. Chem. Lett. 2015, 25, 5097–5101. [Google Scholar] [CrossRef] [PubMed]
- Jo, A.R.; Kim, J.H.; Yan, X.-T.; Yang, S.Y.; Kim, Y.H. Soluble epoxide hydrolase inhibitory components from Rheum undulatum and in silico approach. J. Enzym. Inhib. Med. Chem. 2016, 31, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, C.W.; Tai, B.H.; Yang, S.Y.; Choi, G.-S.; Kang, J.S.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of selaginellin derivatives from Selaginella tamariscina. Molecules 2015, 20, 21405–21414. [Google Scholar] [CrossRef]
- Kim, J.H.; Morgan, A.M.A.; Tai, B.H.; Van, D.T.; Cuong, N.M.; Kim, Y.H. Inhibition of soluble epoxide hydrolase activity by compounds isolated from the aerial parts of Glycosmis stenocarpa. J. Enzym. Inhib. Med. Chem. 2016, 31, 640–644. [Google Scholar] [CrossRef]
- He, X.; Zhao, W.-Y.; Shao, B.; Zhang, B.-J.; Liu, T.-T.; Sun, C.-P.; Huang, H.-L.; Wu, J.-R.; Liang, J.-H.; Ma, X.-C. Natural soluble epoxide hydrolase inhibitors from Inula helenium and their interactions with soluble epoxide hydrolase. Int. J. Biol. Macromol. 2020, 158, 1362–1368. [Google Scholar] [CrossRef]
- Su, X.D.; Li, W.; Kim, J.E.; Yang, S.Y.; Ma, J.Y.; Kim, Y.H. Prenyl-flavonoids from Epimedium koreanum Nakai and their soluble epoxide hydrolase and tyrosinase inhibitory activities. Med. Chem. Res. 2017, 26, 2761–2767. [Google Scholar]
- Li, H.X.; Heo, M.; Go, Y.; Kim, Y.S.; Kim, Y.H.; Yang, S.Y.; Li, W. Coumarin and moracin derivatives from mulberry leaves (Morus alba L.) with soluble epoxide hydrolase inhibitory activity. Molecules 2020, 25, 3967. [Google Scholar] [CrossRef]
- Sun, C.-P.; Zhang, J.; Zhao, W.-Y.; Yi, J.; Yan, J.-K.; Wang, Y.-L.; Morisseau, C.; Liu, Z.-B.; Hammock, B.D.; Ma, X.-C. Protostane-Type triterpenoids as natural soluble epoxide hydrolase inhibitors: Inhibition potentials and molecular dynamics. Bioorg. Chem. 2020, 96, 103637. [Google Scholar] [CrossRef] [PubMed]
- Falck, J.R.; Kodela, R.; Manne, R.; Atcha, K.R.; Puli, N.; Dubasi, N.; Manthati, V.L.; Capdevila, J.H.; Yi, X.Y.; Goldman, D.H.; et al. 14,15-Epoxyeicosa-5,8,11-trienoic acid (14,15-EET) surrogates containing epoxide bioisosteres: Influence upon vascular relaxation and soluble epoxide hydrolase inhibition. J. Med. Chem. 2009, 52, 5069–5075. [Google Scholar] [CrossRef]
- Falck, J.R.; Koduru, S.R.; Mohapatra, S.; Manne, R.; Atcha, R.; Manthati, V.L.; Capdevila, J.H.; Christian, S.; Imig, J.D.; Campbell, W.B. 14,15-Epoxyeicosa-5,8,11-trienoic acid (14,15-EET) surrogates: Carboxylate modifications. J. Med. Chem. 2014, 57, 6965–6972. [Google Scholar] [CrossRef] [PubMed]
- Litovchick, A.; Dumelin, C.E.; Habeshian, S.; Gikunju, D.; Guie, M.A.; Centrella, P.; Zhang, Y.; Sigel, E.A.; Cuozzo, J.W.; Keefe, A.D.; et al. Encoded library synthesis using chemical ligation and the discovery of sEH inhibitors from a 334-million member library. Sci. Rep. 2015, 5, 10916. [Google Scholar] [CrossRef] [PubMed]
- Belyanskaya, S.L.; Ding, Y.; Callahan, J.F.; Lazaar, A.L.; Israel, D.I. Discovering drugs with DNA-encoded library technology: From concept to clinic with an inhibitor of soluble epoxide hydrolase. ChemBioChem 2017, 18, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Abis, G.; Charles, R.L.; Kopec, J.; Yue, W.W.; Atkinson, R.A.; Bui, T.T.; Lynham, S.; Popova, S.; Sun, Y.B.; Fraternali, F.; et al. 15-deoxy-Δ 12, 14-Prostaglandin J 2 inhibits human soluble epoxide hydrolase by a dual orthosteric and allosteric mechanism. Commun. Biol. 2019, 2, 188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Panigrahy, D.; Hwang, S.H.; Yang, J.; Mahakian, L.M.; Wettersten, H.I.; Liu, J.Y.; Wang, Y.; Ingham, E.S.; Tam, S.; et al. Dual inhibition of cyclooxygenase-2 and soluble epoxide hydrolase synergistically suppresses primary tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2014, 111, 11127–11132. [Google Scholar] [CrossRef]
- Jones, R.D.; Liao, J.; Tong, X.; Xu, D.; Sun, L.; Li, H.; Yang, G.Y. Epoxy-Oxylipins and soluble epoxide hydrolase metabolic pathway as targets for NSAID-induced gastroenteropathy and inflammation-associated carcinogenesis. Front. Pharmacol. 2019, 10, 731. [Google Scholar] [CrossRef]
- Schmelzer, K.R.; Inceoglu, B.; Kubala, L.; Kim, I.H.; Jinks, S.L.; Eiserich, J.P.; Hammock, B.D. Enhancement of antinociception by coadministration of nonsteroidal anti-inflammatory drugs and soluble epoxide hydrolase inhibitors. Proc. Natl. Acad. Sci. USA 2006, 103, 13646–13651. [Google Scholar] [CrossRef]
- Wagner, K.; Inceoglu, B.; Hammock, B.D. Soluble epoxide hydrolase inhibition, epoxygenated fatty acids and nociception. Prostaglandins Other Lipid Mediat. 2011, 96, 76–83. [Google Scholar] [CrossRef]
- Goswami, S.K.; Wan, D.; Yang, J.; Da Silva, C.A.T.; Morisseau, C.; Kodani, S.D.; Yang, G.Y.; Inceoglu, B.; Hammock, B.D. Anti-Ulcer efficacy of soluble epoxide hydrolase inhibitor TPPU on diclofenac-induced intestinal ulcers. J. Pharmacol. Exp. Ther. 2016, 357, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Wagner, K.M.; Morisseau, C.; Liu, J.Y.; Dong, H.; Wecksler, A.T.; Hammock, B.D. Synthesis and structure-activity relationship studies of urea-containing pyrazoles as dual inhibitors of cyclooxygenase-2 and soluble epoxide hydrolase. J. Med. Chem. 2011, 54, 3037–3050. [Google Scholar] [CrossRef] [PubMed]
- Abdelazeem, A.H.; Safi El-Din, A.G.; Abdel-Fattah, M.M.; Amin, N.H.; El-Moghazy, S.M.; El-Saadi, M.T. Discovery of novel urea-diarylpyrazole hybrids as dual COX-2/sEH inhibitors with improved anti-inflammatory activity and highly reduced cardiovascular risks. Eur. J. Med. Chem. 2020, 205, 112662. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-Y.; Yang, J.; Inceoglu, B.; Qiu, H.; Ulu, A.; Hwang, S.-H.; Chiamvimonvat, N.; Hammock, B.D. Inhibition of soluble epoxide hydrolase enhances the anti-inflammatory effects of aspirin and 5-lipoxygenase activation protein inhibitor in a murine model. Biochem. Pharmacol. 2010, 79, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Garscha, U.; Romp, E.; Pace, S.; Rossi, A.; Temml, V.; Schuster, D.; Konig, S.; Gerstmeier, J.; Liening, S.; Werner, M.; et al. Pharmacological profile and efficiency in vivo of diflapolin, the first dual inhibitor of 5-lipoxygenase-activating protein and soluble epoxide hydrolase. Sci. Rep. 2017, 7, 9398. [Google Scholar] [CrossRef]
- Temml, V.; Garscha, U.; Romp, E.; Schubert, G.; Gerstmeier, J.; Kutil, Z.; Matuszczak, B.; Waltenberger, B.; Stuppner, H.; Werz, O.; et al. Discovery of the first dual inhibitor of the 5-lipoxygenase-activating protein and soluble epoxide hydrolase using pharmacophore-based virtual screening. Sci. Rep. 2017, 7, 42751. [Google Scholar] [CrossRef]
- Hiesinger, K.; Schott, A.; Kramer, J.S.; Blocher, R.; Witt, F.; Wittman, S.K.; Steinhilber, D.; Pogoryelov, D.; Gerstmeier, J.; Werz, O.; et al. Design of dual inhibitors of soluble epoxide hydrolase and LTA4 hydrolase. ACS Med. Chem. Lett. 2020, 11, 298–302. [Google Scholar] [CrossRef]
- Hefke, L.; Hiesinger, K.; Zhu, W.F.; Kramer, J.S.; Proschak, E. Computer-Aided fragment growing strategies to design dual inhibitors of soluble epoxide hydrolase and LTA4 hydrolase. ACS Med. Chem. Lett. 2020, 11, 1244–1249. [Google Scholar] [CrossRef]
- Moser, D.; Wisniewska, J.M.; Hahn, S.; Achenback, J.; Buscato, E.; Klingler, F.-M.; Hofmann, B.; Steinhilber, D.; Proschak, E. Dual-Target virtual screening by pharmacophore elucidation and molecular shape filtering. ACS Med. Chem. Lett. 2012, 3, 155–158. [Google Scholar] [CrossRef]
- Nandha, B.; Ramareddy, S.A.; Hazra, K. Synthesis of substituted fluorobenzimidazoles as inhibitors of 5-lipoxygenase and soluble epoxide hydrolase for anti-inflammatory activity. Arch. Pharm. 2018, 351, 1800030. [Google Scholar] [CrossRef]
- Meirer, K.; Rodl, C.B.; Wisniewska, J.M.; George, S.; Hafner, A.-K.; Buscato, E.; Klingler, F.-M.; Hahn, S.; Berressem, D.; Wittmann, S.K.; et al. Synthesis and structure-activity relationship studies of novel dual inhibitors of soluble epoxide hydrolase and 5-lipoxygenase. J. Med. Chem. 2013, 56, 1777–1781. [Google Scholar] [CrossRef] [PubMed]
- Achenback, J.; Klingler, F.-M.; Blocher, R.; Moser, D.; Hafner, A.K.; Rodl, C.B.; Kretschmer, S.; Kruger, B.; Lohr, F.; Stark, H.; et al. Exploring the chemical space of multitarget ligands using self-organizing maps. ACS Med. Chem. Lett. 2013, 4, 1169–1172. [Google Scholar] [CrossRef] [PubMed]
- Meirer, K.; Glatzel, D.; Kretschmer, S.; Wittmann, S.K.; Hartmann, M.; Blocher, R.; Angioni, C.; Geisslinger, G.; Steinhilber, D.; Hofmann, B.; et al. Design, synthesis and cellular characterization of a dual inhibitor of 5-lipoxygenase and soluble epoxide hydrolase. Molecules 2017, 22, 45. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.D.; Bhakta, S.; Hwang, S.H.; Pakhomova, S.; Newcomer, M.E.; Morisseau, C.; Hammock, B.D. Identification and optimization of soluble epoxide hydrolase inhibitors with dual potency towards fatty acid amide hydrolase. Bioorg. Med. Chem. Lett. 2018, 28, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.D.; Wan, D.; Wagner, K.M.; Hwang, S.H.; Morisseau, C.; Hammock, B.D. Design and potency of dual soluble epoxide hydrolase/fatty acid amide hydrolase inhibitors. ACS Omega 2018, 3, 14076–14086. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Zhang, B.; Xu, M.; Morisseau, C.; Hwang, S.H.; Hammock, B.D.; Li, Q.X. 1-Trifluoromethoxyphenyl-2-(1-propionylpiperidin-4-yl) urea, a selective and potent dual inhibitor of soluble epoxide hydrolase and p38 kinase intervenes in Alzheimer’s signaling in human nerve cells. ACS Chem. Neurosci. 2019, 10, 4018–4030. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Rotter, M.; Weiser, T.; Wittmann, S.; Weizel, L.; Kaiser, A.; Heering, J.; Goebel, T.; Angioni, C.; Wurglics, M.; et al. A dual modulator of farnesoid X receptor and soluble epoxide hydrolase to counter nonalcoholic steatohepatitis. J. Med. Chem. 2017, 60, 7703–7724. [Google Scholar] [CrossRef] [PubMed]
- Schierle, S.; Helmstadter, M.; Schmidt, J.; Hartmann, M.; Horz, M.; Kaiser, A.; Weizel, L.; Heitel, P.; Proschak, A.; Hernandez-Olmos, V.; et al. Dual farnesoid X receptor/soluble epoxide hydrolase modulators derived from Zafirlukast. ChemMedChem 2020, 15, 50–67. [Google Scholar] [CrossRef]
- Inceoglu, B.; Wagner, K.; Schebb, N.H.; Morisseau, C.; Jinks, S.L.; Ulu, A.; Hegedus, C.; Rose, T.; Brosnan, R.; Hammock, B.D. Analgesia mediated by soluble epoxide hydrolase inhibitors is dependent on cAMP. Proc. Natl. Acad. Sci. USA 2011, 108, 5093–5097. [Google Scholar] [CrossRef]
- Blocher, R.; Wagner, K.M.; Gopireddy, R.R.; Harris, T.R.; Wu, H.; Barnych, B.; Hwang, S.H.; Xiang, Y.K.; Proschak, E.; Morisseau, C.; et al. Orally available soluble epoxide hydrolase/phosphodiesterase 4 dual inhibitor treats inflammatory pain. J. Med. Chem. 2018, 26, 3541–3550. [Google Scholar] [CrossRef]
- Roca, H.; Jones, J.D.; Purica, M.C.; Weidner, S.; Koh, A.J.; Kuo, R.; Wilkinson, J.E.; Wang, Y.; Daignault-Newton, S.; Pienta, K.J.; et al. Apoptosis-induced CXCL5 accelerates inflammation and growth of prostate tumor metastases in bone. J. Clin. Investig. 2018, 128, 248–266. [Google Scholar] [CrossRef] [PubMed]
- Stanford, J.C.; Young, C.; Hicks, D.; Owens, P.; Williams, A.; Vaught, D.B.; Morrison, M.M.; Lim, J.; Williams, M.; Brantley-Sieders, D.M.; et al. Efferocytosis produces a prometastatic landscape during postpartum mammary gland involution. J. Clin. Investig. 2014, 124, 4737–4752. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Liao, J.; Berry, J.E.; Li, X.; Koh, A.J.; Michalski, M.E.; Eber, M.R.; Soki, F.N.; Sadler, D.; Sud, S.; et al. Cyclophosphamide creates a receptive microenvironment for prostate cancer skeletal metastasis. Cancer Res. 2012, 72, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Gartung, A.; Yang, J.; Sukhatme, V.P.; Bielenberg, D.R.; Fernandes, D.; Chang, J.; Schmidt, B.A.; Hwang, S.H.; Zurakowski, D.; Huang, S.; et al. Suppression of chemotherapy-induced cytokine/lipid mediator surge and ovarian cancer by a dual COX-2/sEH inhibitor. Proc. Natl. Acad. Sci. USA 2019, 116, 1698–1703. [Google Scholar] [CrossRef]
- Ho, G.Y.; Woodward, N.; Coward, J.I. Cisplatin versus carboplatin: Comparative review of therapeutic management in solid malignancies. Crit. Rev. Oncol. Hematol. 2016, 102, 37–46. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, H.; Ma, A.H.; Yu, W.; Zimmermann, M.; Yang, J.; Hwang, S.H.; Zhu, D.; Lin, T.Y.; Malfatti, M.; et al. COX-2/sEH dual inhibitor PTUPB potentiates the antitumor efficacy of cisplatin. Mol. Cancer Ther. 2018, 17, 474–483. [Google Scholar] [CrossRef]
- Ghosh, N.; Chaki, R.; Mandal, V.; Mandal, S.C. COX-2 as a target for cancer chemotherapy. Pharmacol. Rep. 2010, 62, 233–244. [Google Scholar] [CrossRef]
- Gridelli, C.; Gallo, C.; Ceribelli, A.; Gebbia, V.; Gamucci, T.; Ciardiello, F.; Carozza, F.; Favaretto, A.; Daniele, B.; Galetta, D.; et al. Factorial phase III randomised trial of rofecoxib and prolonged constant infusion of gemcitabine in advanced non-small-cell lung cancer: The GEmcitabine-COxib in NSCLC (GECO) study. Lancet Oncol. 2007, 8, 500–512. [Google Scholar] [CrossRef]
- Pan, C.X.; Loehrer, P.; Seitz, D.; Helft, P.; Juliar, B.; Ansari, R.; Pletcher, W.; Vinson, J.; Cheng, L.; Sweeney, C. A phase II trial of irinotecan, 5-fluorouracil and leucovorin combined with celecoxib and glutamine as first-line therapy for advanced colorectal cancer. Oncology 2005, 69, 63–70. [Google Scholar] [CrossRef]
- Lowenfels, A.B.; Maisonneuve, P.; Cavallini, G.; Ammann, R.W.; Lankisch, P.G.; Andersen, J.R.; Dimagno, E.P.; Andren-Sandberg, A.; Domellof, L.; Group, I.P.S. Pancreatitis and the risk of pancreatic cancer. N. Engl. J. Med. 1993, 328, 1433–1437. [Google Scholar] [CrossRef]
- Hingorani, S.R.; Wang, L.; Multani, A.S.; Combs, C.; Deramaudt, T.B.; Hruban, R.H.; Rustgi, A.K.; Chang, S.; Tuveson, D.A. Trp53R172H and KrasG12D cooperate to promote chromosomal instability and widely metastatic pancreatic ductal adenocarcinoma in mice. Cancer Cell 2005, 7, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Pylayeva-Gupta, Y.; Grabocka, E.; Bar-Sagi, D. RAS oncogenes: Weaving a tumorigenic web. Nat. Rev. Cancer 2011, 11, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Blasco, R.B.; Francoz, S.; Santamaría, D.; Cañamero, M.; Dubus, P.; Charron, J.; Baccarini, M.; Barbacid, M. C-Raf, but not B-Raf, is essential for development of K-Ras oncogene-driven non-small cell lung carcinoma. Cancer Cell 2011, 19, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Hwang, S.H.; Li, H.; Liu, J.Y.; Hammock, B.D.; Yang, G.Y. Inhibition of chronic pancreatitis and murine pancreatic intraepithelial neoplasia by a dual inhibitor of c-RAF and soluble epoxide hydrolase in LSL-KrasG12D/Pdx-1-Cre mice. Anticancer Res. 2016, 36, 27–37. [Google Scholar]
- Zhang, G.; Panigrahy, D.; Mahakian, L.M.; Yang, J.; Liu, J.Y.; Lee, K.S.S.; Wettersten, H.I.; Ulu, A.; Hu, X.; Tam, S.; et al. Epoxy metabolites of docosahexaenoic acid (DHA) inhibit angiogenesis, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 6530–6535. [Google Scholar] [CrossRef]
- Morin, C.; Sirois, M.; Échavé, V.; Albadine, R.; Rousseau, E. 17, 18-Epoxyeicosatetraenoic acid targets PPARγ and p38 mitogen-activated protein kinase to mediate its anti-inflammatory effects in the lung: Role of soluble epoxide hydrolase. Am. J. Respir. Cell Mol. Biol. 2010, 43, 564–575. [Google Scholar] [CrossRef]
- Kubota, T.; Arita, M.; Isobe, Y.; Iwamoto, R.; Goto, T.; Yoshioka, T.; Urabe, D.; Inoue, M.; Arai, H. Eicosapentaenoic acid is converted via ω-3 epoxygenation to the anti-inflammatory metabolite 12-hydroxy-17, 18-epoxyeicosatetraenoic acid. FASEB J. 2014, 28, 586–593. [Google Scholar] [CrossRef]
- Xia, R.; Sun, L.; Liao, J.; Li, H.; You, X.; Xu, D.; Yang, J.; Hwang, S.H.; Jones, R.D.; Hammock, B.; et al. Inhibition of pancreatic carcinoma growth through enhancing ω-3 Epoxy polyunsaturated fatty acid profile by inhibition of soluble epoxide hydrolase. Anticancer Res. 2019, 39, 3651–3660. [Google Scholar] [CrossRef]
- Freitas, R.D.; Campos, M.M. Protective effects of omega-3 fatty acids in cancer-related complications. Nutrients 2019, 11, 945. [Google Scholar] [CrossRef]
- Dong, X.W.; Jia, Y.L.; Jiang, B.; Jiang, J.X.; Shen, J.; Jin, Y.C.; Guan, Y.; Sun, Y.; Xie, Q.M. Soluble epoxide hydrolase inhibitor AUDA decreases bleomycin-induced pulmonary toxicity in mice by inhibiting the p38/Smad3 pathways. Toxicology 2017, 389, 31–41. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Xing, B.; Yu, Y.; Wang, H.; Chen, G.; Gu, B.; Zhang, G.; Wei, D.; Gu, P.; et al. t-AUCB, an improved sEH inhibitor, suppresses human glioblastoma cell growth by activating NF-κB-p65. J. Neuro Oncol. 2012, 108, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, J.; Zhang, J.; Wang, Y.; Hwang, S.H.; Qi, W.; Wan, D.; Kim, D.; Sun, J.; Sanidad, K.Z.; et al. Lipidomic profiling reveals soluble epoxide hydrolase as a therapeutic target of obesity-induced colonic inflammation. Proc. Natl. Acad. Sci. USA 2018, 115, 5283–5288. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Wecksler, A.T.; Zhang, G.; Morisseau, C.; Nguyen, L.V.; Fu, S.H.; Hammock, B.D. Synthesis and biological evaluation of sorafenib-and regorafenib-like sEH inhibitors. Bioorg. Med. Chem. Lett. 2013, 23, 3732–3737. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das Mahapatra, A.; Choubey, R.; Datta, B. Small Molecule Soluble Epoxide Hydrolase Inhibitors in Multitarget and Combination Therapies for Inflammation and Cancer. Molecules 2020, 25, 5488. https://doi.org/10.3390/molecules25235488
Das Mahapatra A, Choubey R, Datta B. Small Molecule Soluble Epoxide Hydrolase Inhibitors in Multitarget and Combination Therapies for Inflammation and Cancer. Molecules. 2020; 25(23):5488. https://doi.org/10.3390/molecules25235488
Chicago/Turabian StyleDas Mahapatra, Amarjyoti, Rinku Choubey, and Bhaskar Datta. 2020. "Small Molecule Soluble Epoxide Hydrolase Inhibitors in Multitarget and Combination Therapies for Inflammation and Cancer" Molecules 25, no. 23: 5488. https://doi.org/10.3390/molecules25235488
APA StyleDas Mahapatra, A., Choubey, R., & Datta, B. (2020). Small Molecule Soluble Epoxide Hydrolase Inhibitors in Multitarget and Combination Therapies for Inflammation and Cancer. Molecules, 25(23), 5488. https://doi.org/10.3390/molecules25235488