The Effect of Protein-Rich Extract from Bombyx Batryticatus against Glutamate-Damaged PC12 Cells Via Regulating γ-Aminobutyric Acid Signaling Pathway


Abstract
:1. Introduction
2. Results
2.1. Optimization of Extraction Conditions
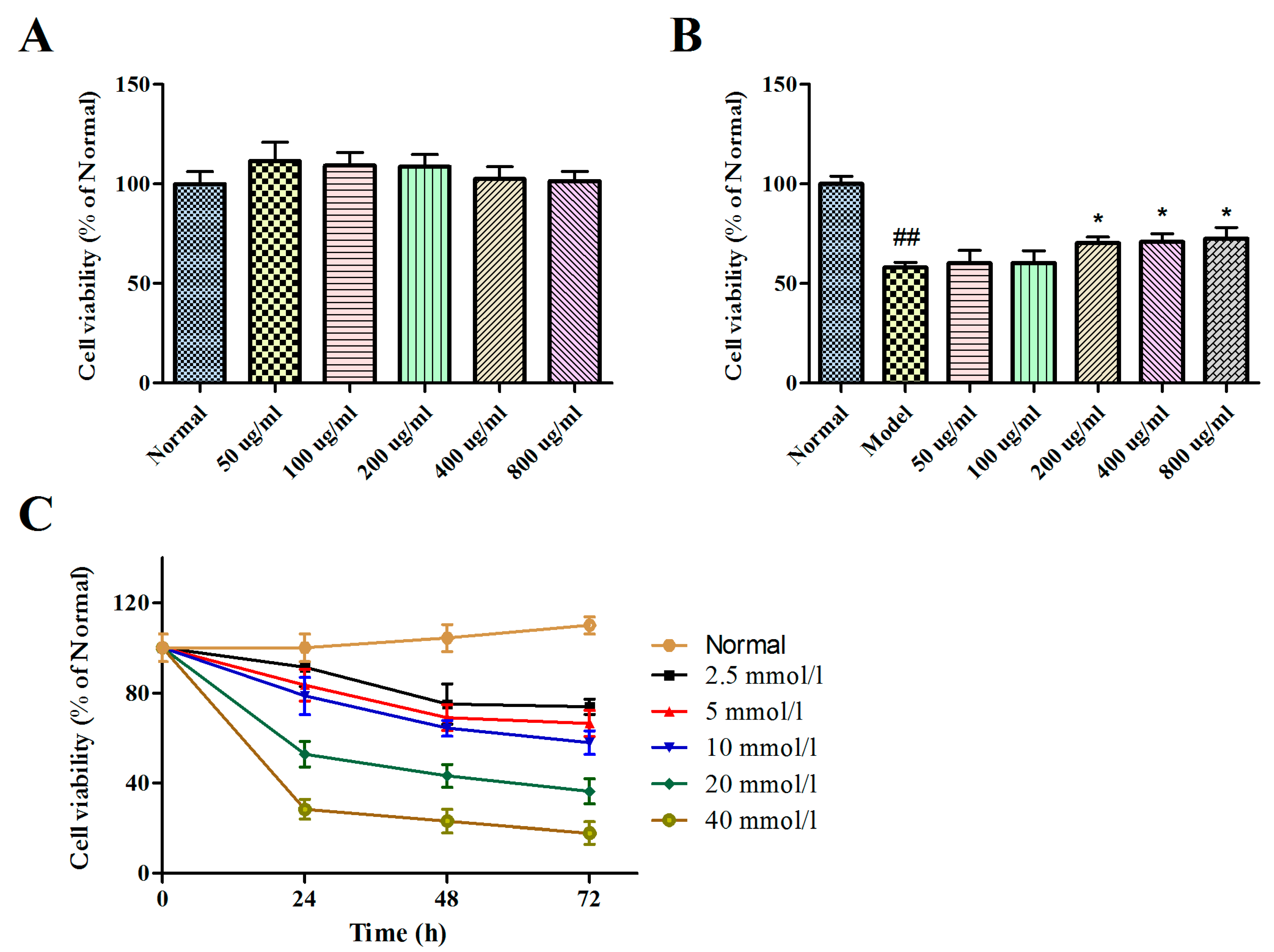
2.2. Effects of BBPs and Glu on Cell Viability of PC12 Cells
2.3. Effects of BBPs on GABA, IL-1β, IL-4, 5-HT, TNF-α and GRα in NGF-Induced PC12 Cells Injured by Glu
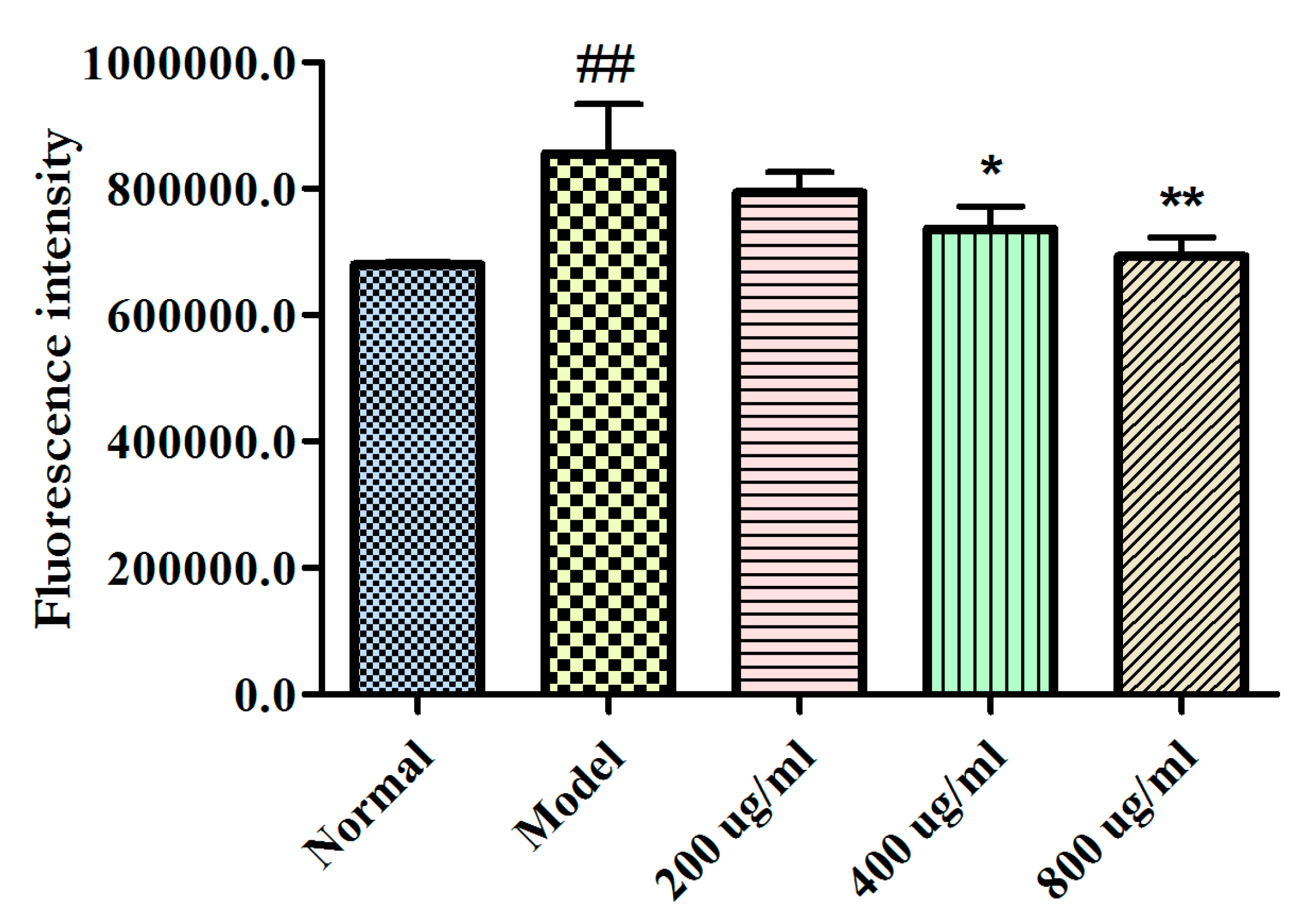
2.4. Effects of BBPs on Ca2+ Levels of NGF-Induced PC12 Cells Injured by Glu
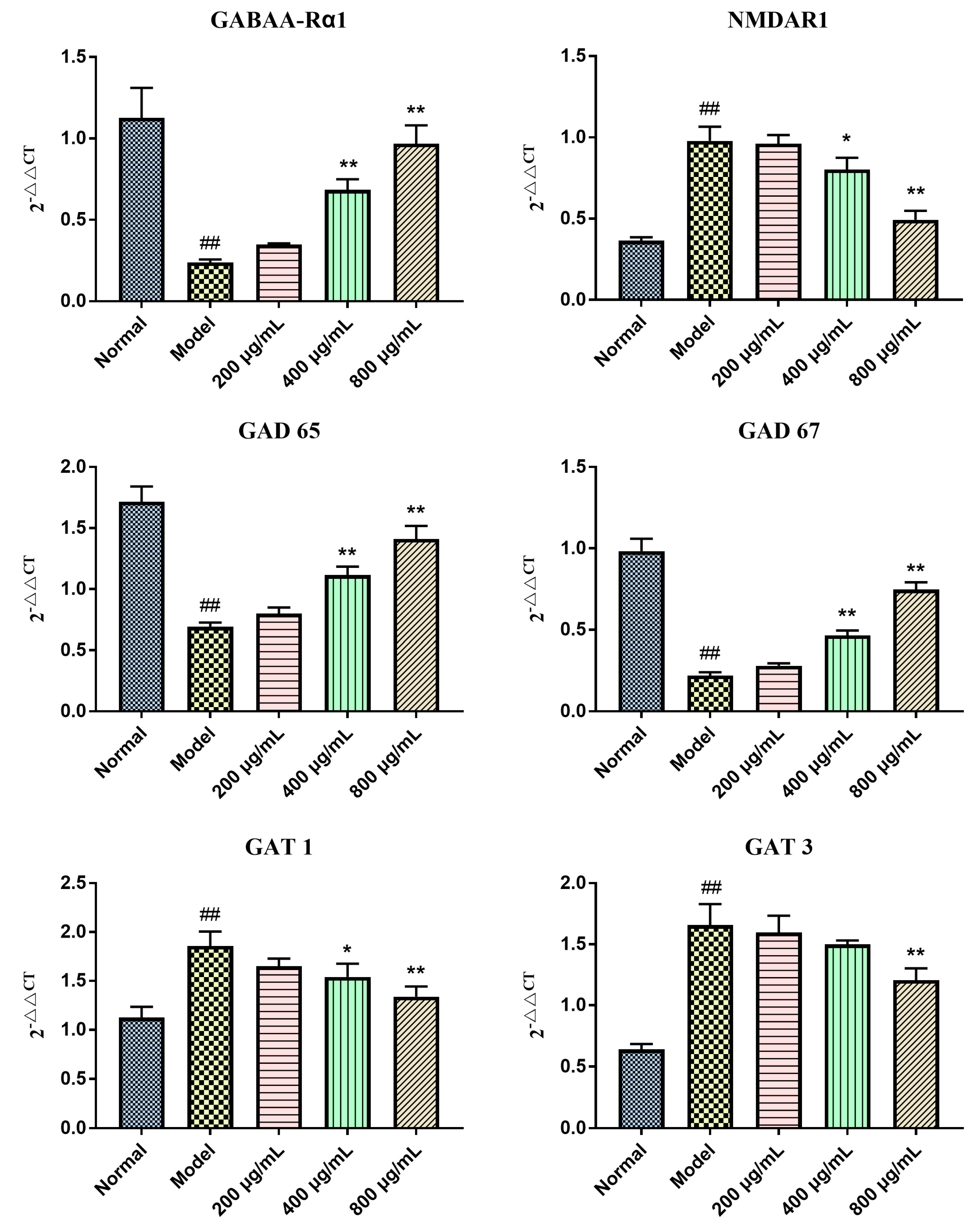
2.5. Effects of BBPs on mRNA Expressions of GABAA-Rα1, NMDAR1, GAD 65, GAT 1, GAT 3 and GAD 67 in NGF-Induced PC12 Cells Injured by Glu
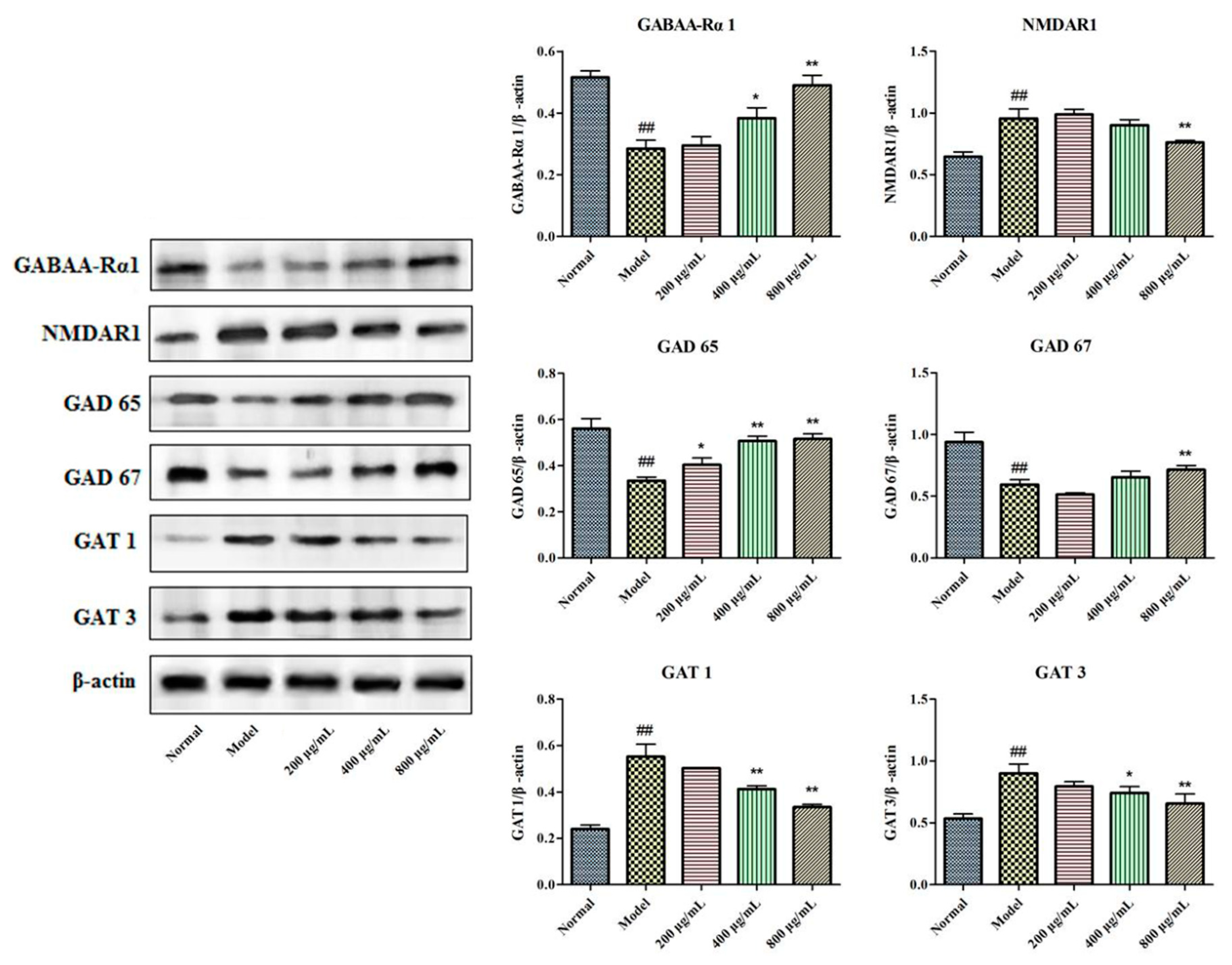
2.6. Effects of BBPs on Protein Expressions of GABAA-Rα1, NMDAR1, GAD 65, GAT 1, GAT 3 and GAD 67 in NGF-Induced PC12 Cells Injured by Glu
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Extraction of BBPs by Ultrasonic-Assisted Extraction (UAE)
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Determination of GABA, IL-1β, IL-4, TNF-α and GRα Content in PC12 Cells
4.6. Measurement of Cytosolic Free Calcium (Ca2+) Release
4.7. Quantitative Real-Time Polymerase Chain Reaction (RT-PCR) Assay
4.8. Western Blot Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gooneratne, I.K.; Green, A.L.; Dugan, P.; Sen, A.; Franzini, A.; Aziz, T.; Cheeran, B. Comparing neurostimulation technologies in refractory focal-onset epilepsy. J. Neurol. Neurosurg. Psychiatry 2016, 87, 1174–1182. [Google Scholar] [CrossRef]
- Weber, Y.G.; Biskup, S.; Helbig, K.L.; Von Spiczak, S.; Lerche, H. The role of genetic testing in epilepsy diagnosis and management. Expert Rev. Mol. Diagn. 2017, 17, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Mula, M. Investigational new drugs for focal epilepsy. Expert Opin. Investig. Drugs 2015, 25, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manford, M. Recent advances in epilepsy. J. Neurol. 2017, 264, 1811–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Yongyan, W.U.; Song, D.; Yan, Z.; Liu, T. Compounds isolated and purified from chloroform active part of bombyx batryticatus and their anticonvulsive activities. Chin. J. Pharm. 2014, 45, 431–433. [Google Scholar]
- Karpova, M.N.; Kuznetzova, L.V.; Klishina, N.Y. Gaba and its Receptors in Pathogenesis of Epilepsy. Uspekhi fiziologicheskikh nauk 2015, 46, 46–59. [Google Scholar]
- Qiu, W.J.; Hu, X.W. Research progress on pathogenesis and treatment of epilepsy. Chin. J. Clin. Phys. 2014, 8, 101–105. [Google Scholar]
- Obata, K. Synaptic inhibition and γ-aminobutyric acid in the mammalian central nervous system. Proc. Jpn. Acad. 2013, 89, 139–156. [Google Scholar] [CrossRef] [Green Version]
- Pinal, C.S.; Tobin, A.J. Uniqueness and redundancy in GABA production. Perspect. Dev. Neurobiol. 1998, 5, 109–118. [Google Scholar]
- Soghomonian, J.-J.; Martin, D.L. Two isoforms of glutamate decarboxylase: Why? Trends Pharmacol. Sci. 1998, 19, 500–505. [Google Scholar] [CrossRef]
- Stork, O.; Ji, F.-Y.; Kaneko, K.; Stork, S.; Yoshinobu, Y.; Moriya, T.; Shibata, S.; Obata, K. Postnatal development of a GABA deficit and disturbance of neural functions in mice lacking GAD65. Brain Res. 2000, 865, 45–58. [Google Scholar] [CrossRef]
- Dalby, N.O. Inhibition of γ-aminobutyric acid uptake: Anatomy, physiology and effects against epileptic seizures. Eur. J. Pharmacol. 2003, 479, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Jing, G.; Zhu, M.D. Advances in the pathogenesis of epilepsy. J. Shenyang Med. Coll. 2012, 14, 111–113. [Google Scholar]
- Ravizza, T.; Vezzani, A. Status epilepticus induces time-dependent neuronal and astrocytic expression of interleukin-1 receptor type I in the rat limbic system. Neuroscience 2006, 137, 301–308. [Google Scholar] [CrossRef]
- Dey, A.; Kang, X.; Qiu, J.; Du, Y.; Jiang, J. Anti-Inflammatory Small Molecules to Treat Seizures and Epilepsy: From Bench to Bedside. Trends Pharmacol. Sci. 2016, 37, 463–484. [Google Scholar] [CrossRef] [Green Version]
- Sonar, S.; Lal, G. Role of Tumor Necrosis Factor Superfamily in Neuroinflammation and Autoimmunity. Front. Immunol. 2015, 6, 364. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Li, X.; Liu, N.; Zheng, P.; Ma, L.; Guo, F.; Sun, T.; Zhou, R.; Yu, J. The Anticonvulsant Effects of Baldrinal on Pilocarpine-Induced convulsion in Adult Male Mice. Molecules 2019, 24, 1617. [Google Scholar] [CrossRef] [Green Version]
- Shandra, A.A.; Godlevsky, L.S.; Vastyanov, R.S.; Oleinik, A.A.; Konovalenko, V.L.; Rapoport, E.N.; Korobka, N.N. The role of TNF-alpha in amygdala kindled rats. Neurosci. Res. 2002, 42, 147–153. [Google Scholar] [CrossRef]
- Li, T.; Zhai, X.; Jiang, J.; Song, X.; Han, W.; Ma, J.; Xie, L.; Cheng, L.; Chen, H.; Jiang, L. Intraperitoneal injection of IL-4/IFN-γ modulates the proportions of microglial phenotypes and improves epilepsy outcomes in a pilocarpine model of acquired epilepsy. Brain Res. 2017, 1657, 120–129. [Google Scholar] [CrossRef]
- Pijnenburg-Kleizen, K.J.; Engels, M.; Mooij, C.F.; Griffin, A.; Krone, N.; Span, P.N.; Van Herwaarden, A.E.; Sweep, F.C.G.J.; Der Grinten, H.L.C.-V. Adrenal Steroid Metabolites Accumulating in Congenital Adrenal Hyperplasia Lead to Transactivation of the Glucocorticoid Receptor. Endocrinology 2015, 156, 3504–3510. [Google Scholar] [CrossRef]
- Yu, C.-W.; Chang, P.-T.; Hsin, L.-W.; Chern, J.-W. Quinazolin-4-one Derivatives as Selective Histone Deacetylase-6 Inhibitors for the Treatment of Alzheimer’s Disease. J. Med. Chem. 2013, 56, 6775–6791. [Google Scholar] [CrossRef] [PubMed]
- Nishina, A.; Kimura, H.; Tsukagoshi, H.; Kozawa, K.; Koketsu, M.; Ninomiya, M.; Sato, D.; Obara, Y.; Furukawa, S. Neurite outgrowth of PC12 cells by 4’-O-β-D-glucopyranosyl-3’,4-dimethoxychalcone from Brassica rapa L. ‘hidabeni’ was enhanced by pretreatment with p38MAPK inhibitor. Neurochem. Res. 2013, 38, 2397–2407. [Google Scholar] [CrossRef] [PubMed]
- Terada, K.; Matsushima, Y.; Matsunaga, K.; Takata, J.; Karube, Y.; Ishige, A. The Kampo medicine Yokukansan (YKS) enhances nerve growth factor (NGF)-induced neurite outgrowth in PC12 cells. Bosn. J. Basic Med. Sci. 2018, 18, 224–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Yan, N.; Huang, Y.; Cao, G.; Deng, J.; Deng, Y. Effects of nerve growth factor on nerve regeneration after corneal nerve damage. Int. J. Clin. Exp. Med. 2014, 7, 4584–4589. [Google Scholar]
- Zeng, X.; Hu, K.; Chen, L.; Zhou, L.; Luo, W.; Li, C.; Zong, W.; Chen, S.; Gao, Q.; Zeng, G.; et al. The effects of ginsenoside compound k against epilepsy by enhancing the gamma-aminobutyric acid signaling pathway. Front. Pharmacol. 2018, 9, 1020. [Google Scholar] [CrossRef]
- Kim, M.H.; Lee, H.J.; Lee, S.-R.; Lee, H.-S.; Huh, J.-W.; Bae, Y.C.; Lee, D.-S. Peroxiredoxin 5 Inhibits Glutamate-Induced Neuronal Cell Death through the Regulation of Calcineurin-Dependent Mitochondrial Dynamics in HT22 Cells. Mol. Cell. Biol. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Soukupová, M.; Binaschi, A.; Falcicchia, C.; Palma, E.; Roncon, P.; Zucchini, S.; Simonato, M. Increased extracellular levels of glutamate in the hippocampus of chronically epileptic rats. Neuroscience 2015, 301, 246–253. [Google Scholar] [CrossRef]
- Albrecht, J.; Zielińska, M. Mechanisms of Excessive Extracellular Glutamate Accumulation in Temporal Lobe Epilepsy. Neurochem. Res. 2017, 42, 1724–1734. [Google Scholar] [CrossRef]
- Yu, S.P.; Strasser, U.; Tian, M.; Choi, D.W.J.S. Nmda receptor-mediated k+ efflux and neuronal apoptosis. Science 1999, 284, 336–339. [Google Scholar] [CrossRef]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The Neuroprotective Properties of Hericium erinaceus in Glutamate-Damaged Differentiated PC12 Cells and an Alzheimer’s Disease Mouse Model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef]
- Huang, H.; Peng, X.; Peng, Y. Modern research progress of bombyx mori. J. Hunan. Univ. Tradit. Chin. Med. 2003, 23, 62–64. [Google Scholar]
- Wu, J.-Y.; Sheikho, A.; Ma, H.; Li, T.-C.; Zhao, Y.-Q.; Zhang, Y.-L.; Wang, D. Molecular mechanisms of Bombyx batryticatus ethanol extract inducing gastric cancer SGC-7901 cells apoptosis. Cytotechnology 2017, 69, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Jia, T.Z.; Cao, Q.C.; Tian, F.; Ying, W.T. A Crude 1-DNJ Extract from Home Made Bombyx Batryticatus Inhibits Diabetic Cardiomyopathy-Associated Fibrosis in db/db Mice and Reduces Protein N-Glycosylation Levels. Int. J. Mol. Sci. 2018, 19, 1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Wang, G.J.; Wang, J.; Chu, Y.L. Advances in studies on constituents and pharmacological actions of bombyx mori. China. Seri. Cul. 2004, 4, 87–89. [Google Scholar]
- Hu, M.; Yu, Z.; Wang, J.; Fan, W.; Liu, Y.; Li, J.; Xiao, H.; Li, Y.; Peng, W.; Wu, C. Traditional Uses, Origins, Chemistry and Pharmacology of Bombyx batryticatus: A Review. Molecules 2017, 22, 1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Liu, Y.; He, L.; Yuan, X.; Peng, W.; Wu, C. Antiepileptic Effects of Protein-Rich Extract from Bombyx batryticatus on Mice and Its Protective Effects against H2O2-Induced Oxidative Damage in PC12 Cells via Regulating PI3K/Akt Signaling Pathways. Oxid. Med. Cell. Longev. 2019, 2019, 7897584. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.-S.; An, H.-G.; Moon, S.-K.; Lee, Y.-C.; Kim, H.-M.; Ko, J.-H.; Kim, C.-H. Bombycis corpus extract (BCE) protects hippocampal neurons against excitatory amino acid-induced neurotoxicity. Immunopharmacol. Immunotoxicol. 2003, 25, 191–201. [Google Scholar] [CrossRef]
- Bai, Y.; Zhao, Q.; He, M.; Ye, X.; Zhang, X. Extensive characterization and differential analysis of endogenous peptides from Bombyx batryticatus using mass spectrometric approach. J. Pharm. Biomed. Anal. 2019, 163, 78–87. [Google Scholar] [CrossRef]
- Cheng, S.-M.; Huang, J.; Wang, H.-Y.; Li, G.-Y.; Lin, R.-C.; Wang, J.-H. Two new compounds from Bombyx batryticatus. J. Asian Nat. Prod. Res. 2014, 16, 825–829. [Google Scholar] [CrossRef]
- Shindyapina, A.V.; Komarova, T.V.; Sheshukova, E.V.; Ershova, N.M.; Tashlitsky, V.N.; Kurkin, A.V.; Yusupov, I.R.; Mkrtchyan, G.V.; Shagidulin, M.Y.; Dorokhov, Y.L. The Antioxidant Cofactor Alpha-Lipoic Acid May Control Endogenous Formaldehyde Metabolism in Mammals. Front. Neurosci. 2017, 11, 651. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.G.; Martinez-Becerra, F.; Antonio, L.; Johnson, R.T.; De Campos, R.P.S.; Siegel, J.M.; Wijesinghe, M.B.; Lazzarino, G.; Lunte, S.M. Carnosine modulates nitric oxide in stimulated murine RAW 264.7 macrophages. Mol. Cell. Biochem. 2017, 431, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Magarkar, A.; Jurkiewicz, P.; Allolio, C.; Hof, M.; Jungwirth, P. Increased Binding of Calcium Ions at Positively Curved Phospholipid Membranes. J. Phys. Chem. Lett. 2017, 8, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Krisanova, N.; Pozdnyakova, N.; Pastukhov, A.; Dudarenko, M.; Maksymchuk, O.; Parkhomets, P.; Sivko, R.; Borisova, T. Vitamin D3 deficiency in puberty rats causes presynaptic malfunctioning through alterations in exocytotic release and uptake of glutamate/GABA and expression of EAAC-1/GAT-3 transporters. Food Chem. Toxicol. 2019, 123, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, C.; Nistér, M. Protein extraction from solid tissue. Methods. Mol. Biol. 2011, 675, 307–312. [Google Scholar]
- Chen, Y.; Li, C.; Zhu, J.; Xie, W.; Hu, X.; Song, L.; Zi, J.; Yu, R. Purification and characterization of an antibacterial and anti-inflammatory polypeptide from Arca subcrenata. Int. J. Biol. Macromol. 2017, 96, 177–184. [Google Scholar] [CrossRef]
- Lin, K.H.; Li, C.Y.; Hsu, Y.M.; Tsai, F.J.; Tang, C.H.; Yang, J.-S.; Wang, Z.H.; Yin, M.C. Oridonin, A natural diterpenoid, protected NGF-differentiated PC12 cells against MPP+- and kainic acid-induced injury. Food Chem. Toxicol. 2019, 133, 110765. [Google Scholar] [CrossRef]
- Overbeeke, R.; Yildirim, M.; Reutelingsperger, C.P.; Haanen, C.; Vermes, I. Sequential occurrence of mitochondrial and plasma membrane alterations, fluctuations in cellular Ca2+ and pH during initial and later phases of cell death. Apoptosis 1999, 4, 455–460. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Sum of Squares | df | Mean Square | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Model | 2.59 | 9 | 0.29 | 151.46 | <0.0001 |
| A-A | 0.039 | 1 | 0.039 | 20.59 | 0.0027 |
| B-B | 0.070 | 1 | 0.070 | 36.94 | 0.0005 |
| C-C | 0.58 | 1 | 0.58 | 303.54 | <0.0001 |
| AB | 0.14 | 1 | 0.14 | 75.86 | <0.0001 |
| AC | 4 × 10−4 | 1 | 4 × 10−4 | 0.21 | 0.6606 |
| BC | 4.225 × 10−3 | 1 | 4.225 × 10−3 | 2.22 | 0.1799 |
| A2 | 0.64 | 1 | 0.64 | 334.28 | <0.0001 |
| B2 | 0.35 | 1 | 0.35 | 181.24 | <0.0001 |
| C2 | 0.60 | 1 | 0.60 | 313.13 | <0.0001 |
| Residual | 0.013 | 7 | 1.904 × 10−3 | ||
| Lack of Fit | 6.325 × 10−3 | 3 | 2.108 × 10−3 | 1.20 | 0.4153 |
| Pure Error | 7 × 10−3 | 4 | 1.750 × 10−3 | -- | -- |
| Cor Total | 2.61 | 16 | -- | -- | |
| Standard deviation | 0.044 | R2 | 0.9949 | -- | -- |
| C.V.% | 2.19 | Adj R2 | 0.9883 | -- | -- |
| Adeq Precision | 32.686 | Pred R2 | 0.9570 | -- | -- |
| Run | Extracting Time (A) (h) | Material Liquid Than (B) (mL/g) | Ultrasonic Power (C) (W) | Extraction Yield (%) |
|---|---|---|---|---|
| 1 | 0.75 | 3.00 | 220 | 1.64 |
| 2 | 1.25 | 3.00 | 220 | 2.18 |
| 3 | 0.75 | 5.00 | 220 | 1.83 |
| 4 | 1.25 | 5.00 | 220 | 1.61 |
| 5 | 0.75 | 4.00 | 180 | 1.38 |
| 6 | 1.25 | 4.00 | 180 | 1.48 |
| 7 | 0.75 | 4.00 | 260 | 1.95 |
| 8 | 1.25 | 4.00 | 260 | 2.09 |
| 9 | 1.00 | 3.00 | 180 | 1.71 |
| 10 | 1.00 | 5.00 | 180 | 1.46 |
| 11 | 1.00 | 3.00 | 260 | 2.13 |
| 12 | 1.00 | 5.00 | 260 | 2.01 |
| 13 | 1.00 | 4.00 | 220 | 2.48 |
| 14 | 1.00 | 4.00 | 220 | 2.44 |
| 15 | 1.00 | 4.00 | 220 | 2.51 |
| 16 | 1.00 | 4.00 | 220 | 2.47 |
| 17 | 1.00 | 4.00 | 220 | 2.55 |
| Gene Name | Forward Primer | Reverse Primer | Product Length |
|---|---|---|---|
| β-actin | GAAGATCAAGATCATTGCTCC | TACTCCTGCTTGCTGATCCA | 111 bp |
| GABAA-Rα1 | CGCTCAGTGGTTGTGGCAGAAGATGG | GTCACGGTCAGAACGGTCGTCACTCC | 272 bp |
| GAD 67 | GTGCTGCTCCAGTGTTCTGCCATCC | AATCCCACAGTGCCCTTTGCTTTCCA | 203 bp |
| GAD 65 | CAAGTGGAAGCTGAACGGTGTGGAGA | TCTGACCAGGAGAGCCGAACATTGC | 100 bp |
| NMDAR1 | GGCACACAGGAGCGGGTAAACAACAG | AAGCGGTCCAGCAGGTACAGCATCA | 296 bp |
| GAT 1 | TTGGCTGGCGGGCGTGTTTCTCTTCA | TGCGGCTGCTCAGGACCATTCTCA | 246 bp |
| GAT 3 | GGTGCTGGCTCATGGCTCTGTCCT | AAGTGCGTCTCCTTCTCTGTGATGGC | 238 bp |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.-Y.; Hu, M.-B.; Li, R.-L.; Zhao, R.; Fan, L.-H.; Wang, L.; Peng, W.; Liu, Y.-J.; Wu, C.-J. The Effect of Protein-Rich Extract from Bombyx Batryticatus against Glutamate-Damaged PC12 Cells Via Regulating γ-Aminobutyric Acid Signaling Pathway. Molecules 2020, 25, 553. https://doi.org/10.3390/molecules25030553
He L-Y, Hu M-B, Li R-L, Zhao R, Fan L-H, Wang L, Peng W, Liu Y-J, Wu C-J. The Effect of Protein-Rich Extract from Bombyx Batryticatus against Glutamate-Damaged PC12 Cells Via Regulating γ-Aminobutyric Acid Signaling Pathway. Molecules. 2020; 25(3):553. https://doi.org/10.3390/molecules25030553
Chicago/Turabian StyleHe, Li-Ying, Mei-Bian Hu, Ruo-Lan Li, Rong Zhao, Lin-Hong Fan, Li Wang, Wei Peng, Yu-Jie Liu, and Chun-Jie Wu. 2020. "The Effect of Protein-Rich Extract from Bombyx Batryticatus against Glutamate-Damaged PC12 Cells Via Regulating γ-Aminobutyric Acid Signaling Pathway" Molecules 25, no. 3: 553. https://doi.org/10.3390/molecules25030553