
The Microbiome and Its Implications in Cancer Immunotherapy

Abstract
:1. Introduction
2. The Microbiome and Its Association with Various Kinds of Cancer
2.1. Colon Cancer
2.2. Breast Cancer
2.3. Oral Cancer
2.4. Liver Cancer
3. Microbiome Impact on Immunotherapy
3.1. Microbiome Implications in CTLA-4 Based Immunotherapy
3.2. Microbiome Implications in PD-1/PD-L1 Inhibitor-Based Immunotherapy
3.3. Microbiome Implications in Allo-HSCT (AHSCT)
3.4. Microbiome Implications in Probiotic Immunotherapy
3.4.1. Liver Cancer
3.4.2. Breast Cancer (BC)
3.4.3. Colorectal Cancer (CRC)
3.4.4. Oral Cancer
3.5. Microbiome Implications in CpG-Oligonucleotide (CpG ODN) Immunotherapy
3.6. Microbiome Implications in Adoptive Cell Therapy (ACT)
3.7. Microbiome Implications in Fecal Microbiota Transplantation (FMT)
3.8. Microbiome—Host Crosstalk, Signalling and Immunomodulation
3.9. Microbiome and Cancer Prevention
4. Summary and Concluding Remarks
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| CTLA-4 | Cytotoxic T-lymphocyte antigen-4 |
| GBA | Gut Brain Axis |
| EBV | Epstein–Barr virus |
| HIV | Human Immunodeficiency Virus |
| HPV | Human Papilloma |
| HPTLV-1 | Type 1 human T-cell lymphotropic virus |
| Hb-B | Hepatitis B |
| Hb-C | Hepatitis C |
| DMAB | 3, 2′-dimethyl-4-aminobiphenol |
| NHMI | Nitrosoheptamethyleneimine |
| DEN | N-nitrosodiethylamine |
| COPD | Chronic obstructive pulmonary disease |
| CCl4 | Carbon Tetra Chloride |
| DCA | Deoxycholic Acid |
| DNA | Deoxyribonucleic Acid |
| MALT | Mucosa-associated lymphoid tissue |
| ISPID | Immunoproliferative small intestinal disease |
| TNF | Tumor Necrosis Factor |
| MAMP | Microbiome associated molecular pattern |
| TLR | Toll Like Receptors |
| LPS | Lipopolysaccharides |
| CRC | Colorectal Cancer |
| SCFA | Short Chain Fatty Acids |
| DSS | Dextran Sulfate Sodium |
| AOM | Azoxymethane |
| TIGIT | T cell immunoreceptor with Ig and ITIM domains |
| IL | Interleukin |
| BC | Breast Cancer |
| rRNA | ribosomal Ribonucleic Acid |
| NAFLD | Non-alcoholic liver fatty liver disease |
| HCC | Hepatocellular carcinoma |
| mAbs | monoclonal Antibodies |
| PD-1 | programmed cell death protein1 |
| PD-L1 | programmed cell death ligand protein 1 |
| ICIs | Immune Check point Inhibitors |
| TME | Tumor Microenvironment |
| APCs | Antigen Presenting Cells |
| FMT | Faecal Microbial Transplantation |
| HSCT | hematopoietic stem cell transplantation |
| MDACC | MD Anderson Cancer Centre |
| ICB | immune checkpoint blockade |
| GVHD | Graft versus host disease |
| Th17 | Thyroxin receptor 17 |
| pDC | plasmacytoid DC (Dendritic cells) |
| TILs | Tumor Infiltrating Lymphocytes |
| BiTEs | Bispecific T-cell engagers |
| CAR | Chimeric Antigen Receptor |
| ACT | Adoptive Cell Therapy |
| CDI | Clostridium difficali infection |
| CNS | Central Nervous System |
| ANS | Autonomous Nervous System |
| ENS | Enteric Nervous System |
| GABA | Gama Amino Butyric Acid |
| mTOR | mammalian target of rapamycin |
| GALT | Gut Associated lymphoid tissues |
| NK | Natural Killer cells |
| DC | Dendritic cells |
| MPL | Monophosphoryl Lipid |
References
- Zitvogel, L.; Daillère, R.; Roberti, M.P.; Routy, B.; Kroemer, G. Anticancer effects of the microbiome and its products. Nat. Rev. Genet. 2017, 15, 465–478. [Google Scholar] [CrossRef]
- Brown, E.M.; Sadarangani, M.; Finlay, B.B. The role of the immune system in governing host-microbe interactions in the intestine. Nat. Immunol. 2013, 14, 660–667. [Google Scholar] [CrossRef]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.F. Development of intestinal microbiota in in-fants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef]
- Walker, R.W.; Clemente, J.C.; Peter, I.; Loos, R.J. The prenatal gut microbiome: Are we colonized with bacteria in utero? Pediatric Obes. 2017, 12, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Purchiaroni, F.; Tortora, A.; Gabrielli, M.; Bertucci, F.; Gigante, G.; Ianiro, G.; Ojetti, V.; Scarpellini, E.; Gasbarrini, A. The role of intestinal microbiota and the immune system. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 323–333. [Google Scholar] [PubMed]
- Wu, H.-J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kivit, S.; Tobin, M.C.; Forsyth, C.; Keshavarzian, A.; Landay, A.L. Regulation of Intestinal Immune Responses through TLR Activation: Implications for Pro- and Prebiotics. Front. Immunol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nu-trients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Cussotto, S.; Sandhu, K.V.; Dinan, T.G.; Cryan, J.F. The Neuroendocrinology of the Microbiota-Gut-Brain Axis: A Behavioural Perspective. Front. Neuroendocr. 2018, 51, 80–101. [Google Scholar] [CrossRef]
- Ahmad, A.F.; Dwivedi, G.; O’Gara, F.; Caparros-Martin, J.; Ward, N.C. The gut microbiome and cardiovascular disease: Cur-rent knowledge and clinical potential. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H923–H938. [Google Scholar] [CrossRef]
- Sharma, M.; Li, Y.; Stoll, M.L.; Tollefsbol, T.O. The epigenetic connection between the gut microbiome in obesity and diabe-tes. Front. Genet. 2020, 10, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapp, M.; Aurora, N.; Herrera, L.; Bhatia, M.; Wilen, E.; Wakefield, S. Gut microbiota’s effect on mental health: The gut-brain axis. Clin. Pr. 2017, 7, 987. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.; Gasbarrini., A.; Mele, M.C. What is the healthy gut mi-crobiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sears, C.L.; Garrett, W.S. Microbes, Microbiota, and Colon Cancer. Cell Host Microbe 2014, 15, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsilimigras, M.C.B.; Fodor, A.; Jobin, C. Carcinogenesis and therapeutics: The microbiota perspective. Nat. Microbiol. 2017, 2, 1–10. [Google Scholar] [CrossRef]
- Boursi, B.; Mamtani, R.; Haynes, K.; Yang, Y.-X. Recurrent antibiotic exposure may promote cancer formation—Another step in understanding the role of the human microbiota? Eur. J. Cancer 2015, 51, 2655–2664. [Google Scholar] [CrossRef] [Green Version]
- Saus, E.; Iraola-Guzmán, S.; Willis, J.R.; Brunet-Vega, A.; Gabaldón, T. Microbiome and colorectal cancer: Roles in carcinogenesis and clinical potential. Mol. Asp. Med. [CrossRef]
- Yang, Y.; Jobin, C. Novel insights into microbiome in colitis and colorectal cancer. Curr. Opin. Gastroenterol. 2017, 33, 422–427. [Google Scholar] [CrossRef]
- Brennan, C.A.; Garrett, W.S. Gut Microbiota, Inflammation, and Colorectal Cancer. Annu. Rev. Microbiol. 2016, 70, 395–411. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, D.V. The Human Gutome: Nutrigenomics of the Host–Microbiome Interactions. OMICS: A J. Integr. Biol. 2011, 15, 419–430. [Google Scholar] [CrossRef]
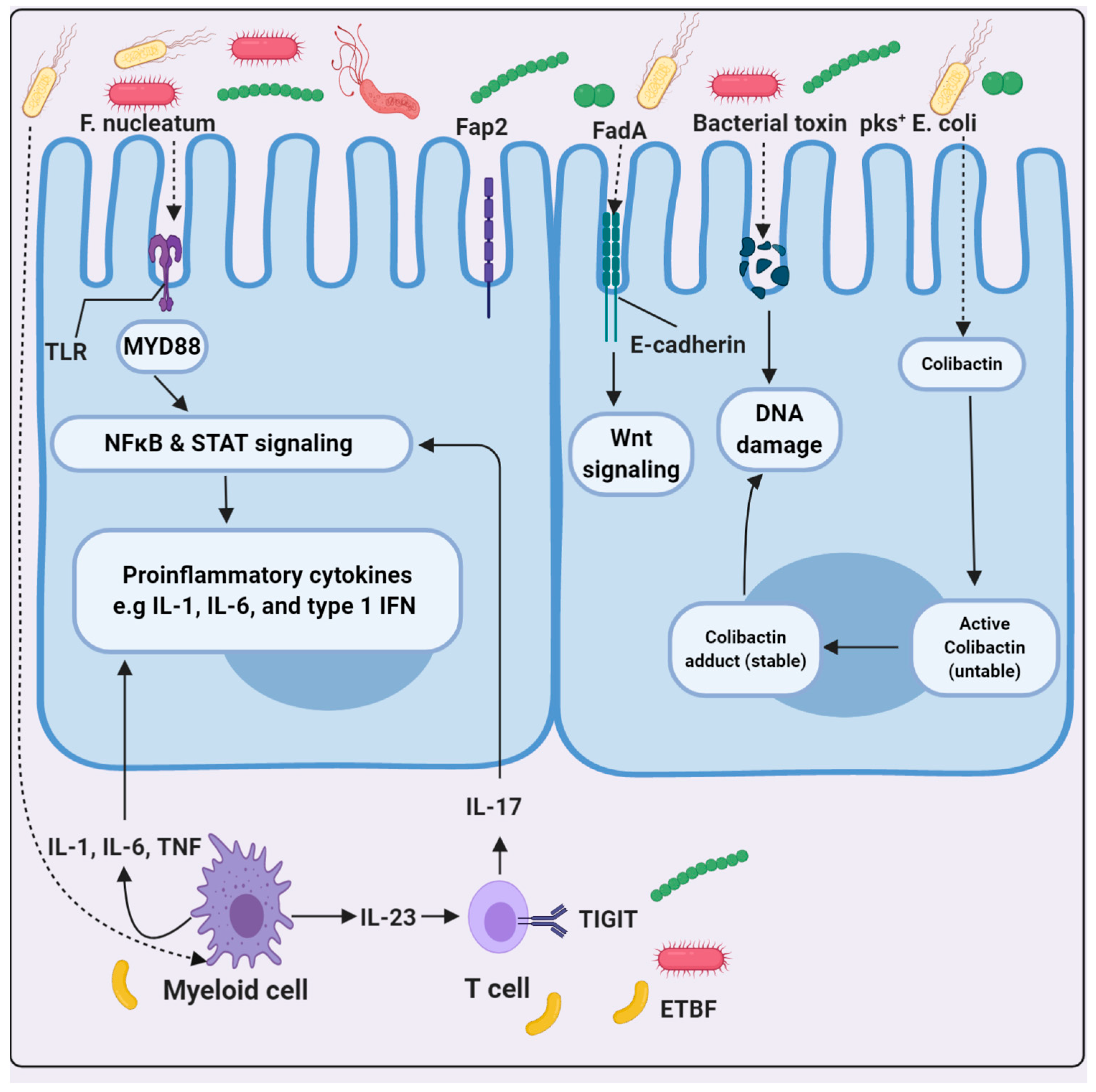
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacteriumnucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2011, 22, 292–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamurugan, R.; Rajendiran, E.; George, S.; Samuel, G.V.; Ramakrishna, B.S. Real-time polymerase chain reaction quantifi-cation of specific butyrate-producing bacteria, Desulfovibrio and Enterococcusfaecalis in the feces of patients with colorectal cancer. J. Gastroenterol. Hepatol. 2008, 23, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota be-tween colorectal cancer patients and healthy volunteers. ISME J. 2012, 6, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Yang, X.; Zhang, R.; Li, J.; Xiao, X.; Hu, Y.; Chen, Y.; Yang, F.; Lu, N.; Wang, Z.; et al. Dysbiosis Signature of Fecal Microbiota in Colorectal Cancer Patients. Microb. Ecol. 2013, 66, 462–470. [Google Scholar] [CrossRef]
- Lee, S.A.; Liu, F.; Riordan, S.M.; Lee, C.S.; Zhang, L. Global Investigations of Fusobacterium nucleatum in Human Colorectal Cancer. Front. Oncol. 2019, 9, 566. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Ji, G. Fusobacterium nucleatum-positive colorectal cancer. Oncol. Lett. 2019, 18, 975–982. [Google Scholar]
- Khan, A.A.; Shrivastava, A.; Khurshid, M. Normal to cancer microbiome transformation and its implication in cancer diag-nosis. Biochim. Biophys. Acta. 2012, 1826, 331–337. [Google Scholar]
- Sharma, V.; Chauhan, V.S.; Nath, G.; Kumar, A.; Shukla, V.K. Role of bile bacteria in gallbladder carcinoma. Hepato Gastroenterol. 2007, 54, 1622–1625. [Google Scholar]
- Narikiyo, M.; Tanabe, C.; Yamada, Y.; Igaki, H.; Tachimori, Y.; Kato, H.; Muto, M.; Montesano, R.; Sakamoto, H.; Nakajima, Y.; et al. Frequent and preferential infection of Treponema denticola, Streptococcus mitis, and Streptococcus anginosus in esophageal cancers. Cancer Sci. 2004, 95, 569–574. [Google Scholar] [CrossRef]
- Macfarlane, S.; Furrie, E.; Macfarlane, G.T.; Dillon, J.F. Microbial Colonization of the Upper Gastrointestinal Tract in Patients with Barrett’s Esophagus. Clin. Infect. Dis. 2007, 45, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.A.; Murphy, S.J.; Johnston, B.T.; Watson, R.G.; Ferguson, H.R.; Bamford, K.B.; Ghazy, A.; McCarron, P.; McGui-gan, J.; Reynolds, J.V.; et al. Relationship between Helicobacter pylori infection and gastric atrophy and the stages of the oe-sophageal inflammation, metaplasia, adenocarcinoma sequence: Results from the FINBAR case-control study. Gut 2008, 57, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Mager, D.L.; Haffajee, A.; Devlin, F.F.M.P.M.; Norris, C.M.; Posner, M.R.; Goodson, J.M. The salivary microbiota as a diagnostic indicator of oral cancer: A descriptive, non-randomized study of cancer-free and oral squamous cell carcinoma subjects. J. Transl. Med. 2005, 3, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.-L.; Shi, Y.; Zhou, L.; Wu, C.-P.; Cao, P.-Y.; Tao, L.; Xu, C.; Hou, D.-S.; Wang, Y.-Z. The Composition of Microbiome in Larynx and the Throat Biodiversity between Laryngeal Squamous Cell Carcinoma Patients and Control Population. PLoS ONE 2013, 8, e66476. [Google Scholar] [CrossRef]
- Farrell, J.J.; Zhang, L.; Zhou, H.; Chia, D.; Elashoff, D.; Akin, D.; Paster, B.J.; Joshipura, K.; Wong, D.T. Variations of oral mi-crobiota are associated with pancreatic diseases including pancreatic cancer. Gut 2012, 61, 582–588. [Google Scholar] [CrossRef]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; Delany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 2013, 98, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, S.S.; Doedens, A.; Burns, M.B. The promise and challenge of cancer microbiome research. Genome Biol. 2020, 21, 1–22. [Google Scholar] [CrossRef]
- Kim, N.H.; Park, J.P.; Jeon, S.H.; Lee, Y.J.; Choi, H.J.; Jeong, K.M.; Lee, J.G.; Choi, S.P.; Lim, J.H.; Kim, Y.H.; et al. Purulent Pericarditis Caused by Group G Streptococcus as an Initial Presentation of Colon Cancer. J. Korean Med. Sci. 2002, 17, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, K.; Konishi, F.; Mitsuoka, T.; Terada, A.; Itoh, K.; Narushima, S.; Kumemura, M.; Kimura, H. Factors influencing the development of sigmoid colon cancer: Bacteriologic and biochemical studies. Cancer 1996, 8, 1701–1706. [Google Scholar] [CrossRef]
- Scanlan, P.D.; Shanahan, F.; Clune, Y.; Collins, J.K.; O’Sullivan, G.C.; O’Riordan, M.; Holmes, E.; Wang, Y.; Marchesi, J.R. Culture-independent analysis of the gut microbiota in colorectal cancer and polyposis. Environ. Microbiol. 2008, 10, 789–798. [Google Scholar] [CrossRef]
- Nath, G.; Gulati, A.K.; Shukla, V.K. Role of bacteria in carcinogenesis, with special reference to carcinoma of the gallbladder. World J. Gastroenterol. WJG 2010, 16, 5395. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Severgnini, M.; Pecere, S.; Ponziani, F.R.; Boškoski, I.; Larghi, A.; Quaranta, G.; Masucci, L.; Ianiro, G.; Camboni, T.; et al. Esophageal microbiome signature in patients with Barrett’s esophagus and esophageal adenocarcinoma. PLoS ONE 2020, 15, e0231789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, Y.; Zheng, H.J.; Zhang, C.P. The Oral Microbiota May Have Influence on Oral Cancer. Front. Cell. Infect. Microbiol. 2020, 9, 476. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Shi, Y.; Zhou, X.; Wu, C.; Cao, P.; Xu, C.; Hou, D.; Wang, Y.; Zhou, L. Microbiota in the Throat and Risk Factors for Laryngeal Carcinoma. Appl. Environ. Microbiol. 2014, 80, 7356–7363. [Google Scholar] [CrossRef] [Green Version]
- Tuominen, H.; Rautava, J. Oral Microbiota and Cancer Development. Pathobiology 2020, 1–11. [Google Scholar] [CrossRef]
- Plottel, C.S.; Blaser, M.J. Microbiome and Malignancy. Cell Host Microbe 2011, 10, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Gao, J.; Zhou, Q.; Liu, S.; Wen, C.; Yang, X. Role of intestinal flora in colorectal cancer from the metabolite perspec-tive: A systematic review. Cancer Manag. Res. 2018, 10, 199. [Google Scholar] [CrossRef] [Green Version]
- Oke, S.; Martin, A. Insights into the role of the intestinal microbiota in colon cancer. Ther. Adv. Gastroenterol. 2017, 10, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Francescone, R.; Hou, V.; Grivennikov, S.I. Microbiome, Inflammation, and Cancer. Cancer J. 2014, 20, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.S.; Watanabe, K. Effect of Intestinal Microflora on 3,2′-Dimethyl-4-aminobiphenyl-Induced Carcinogenesis in F344 Rats2. J. Natl. Cancer Inst. 1978, 61, 1269–1271. [Google Scholar] [CrossRef]
- Schreiber, H.; Nettesheim, P.; Lijinsky, W.; Richter, C.B.; Walburg, J.H.E. Induction of Lung Cancer in Germfree, Specific-Pathogen-Free, and Infected Rats by N-Nitrosoheptamethyleneimine: Enhancement by Respiratory Infection2. J. Natl. Cancer Inst. 1972, 49, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Lofgren, J.L.; Whary, M.T.; Ge, Z.; Muthupalani, S.; Taylor, N.S.; Mobley, M.; Potter, A.; Varro, A.; Eibach, D.; Suerbaum, S.; et al. Lack of commensal flora in Helicobacter pylori-infected INS-GAS mice reduces gastritis and delays intraepithelial neo-plasia. Gastroenterology 2011, 140, 210–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.W.; Rickman, B.; Rogers, A.B.; Ge, Z.; Wang, T.C.; Fox, J.G. Helicobacter pylori eradication prevents progression of gas-tric cancer in hypergastrinemic INS-GAS mice. Cancer Res. 2008, 68, 3540–3548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dapito, D.H.; Mencin, A.; Gwak, G.-Y.; Pradere, J.-P.; Jang, M.-K.; Mederacke, I.; Caviglia, J.M.; Khiabanian, H.; Adeyemi, A.; Bataller, R.; et al. Promotion of Hepatocellular Carcinoma by the Intestinal Microbiota and TLR4. Cancer Cell 2012, 21, 504–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.-X.; Yan, H.-X.; Liu, Q.; Yang, W.; Wu, H.-P.; Dong, W.; Tang, L.; Lin, Y.; He, Y.-Q.; Zou, S.-S.; et al. Endotoxin accumulation prevents carcinogen-induced apoptosis and promotes liver tumorigenesis in rodents. Hepatology 2010, 52, 1322–1333. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef]
- Li, Y.; Kundu, P.; Seow, S.W.; de Matos, C.T.; Aronsson, L.; Chin, K.C.; Kärre, C.; Sven Pettersson, S.; Greicius, G. Gut microbiota accelerate tumor growth via c-jun and STAT3 phosphorylation in APCMin/+ mice. Carcinogenesis 2012, 33, 1231–1238. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Y.; Shaw, M.H.; Redondo, G.; Nunez, G. The innate immune receptor Nod1 protects the intestine from inflamma-tion-induced tumorigenesis. Cancer Res. 2008, 68, 10060–10067. [Google Scholar] [CrossRef] [Green Version]
- Kostovcikova, K.; Kverka, M.; Zakostelska, Z.; Hudcovic, T.; Hrncir, T.; Stepankova, R.; Rossmann, P.; Ridl, J.; Kostovcik, M.; Mrazek, J.; et al. Altered Gut Microbiota Promotes Colitis-Associated Cancer in IL-1 Receptor–Associated Kinase M–Deficient Mice. Inflamm. Bowel Dis. 2013, 19, 1266–1277. [Google Scholar] [CrossRef] [Green Version]
- Couturier-Maillard, A.; Thomas Secher, T.; Rehman, A.; Normand, S.; Arcangelis, A.; Haesler, R.; Huot, L.; Grandjean, T.; Bressenot, A.; Delanoye-Crespin, A.; et al. NOD2-mediated dysbiosis predisposes mice to transmissible colitis and colorec-tal cancer. J. Clin. Investig. 2013, 123, 700–711. [Google Scholar]
- Cuzick, J.; Otto, F.; Baron, J.A.; Brown, P.H.; Burn, J.; Greenwald, P.; Jankowski, J.; La Vecchia, C.; Meyskens, F.; Senn, H.J.; et al. Aspirin and non-steroidal anti-inflammatory drugs for cancer prevention: An international consensus statement. Lancet Oncol. 2009, 10, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Bosetti, C.; Gallus, S.; La Vecchia, C. Aspirin and cancer risk: An updated quantitative review to 2005. Cancer Causes Control 2006, 17, 871–888. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; Perez-Chanona, E.; Mühlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.-J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal Inflammation Targets Cancer-Inducing Activity of the Microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooks, M.G.; Garrett, W.S. Bacteria, food, and cancer. F1000 Biol. Rep. 2011, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Willett, W.C. Diet and cancer: An evolving picture. JAMA 2005, 293, 233–234. [Google Scholar] [CrossRef]
- Ma, J.-L.; Zhang, L.; Brown, L.M.; Li, J.-Y.; Shen, L.; Pan, K.-F.; Liu, W.-D.; Hu, Y.; Han, Z.-X.; Crystal-Mansour, S.; et al. Fifteen-Year Effects of Helicobacter pylori, Garlic, and Vitamin Treatments on Gastric Cancer Incidence and Mortality. J. Natl. Cancer Inst. 2012, 104, 488–492. [Google Scholar] [CrossRef]
- Wong, B.C.; Lam, S.K.; Wong, W.M.; Chen, J.S.; Zheng, T.T.; Feng, R.E.; Lai, K.C.; Hu, W.H.; Yuen, S.T.; Leung, S.Y.; et al. Hel-icobacter pylori eradication to prevent gastric cancer in a high-risk region of China: A randomized controlled trial. JAMA 2004, 291, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Peek, R.M.; Blaser, M.J. Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2002, 2, 28–37. [Google Scholar] [CrossRef]
- Fox, J.G.; Wang, T.C. Inflammation, atrophy, and gastric cancer. J. Clin. Investig. 2007, 117, 60–69. [Google Scholar] [CrossRef]
- Lecuit, M.; Abachin, E.; Martin, A.; Poyart, C.; Pochart, P.; Suarez, F.; Bengoufa, D.; Feuillard, J.; Lavergne, A.; Gordon, J.I.; et al. Immunoproliferative Small Intestinal Disease Associated withCampylobacter jejuni. N. Engl. J. Med. 2004, 350, 239–248. [Google Scholar] [CrossRef]
- Senff, N.J.; Noordijk, E.M.; Kim, Y.H.; Bagot, M.; Berti, E.; Cerroni, L.; Dummer, R.; Duvic, M.; Hoppe, R.T.; Pimpinelli, N.; et al. European Organization for Research and Treatment of Cancer and International Society for Cutaneous Lymphoma con-sensus recommendations for the management of cutaneous B-cell lymphomas. Blood 2008, 112, 1600–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreri, A.J.; Govi, S.; Pasini, E.; Mappa, S.; Bertoni, F.; Zaja, F.; Montalbán, C.; Stelitano, C.; Cabrera, M.E.; Giordano Resti, A. Chlamydophila psittaci eradication with doxycycline as first-line targeted therapy for ocular adnexae lymphoma: Final results of an international phase II trial. J. Clin. Oncol. 2012, 30, 2988–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islami, F.; Kamangar, F. Helicobacter pylori and Esophageal Cancer Risk: A Meta-analysis. Cancer Prev. Res. 2008, 1, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caygill, C.; Hill, M.; Braddick, M.; Sharp, J. Cancer mortality in chronic typhoid and paratyphoid carriers. Lancet 1994, 343, 83–84. [Google Scholar] [CrossRef]
- Welton, J.; Marr, J.; Friedman, S. Association between hepatobiliary cancer and typhoid carrier state. Lancet 1979, 313, 791–794. [Google Scholar] [CrossRef]
- Rao, V.P.; Poutahidis, T.; Ge, Z.; Nambiar, P.R.; Horwitz, B.H.; Fox, J.G.; Erdman, S.E. Proinflammatory CD4+CD45RBhi Lymphocytes Promote Mammary and Intestinal Carcinogenesis in ApcMin/+ Mice. Cancer Res. 2006, 66, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, J.M.; Fox, J.G.; Anver, M.R.; Haines, D.C.; George, C.V.; Collins, M.J.J.; Gorelick, P.L.; Nagashima, K.; Gonda, M.A.; Gilden, R.V. Chronic active hepatitis and associated liver tumors in mice caused by a persistent bacterial infection with a novel Helicobacter species. J. Natl. Cancer Inst. 1994, 86, 1222–1227. [Google Scholar] [CrossRef]
- Erdman, S.E.; Rao, V.P.; Poutahidis, T.; Rogers, A.B.; Taylor, C.L.; Jackson, E.A.; Ge, Z.; Lee, C.W.; Schauer, D.B.; Wogan, G.N. Nitric oxide and TNF-α trigger colonic inflammation and carcinogenesis in Helicobacter hepaticus-infected, Rag2-deficient mice. Proc. Natl. Acad. Sci. USA 2009, 106, 1027–1032. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.S.; Narisawa, T.; Wright, P.; Vukusich, D.; Weisburger, J.H.; Wynder, E.L. Colon carcinogenesis with azoxymethane and dimethylhydrazine in germ-free rats. Cancer Res. 1975, 35, 287–290. [Google Scholar]
- Reddy, B.S.; Weisburger, J.H.; Narisawa, T.; Wynder, E.L. Colon carcinogenesis in germ-free rats with 1,2-dimethylhydrazine and N-methyl-n’-nitro-N-nitrosoguanidine. Cancer Res. 1974, 34, 2368–2372. [Google Scholar]
- Vannucci, L.; Stepankova, R.; Kozakova, H.; Fiserova, A.; Rossmann, P.; Tlaskalova-Hogenova, H. Colorectal carcinogenesis in germ-free and conventionally reared rats: Different intestinal environments affect the systemic immunity. Int. J. Oncol. 2008, 32, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dove, W.F.; Clipson, L.; A Gould, K.; Luongo, C.; Marshall, D.J.; Moser, A.R.; A Newton, M.; Jacoby, R.F. Intestinal neoplasia in the ApcMin mouse: Independence from the microbial and natural killer (beige locus) status. Cancer Res. 1997, 57, 812–814. [Google Scholar] [PubMed]
- Grivennikov, S.I.; Wang, K.; Mucida, D.; Stewart, C.A.; Schnabl, B.; Jauch, D.; Taniguchi, K.; Yu, G.-Y.; Oesterreicher, C.H.; Hung, K.E.; et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nat. Cell Biol. 2012, 491, 254–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponziani, F.R.; Nicoletti, A.; Gasbarrini, A.; Pompili, M. Diagnostic and therapeutic potential of the gut microbiota in patients with early hepatocellular carcinoma. Ther. Adv. Med Oncol. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Toffanin, S.; Cornella, H.; Harrington, A.; Llovet, J.; Groszmann, R.; Iwakiri, Y.; Taddei, T. HCC is promoted by bacterial transloca-tion and TLR-4 signaling: A new paradigm for chemoprevention and management. Hepatology 2000, 56, 1998–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse Microbiota Models: Comparing Germ-Free Mice and Antibiotics Treatment as Tools for Modifying Gut Bacteria. Front. Physiol. 2018, 9, 1534. [Google Scholar] [CrossRef] [Green Version]
- Pragman, A.A.; Kim, H.B.; Reilly, C.S.; Wendt, C.; Isaacson, R.E. The lung microbiome in moderate and severe chronic ob-structive pulmonary disease. PLoS ONE 2012, 7, e47305. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.; Murphy, T.F. Infection in the Pathogenesis and Course of Chronic Obstructive Pulmonary Disease. N. Engl. J. Med. 2008, 359, 2355–2365. [Google Scholar] [CrossRef]
- Houghton, A.M. Mechanistic links between COPD and lung cancer. Nat. Rev. Cancer 2013, 13, 233–245. [Google Scholar] [CrossRef]
- Melkamu, T.; Qian, X.; Upadhyaya, P.; O’Sullivan, M.G.; Kassie, F. Lipopolysaccharide enhancesmouse lung tumorigenesis: A model for inflammation-driven lung cancer. Vet. Pathol. 2013, 50, 895–902. [Google Scholar] [CrossRef]
- Ochi, A.; Nguyen, A.H.; Bedrosian, A.S.; Mushlin, H.M.; Zarbakhsh, S.; Barilla, R.; Zambirinis, C.P.; Fallon, N.C.; Rehman, A.; Pylayeva-Gupta, Y.; et al. MyD88 inhibition amplifies dendritic cell capacity to promote pancreaticbcarcinogenesis via Th2 cells. J. Exp. Med. 2012, 209, 1671–1687. [Google Scholar] [CrossRef] [PubMed]
- Michaud, D.S.; Joshipura, K.; Giovannucci, E.; Fuchs, C.S. A Prospective Study of Periodontal Disease and Pancreatic Cancer in US Male Health Professionals. J. Natl. Cancer Inst. 2007, 99, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Ammer-Herrmenau, C.; Pfisterer, N.; Weingarten, M.F.; Neesse, A. The microbiome in pancreatic diseases: Recent advances and future perspectives. United Eur. Gastroenterol. J. 2020, 8, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Seregin, S.S.; Chen, J.; Chen, G.Y. Nod1 Limits Colitis-Associated Tumorigenesis by Regulating IFN-γ Production. J. Immunol. 2016, 196, 5121–5129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udden, S.N.; Peng, L.; Gan, J.-L.; Shelton, J.M.; Malter, J.S.; Hooper, L.V.; Zaki, H. NOD2 Suppresses Colorectal Tumorigenesis via Downregulation of the TLR Pathways. Cell Rep. 2017, 19, 2756–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Elinav, E.; Huber, S.; Booth, C.J.; Strowig, T.; Jin, C.; Eisenbarth, S.C.; Flavell, R.A. Inflammation-induced tumorigen-esis in the colon is regulated by caspase-1 and NLRC. Proc. Natl. Acad. Sci. USA 2010, 107, 21635–21640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaki, H.; Lamkanfi, M.; Kanneganti, T.-D. The Nlrp3 inflammasome: Contributions to intestinal homeostasis. Trends Immunol. 2011, 32, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Zaki, H.; Vogel, P.; Malireddi, R.K.S.; Body-Malapel, M.; Anand, P.K.; Bertin, J.; Green, D.R.; Lamkanfi, M.; Kanneganti, T.-D. The NOD-Like Receptor NLRP12 Attenuates Colon Inflammation and Tumorigenesis. Cancer Cell 2011, 20, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Y.; Liu, M.; Wang, F.; Bertin, J.; Núñez, G. A Functional Role for Nlrp6 in Intestinal Inflammation and Tumorigenesis. J. Immunol. 2011, 186, 7187–7194. [Google Scholar] [CrossRef]
- Zhan, Y.; Chen, P.J.; Sadler, W.D.; Wang, F.; Poe, S.; Núñez, G.; Eaton, K.A.; Chen, G.Y. Gut microbiota protects against gas-trointestinal tumorigenesis caused by epithelial injury. Cancer Res. 2013, 73, 7199–7210. [Google Scholar] [CrossRef] [Green Version]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobac-terium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mima, K.; Sukawa, Y.; Nishihara, R.; Qian, Z.R.; Yamauchi, M.; Inamura, K.; Kim, S.A.; Masuda, A.; Nowak, J.A.; Nosho, K.; et al. Fusobacterium nucleatum and T Cells in Colorectal Carcinoma. JAMA Oncol. 2015, 1, 653–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galon, J.; Costes, A.; Sanchez-Cabo, F.; Kirilovsky, A.; Mlecnik, B.; Lagorce-Pagès, C.; Tosolini, M.; Camus, M.; Berger, A.; Wind, P.; et al. Type, Density, and Location of Immune Cells Within Human Colorectal Tumors Predict Clinical Outcome. Science 2006, 313, 1960–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fragomeni, S.M.; Sciallis, A.; Jeruss, J.S. Molecular Subtypes and Local-Regional Control of Breast Cancer. Surg. Oncol. Clin. North Am. 2018, 27, 95–120. [Google Scholar] [CrossRef]
- Lacey, J.V.; Kreimer, A.R.; Buys, S.S.; Marcus, P.M.; Chang, S.-C.; Leitzmann, M.F.; Hoover, R.N.; Prorok, P.; Berg, C.D.; Hartge, P. Breast cancer epidemiology according to recognized breast cancer risk factors in the Prostate, Lung, Colorectal and Ovarian (PLCO) Cancer Screening Trial Cohort. BMC Cancer 2009, 9, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacteriumnucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its Fad Aadhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar]
- Nougayrède, J.-P.; Homburg, S.; Taieb, F.; Boury, M.; Brzuszkiewicz, E.; Gottschalk, G.; Buchrieser, C.; Hacker, J.; Ulrich Do-brindt, U.; Oswald, E.; et al. Escherichiacoli induces DNA double-strand breaks in eukaryotic cells. Science 2006, 313, 848–851. [Google Scholar] [CrossRef]
- Wang, T.C.; Goldenring, J.R.; Dangler, C.; Ito, S.; Mueller, A.; Jeon, W.K.; Koh, T.J.; Fox, J.G. Mice lacking secretory phospho-lipase A 2 show altered apoptosis and differentiation with Helicobacterfelis infection. Gastroenterology 1998, 114, 675–689. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Medzhitov, R. Toll-like receptors and cancer. Nat. Rev. Cancer 2008, 9, 57–63. [Google Scholar] [CrossRef]
- Thomas, R.M.; Jobin, C. The microbiome and cancer: Is the ‘oncobiome’mirage real? Trends Cancer 2015, 1, 24–35. [Google Scholar] [CrossRef] [Green Version]
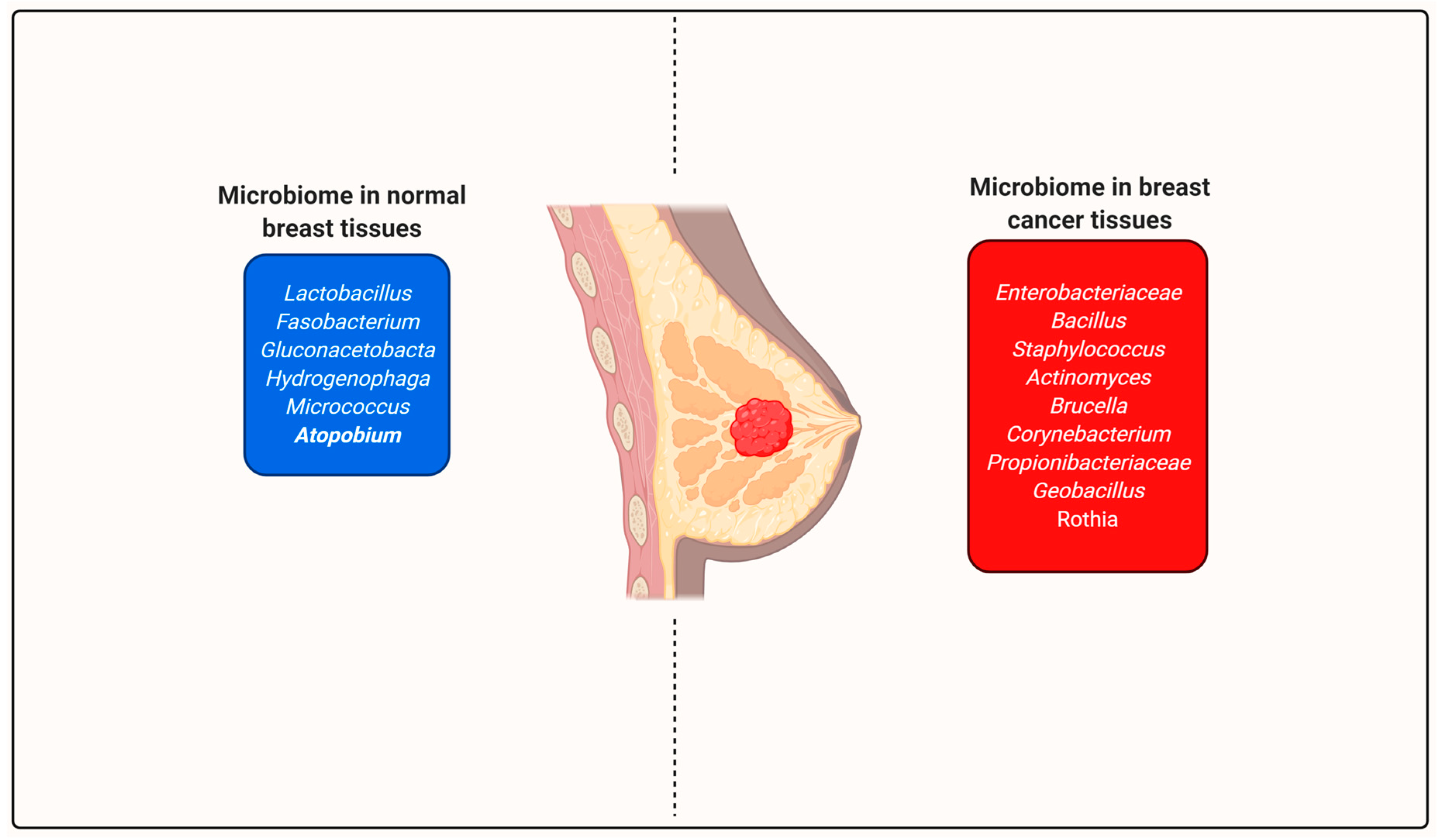
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O’Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of Human Breast Tissue. Appl. Environ. Microbiol. 2014, 80, 3007–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The Microbiota of Breast Tissue and Its Association with Breast Cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, A.-Q.; Li, L. The Potential Role of Probiotics in Cancer Prevention and Treatment. Nutr. Cancer 2016, 68, 535–544. [Google Scholar] [CrossRef]
- Ghosh, T.; Beniwal, A.; Semwal, A.; Navani, N.K. Mechanistic insights into probiotic properties of lactic acid bacteria associ-ated with ethnic fermented dairy products. Front. Microbiol. 2019, 10, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyetayo, V.O.; Oyetayo, F.L. Potential of probiotics as biotherapeutic agents targeting the innate immune system. Afr. J. Biotechnol. 2005, 4, 123–127. [Google Scholar]
- Ohata, H.; Kitauchi, S.; Yoshimura, N.; Mugitani, K.; Iwane, M.; Nakamura, H.; Yoshikawa, A.; Yanaoka, K.; Arii, K.; Tamai, H.; et al. Progression of chronic atrophic gastritis associated with Helicobacter pylori infection increases risk of gastric can-cer. Int. J. Cancer 2004, 109, 138–143. [Google Scholar] [CrossRef]
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome – A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.; Yang, L.; Paster, B.J.; Ganly, I.; Morris, L.; Pei, Z.; Hayes, R.B. Oral Microbiome Profiles: 16S rRNA Pyrosequencing and Microarray Assay Comparison. PLoS ONE 2011, 6, e22788. [Google Scholar] [CrossRef] [Green Version]
- Olsen, I. Yilmaz, Özlem Possible role of Porphyromonas gingivalis in orodigestive cancers. J. Oral Microbiol. 2019, 11, 1563410. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Gail, M.H.; Shi, J.; Klepac-Ceraj, V.; Paster, B.J.; Dye, B.A.; Wang, G.Q.; Wei, W.Q.; Fan, J.H.; Qiao, Y.L.; et al. Associa-tion between upper digestive tract microbiota and cancer-predisposing states in the esophagus and stomach. Cancer Epidemi-ol. Prev. Biomark 2014, 23, 735–741. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, G.; Kreimer, A.R.; Viscidi, R.; Pawlita, M.; Fakhry, C.; Koch, W.M.; Westra, W.H.; Gillison, M.L. Case–Control Study of Human Papillomavirus and Oropharyngeal Cancer. N. Engl. J. Med. 2007, 356, 1944–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, K.; Sonkodi, I.; Szöke, I.; Nagy, E.; Newman, H. The microflora associated with human oral carcinomas. Oral Oncol. 1998, 34, 304–308. [Google Scholar] [CrossRef]
- Gupta, H.; Youn, G.S.; Shin, M.J.; Suk, K.T. Role of Gut Microbiota in Hepatocarcinogenesis. Microorganisms 2019, 7, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharpton, S.R.; Maraj, B.; Harding-Theobald, E.; Vittinghoff, E.; Terrault, N.A. Gut microbiome–targeted therapies in nonal-coholic fatty liver disease: A systematic review, meta-analysis, and meta-regression. Am. J. Clin. Nutr. 2019, 110, 139–149. [Google Scholar] [CrossRef]
- Killian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Araujo, A.R.; Rosso, N.; Bedogni, G.; Tiribelli, C.; Bellentani, S. Global epidemiology of non-alcoholic fatty liver dis-ease/non-alcoholic steatohepatitis: What we need in the future. Liver Int. 2018, 38, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Brunt, E.M.; Wong, V.W.S.; Nobili, V.; Day, C.P.; Sookoian, S.; Maher, J.J.; Bugianesi, E.; Sirlin, C.B.; Neuschwander-Tetri, B.A.; Rinella, M.E. Nonalcoholic fatty liver disease. Nat. Rev. Dis. Primers 2015, 1, 1–22. [Google Scholar] [CrossRef]
- Zhou, R.; Fan, X.; Schnabl, B. Role of the intestinal microbiome in liver fibrosis development and new treatment strategies. Transl. Res. 2019, 209, 22–38. [Google Scholar] [CrossRef]
- De Palma, G.; Nadal, I.; Collado, M.C.; Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br. J. Nutr. 2009, 102, 1154–1160. [Google Scholar] [CrossRef] [Green Version]
- Haran, J.P.; Bhattarai, S.K.; Foley, S.E.; Dutta, P.; Ward, D.V.; Bucci, V.; McCormick, B.A. Alzheimer’s Disease Microbiome Is Associated with Dysregulation of the Anti-Inflammatory P-Glycoprotein Pathway. mBio 2019, 10, e00632-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoudiandehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; Louie, G.; Kueider-Paisley, A.; Moseley, M.A.; Thompson, J.W.; et al. Altered bile acid profile associates with cognitive impairment in Alzheimer’s disease—An emerging role for gut microbiome. Alzheimer’s Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, A.; Bonelli, P.; Tuccillo, F.M.; Goldfine, I.D.; Evans, J.L.; Buonaguro, F.M.; Mancini, A. Role of gut microbiota and oxidative stress in the progression of non-alcoholic fatty liver disease to hepatocarcinoma: Current and innovative thera-peutic approaches. Redox biol. 2018, 15, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Betrapally, N.S.; Gillevet, P.M.; Bajaj, J.S. Changes in the intestinal microbiome and alcoholic and nonalcoholic liver diseas-es: Causes or effects? Gastroenterology 2016, 150, 1745–1755. [Google Scholar] [CrossRef] [Green Version]
- Schwimmer, J.B.; Johnson, J.S.; Angeles, J.E.; Behling, C.; Belt, P.H.; Borecki, I.; Bross, C.; Durelle, J.; Goyal, N.P.; Hamilton, G.; et al. Microbiome signatures associated with steatohepatitis and moderate to severe fibrosis in children with nonalco-holic fatty liver disease. Gastroenterology 2019, 157, 1109–1122. [Google Scholar] [CrossRef] [Green Version]
- Waterboer, T.; Dondog, B.; Michael, K.M.; Michel, A.; Schmitt, M.; Vaccarella, S.; Franceschi, S.; Clifford, G.; Pawlita, M. Dried Blood Spot Samples for Seroepidemiology of Infections with Human Papillomaviruses, Helicobacter pylori, Hepatitis C Virus, and JC Virus. Cancer Epidemiol. Biomark. Prev. 2012, 21, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; Xie, G.; Jia, W. Bile Acid–microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111. [Google Scholar] [CrossRef] [Green Version]
- Peck, S.C.; Denger, K.; Burrichter, A.; Irwin, S.M.; Balskus, E.P.; Schleheck, D. A glycyl radical enzyme enables hydrogen sulfide production by the human intestinal bacterium Bilophilawadsworthia. Proc. Natl. Acad. Sci. USA 2019, 116, 3171–3176. [Google Scholar] [CrossRef] [Green Version]
- Ji, S.; Choi, Y.S. Bacterial invasion and persistence: Critical events in the pathogenesis of periodontitis? J. Periodontal Res. 2014, 50, 570–585. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a Preventable Disease that Requires Major Lifestyle Changes. Pharm. Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef]
- Hounsome, N.; Tomos, D.; Edwards-Jones, G. Plant Metabolites and Nutritional Quality of Vegetables. J. Food Sci. 2008, 73, R48–R65. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermudez-Humaran, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy me-tabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Factories 2017, 16, 1–10. [Google Scholar]
- Sanders, M.E.; Klaenhammer, T.R. Invited Review: The Scientific Basis of Lactobacillus acidophilus NCFM Functionality as a Probiotic. J. Dairy Sci. 2001, 84, 319–331. [Google Scholar] [CrossRef]
- Im, E.; Pothoulakis, C. Recent advances in Saccharomyces boulardii research. Gastroentérologie Clinique et Biologique 2010, 34, S62–S70. [Google Scholar] [CrossRef]
- Roderburg, C.; Luedde, T. The role of the gut microbiome in the development and progression of liver cirrhosis and hepa-tocellular carcinoma. Gut Microbes 2014, 5, 441–445. [Google Scholar] [CrossRef]
- Yu, L.X.; Schwabe, R.F. The gut microbiome and liver cancer: Mechanisms and clinical translation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 527. [Google Scholar] [CrossRef]
- Peggs, K.S.; Quezada, S.A.; Allison, J.P. Cancer immunotherapy: Co-stimulatory agonists and co-inhibitory antagonists. Clin. Exp. Immunol. 2009, 157, 9–19. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Immune checkpoint blockade: A common denominator approach to cancer thera-py. Cancer Cell 2015, 27, 450–461. [Google Scholar] [CrossRef] [Green Version]
- Mazzarella, L.; Duso, B.A.; Trapani, D.; Belli, C.; D’Amico, P.; Ferraro, E.; Viale, G.; Curigliano, G. The evolving landscape of ‘next-generation’immune checkpoint inhibitors: A review. Eur. J. Cancer 2019, 117, 14–31. [Google Scholar] [CrossRef]
- Huang, J.; Jiang, Z.; Wang, Y.; Fan, X.; Cai, J.; Yao, X.; Liu, L.; Huang, J.; He, J.; Xie, C.; et al. Modulation of gut microbiota to over-come resistance to immune checkpoint blockade in cancer immunotherapy. Curr. Opin. Pharmacol. 2020, 54, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lengfelder, I.; Sava, I.G.; Hansen, J.J.; Kleigrewe, K.; Herzog, J.; Neuhaus, K.; Hofmann, T.; Sartor, R.B.; Haller, D. Complex Bacterial Consortia Reprogram the Colitogenic Activity of Enterococcus faecalis in a Gnotobiotic Mouse Model of Chronic, Immune-Mediated Colitis. Front. Immunol. 2019, 10, 1420. [Google Scholar] [CrossRef] [PubMed]
- Paul, G.; Khare, V.; Gasche, C. Inflamed gut mucosa: Downstream of interleukin-10. Eur. J. Clin. Investig. 2012, 42, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.V.; Xiang, W.V.; Kwak, C.; Yang, Y.; Lin, X.W.; Ota, M.; Sarpel, U.; Rifkin, D.B.; Xu, R.; Littman, D.R. GPR15-Mediated Homing Controls Immune Homeostasis in the Large Intestine Mucosa. Science 2013, 340, 1456–1459. [Google Scholar] [CrossRef] [Green Version]
- Sopta, M.L. Toll Like Receptors (Tlr) in Autoimmune Diseases and Atherosclerosis. Ph.D. Thesis, School of Medicine, University of Zagreb, Zagreb, Croatia, 2016. [Google Scholar]
- Alexander, J.L.; Wilson, I.D.; Teare, J.; Marchesi, J.R.; Nicholson, J.K.; Kinross, J.M. Gut microbiota modulation of chemo-therapy efficacy and toxicity. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 356–365. [Google Scholar] [CrossRef]
- Ma, W.; Mao, Q.; Xia, W.; Dong, G.; Yu, C.; Jiang, F. Gut Microbiota Shapes the Efficiency of Cancer Therapy. Front. Microbiol. 2019, 10, 1050. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Rath, A.K.; Chaudhary, A.P.; Thakar, A.; Mohanti, B.K.; Bahadur, S. Lactobacillus brevis CD2 lozenges reduce radi-ation- and chemotherapy-induced mucositis in patients with head and neck cancer: A randomized double-blind placebo-controlled study. Eur. J. Cancer 2012, 48, 875–881. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The Influence of the Gut Microbiome on Can-cer, Immunity, and Cancer Immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal Bacteria Control Cancer Response to Therapy by Modulating the Tumor Microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Paulos, C.M.; Wrzesinski, C.; Kaiser, A.; Hinrichs, C.S.; Chieppa, M.; Cassard, L.; Palmer, D.C.; Boni, A.; Muranski, P.; Yu, Z.; et al. Microbial translocation augments the function of adoptively transferred self/ tumor-specific CD8+ T cells via TLR4 sig-naling. J. Clin. Investig. 2007, 117, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Le Roux, K.; Monot, C.; Boselli, L.; Routier, E.; Cassard, L.; Collins, M.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Jenq, R.R.; Taur, Y.; Devlin, S.M.; Ponce, D.M.; Goldberg, J.D.; Ahr, K.F.; Littmann, E.R.; Ling, L.; Gobourne, A.C.; Miller, L.C.; et al. Intestinal Blautia is associated with reduced death from graft-versus-host disease. Biol. Blood Marrow Transpl. 2015, 21, 1373–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubin, K.; Callahan, M.K.; Ren, B.; Khanin, R.; Viale, A.; Ling, L.; No, D.; Gobourne, A.; Littmann, E.; Huttenhower, C.; et al. Intestinal microbiome analyses identify melanoma patients at risk for checkpointblockade-induced colitis. Nat. Commun. 2016, 7, 0391. [Google Scholar] [CrossRef] [Green Version]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [Green Version]
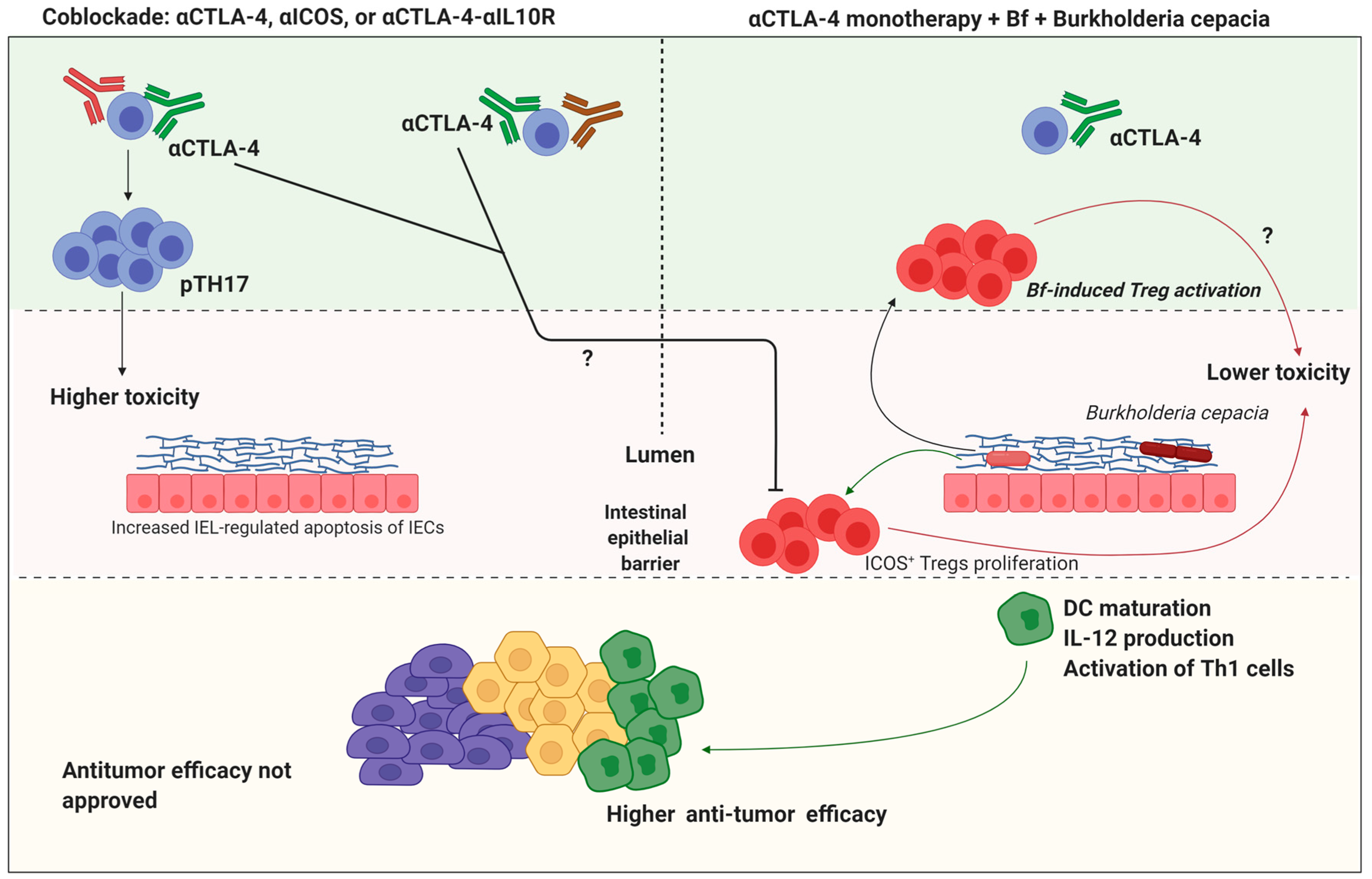
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.M.; et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef] [Green Version]
- Peled, J.; Devlin, S.M.; Staffas, A.; Lumish, M.; Khanin, R.; Littmann, E.R.; Ling, L.; Kosuri, S.; Maloy, M.; Slingerland, J.B.; et al. Intestinal Microbiota and Relapse After Hematopoietic-Cell Transplantation. J. Clin. Oncol. 2017, 35, 1650–1659. [Google Scholar] [CrossRef] [Green Version]
- Frankel, A.E.; Coughlin, L.A.; Kim, J.; Froehlich, T.W.; Xie, Y.; Frenkel, E.P.; Koh, A.Y. Metagenomic Shotgun Sequencing and Unbiased Metabolomic Profiling Identify Specific Human Gut Microbiota and Metabolites Associated with Immune Checkpoint Therapy Efficacy in Melanoma Patients. Neoplasia 2017, 19, 848–855. [Google Scholar] [CrossRef]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.-L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti–PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Routy, B.; Le Chatelier, E.; DeRosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1–based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansom, D.M.; Walker, L.S. The role of CD28 and cytotoxic T-lymphocyte antigen-4 (CTLA-4) in regulatory T-cell biology. Immunol. Rev. 2006, 212, 131–148. [Google Scholar] [CrossRef]
- Gasser, M.; Waaga-Gasser, A.M. Therapeutic antibodies in cancer therapy. In Protein Targeting Compounds; Springer: Berlin/Heidelberg, Germany, 2016; pp. 95–120. [Google Scholar]
- Yi, M.; Qin, S.; Chu, Q.; Wu, K. The role of gut microbiota in immune checkpoint inhibitor therapy. HepatoBiliary Surg. Nutr. 2018, 7, 481–483. [Google Scholar] [CrossRef]
- Seow, S.W.; Rahmat, J.N.B.; Mohamed, A.A.K.; Mahendran, R.; Lee, Y.K.; Bay, B.H. Lactobacillus species is more cytotoxic to human bladder cancer cells than Mycobacterium Bovis (bacillus Calmette-Guerin). J. Urol. 2002, 168, 2236–2239. [Google Scholar] [CrossRef]
- Seidel, J.A.; Otsuka, A.; Kabashima, K. Anti-PD-1 and Anti-CTLA-4 Therapies in Cancer: Mechanisms of Action, Efficacy, and Limitations. Front. Oncol. 2018, 8, 86. [Google Scholar] [CrossRef]
- Zitvogel, L.; Ma, Y.; Raoult, D.; Kroemer, G.; Gajewski, T.F. The microbiome in cancer immunotherapy: Diagnostic tools and therapeutic strategies. Science 2018, 359, 1366–1370. [Google Scholar] [CrossRef] [Green Version]
- Puzanov, I.; Diab, A.; Abdallah, K.; Bingham, C.O.; Brogdon, C.; Dadu, R.; Hamad, L.; Kim, S.; Lacouture, M.E.; LeBoeuf, N.R.; et al. Managing toxicities associated with immune checkpoint inhibitors: Consensus recommendations from the Socie-ty for Immunotherapy of Cancer (SITC) Toxicity Management Working Group. J. Immunother. Cancer 2017, 5, 1–28. [Google Scholar] [CrossRef] [Green Version]
- De La Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin is associated with higher relative abundance of mucin-degrading Akkermansia muciniphila and several short-chain fatty acid–producing microbiota in the gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, S.; Calin, G.A. Gut microbiota: A new player in regulating immune- and chemo-therapy efficacy. Cancer Drug Resist. 2020, 3, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- York, A. Gut microbiota sways response to cancer immunotherapy. Nat. Rev. Genet. 2018, 16, 121. [Google Scholar] [CrossRef] [PubMed]
- Cooke, K.R.; Olkiewicz, K.; Erickson, N.; Ferrara, J.L. The role of endotoxin and the innate immune response in the patho-physiology of acute graft versus host disease. J. Endotoxin Res. 2002, 8, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Penack, O.; Holler, E.; van den Brink, M.R. Graft-versus-host disease: Regulation by microbe-associated molecules and in-nate immune receptors. Blood J. Am. Soc. Hematol. 2010, 115, 1865–1872. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, S.; Erturk-Hasdemir, D.; Ochoa-Reparaz, J.; Reinecker, H.-C.; Kasper, D.L. Plasmacytoid Dendritic Cells Mediate Anti-inflammatory Responses to a Gut Commensal Molecule via Both Innate and Adaptive Mechanisms. Cell Host Microbe 2014, 15, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Stingele, F.; Corthésy, B.; Kusy, N.; Porcelli, S.A.; Kasper, D.L.; Tzianabos, A.O. Zwitterionic Polysaccharides Stimulate T Cells with No Preferential Vβ Usage and Promote Anergy, Resulting in Protection against Experimental Abscess Formation. J. Immunol. 2004, 172, 1483–1490. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.-Y.; Ning, M.-X.; Chen, D.-K.; Ma, W.-T. Interactions Between the Gut Microbiota and the Host Innate Immune Response Against Pathogens. Front. Immunol. 2019, 10, 607. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.M.; Jesus, E.C.; Lopes, A.; Aguiar, S.J.; Begnami, M.D.; Rocha, R.M.; Carpinetti, P.A.; Camargo, A.A.; Hoffmann, C.; Freitas, H.C.; et al. Tissue-Associated Bacterial Alterations in Rectal Carcinoma Patients Revealed by 16S rRNA Com-munity Profiling. Front. Cell. Infect. Microbiol. 2016, 6, 179. [Google Scholar] [CrossRef] [Green Version]
- Colosimo, D.A.; Kohn, J.A.; Luo, P.M.; Piscotta, F.J.; Han, S.M.; Pickard, A.J.; Rao, A.; Cross, J.R.; Cohen, L.J.; Brady, S.F. Mapping Interactions of Microbial Metabolites with Human G-Protein-Coupled Receptors. Cell Host Microbe 2019, 26, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Viaud, S.; Daillere, R.; Boneca, I.G.; Lepage, P.; Langella, P.; Chamaillard, M.; Pittet, M.J.; Ghiringhelli, F.; Trinchieri, G.; Goldszmid, R.; et al. Gut microbiome and anticancer immune response: Really hot Sh*t! Cell Death Differ. 2014, 22, 199–214. [Google Scholar] [CrossRef] [Green Version]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daillère, R.; Vétizou, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-Colame, V.; Duong, C.P.; Flament, C.; Lepage, P.; Roberti, M.P.; et al. Enterococcus hirae and Barnesiellaintestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, K.; Baptista, A.P.; Tamoutounour, S.; Zhuang, L.; Bouladoux, N.; Martins, A.J.; Huang, Y.; Gerner, M.Y.; Belkaid, Y.; Germain, R.N. Innate and adaptive lymphocytes sequentially shape the gut microbiota and lipid metabolism. Nat. Cell Biol. 2018, 554, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; Vétizou, M.; Waldschmitt, N.; Kroemer, G.; Chamaillard, M.; Boneca, I.G.; Zitvogel, L. Fine-tuning cancer immu-notherapy: Optimizing the gut microbiome. Cancer Res. 2016, 76, 4602–4607. [Google Scholar]
- Miller, P.L.; Carson, T.L. Mechanisms and microbial influences on CTLA-4 and PD-1-based immunotherapy in the treatment of cancer: A narrative review. Gut Pathog. 2020, 12, 1–10. [Google Scholar] [CrossRef]
- Schönrich, G.; Raftery, M.J. The PD-1/PD-L1 Axis and Virus Infections: A Delicate Balance. Front. Cell. Infect. Microbiol. 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.T.; Lee, S.H.; Heo, Y.-S. Molecular Interactions of Antibody Drugs Targeting PD-1, PD-L1, and CTLA-4 in Immuno-Oncology. Molecules 2019, 24, 1190. [Google Scholar] [CrossRef] [Green Version]
- Harris, E. Immunotherapeutics for the treatment of prostate cancer: A patent landscape based on key therapeutic mechanisms of actions. Pharm. Pat. Anal. 2018, 7, 47–57. [Google Scholar] [CrossRef]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Mager, L.F.; Burkhard, R.; Cooke, N.C.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; Lewis, I.A.; et al. Immunotherapy efficacy in colorectal cancer is dependent on activation of a microbial-metabolite-immune circuit. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Dredge, K.; Marriott, B.J.; Todryk, S.M.; Dalgleish, A.G. Adjuvants and the promotion of Th1-type cytokines in tumour immunotherapy. Cancer Immunol. Immunother. 2002, 51, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Jiao, D.; Qin, S.; Chu, Q.; Li, A.; Wu, K. Manipulating Gut Microbiota Composition to Enhance the Therapeutic Effect of Cancer Immunotherapy. Integr Cancer Ther. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oku, S.; Takeshita, T.; Futatsuki, T.; Kageyama, S.; Asakawa, M.; Mori, Y.; Miyamoto, T.; Hata, J.; Ninomiya, T.; Kashiwazaki, H.; et al. Disrupted tongue microbiota and detection of nonindigenous bacteria on the day of allogeneic hematopoietic stem cell transplantation. PLoS Pathog. 2020, 16, e1008348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, H.K.; Elhaddad, A.M.; Fahmy, O.A.; Samra, M.A.; Abdelfattah, R.M.; El-Nahass, Y.H.; Fathy, G.M.; Abdelhady, M.S. Allogeneic hematopoietic stem cell transplantation for non-malignant hematological disorders. J. Adv. Res. 2015, 6, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fan, Z.-P.; Wu, M.-Q.; Sun, J.; Wu, X.-L.; Xu, D.; Jiang, Q.-L.; Zhang, Y.; Huang, F.; Wei, Y.-Q.; et al. Allo-HSCT for acute leukemia of ambiguous lineage in adults: The comparison between standard conditioning and intensified conditioning regimens. Ann. Hematol. 2012, 92, 679–687. [Google Scholar] [CrossRef]
- Taur, Y.; Xavier, J.B.; Lipuma, L.; Ubeda, C.; Goldberg, J.; Gobourne, A.; Lee, Y.J.; Dubin, K.A.; Socci, N.D.; Viale, A.; et al. Intestinal Domination and the Risk of Bacteremia in Patients Undergoing Allogeneic Hematopoietic Stem Cell Transplantation. Clin. Infect. Dis. 2012, 55, 905–914. [Google Scholar] [CrossRef]
- Li, L. Infectious Microecology in Liver Disease. In Infectious Microecology; Springer: Berlin/Heidelberg, Germany, 2014; pp. 317–332. [Google Scholar]
- Ashraf, R.; Shah, N.P. Selective and differential enumerations of Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus, Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium spp. in yoghurt—A review. Int. J. Food Microbiol. 2011, 149, 194–208. [Google Scholar] [CrossRef]
- Eslami-S, Z.; Majidzadeh-A, K.; Halvaei, S.; Babapirali, F.; Esmaeili, R. Microbiome and Breast Cancer: New Role for an Ancient Population. Front. Oncol. 2020, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Tharmaraj, N.; Shah, N.P. Survival of Lactobacillus acidophilus, Lactobacillus paracasei subsp. paracasei, Lactobacillus rhamnosus, Bifidobacterium animalis and Propionibacterium in cheese-based dips and the suitability of dips as effective carriers of probiotic bacteria. Int. Dairyj. 2004, 14, 1055–1066. [Google Scholar] [CrossRef]
- Ambalam, P.; Raman, M.; Purama, R.K.; Doble, M. Probiotics, prebiotics and colorectal cancer prevention. Best Pr. Res. Clin. Gastroenterol. 2016, 30, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Cai, X.; Zhang, J.; Wang, W.; Sheng, Q.; Hua, H.; Zhou, X. Role of Gut Microbiota in the Development and Treatment of Colorectal Cancer. Digestion 2018, 100, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar] [CrossRef]
- Jones, S.E.; Versalovic, J. Probiotic Lactobacillus reuteri biofilms produce antimicrobial and anti-inflammatory factors. BMC Microbiol. 2009, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Duran-Pinedo, A.E.; Frias-Lopez, J. Beyond microbial community composition: Functional activities of the oral microbiome in health and disease. Microbes Infect. 2015, 17, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Jahrsdörfer, B.; Weiner, G.J. CpG oligodeoxynucleotides as immunotherapy in cancer. Updat. Cancer Ther. 2008, 3, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Aliberti, J.; Hoffmann, K.F.; Jankovic, D.; Feng, C.G.; Kullberg, M.C.; Sher, A. Experimental models for the analysis of IL-10 function. In Interleukin-10; Marincola, F.M., Ed.; Taylor & Francis: Abingdon, UK, 2006. [Google Scholar] [CrossRef]
- Donnelly, R.P.; Dickensheets, H.; Finbloom, D.S. The Interleukin-10 Signal Transduction Pathway and Regulation of Gene Expression in Mononuclear Phagocytes. J. Interf. Cytokine Res. 1999, 19, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Ni, G.; Wang, T.; Walton, S.F.; Zhu, B.; Chen, S.; Wu, X.; Wang, Y.; Wei, M.Q.; Liu, X.S. Manipulating IL-10 signalling blockade for better immunotherapy. Cell. Immunol. 2015, 293, 126–129. [Google Scholar] [CrossRef]
- Rigopoulou, E.I.; Abbott, W.G.; Haigh, P.; Naoumov, N.V. Blocking of interleukin-10 receptor—A novel approach to stimulate T-helper cell type 1 responses to hepatitis C virus. Clin. Immunol. 2005, 117, 57–64. [Google Scholar] [CrossRef]
- Adamus, T.; Kortylewski, M. The revival of CpG oligonucleotide-based cancer immunotherapies. Współczesna Onkol. 2018, 2018, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Zhang, Z. The application of nanotechnology in immune checkpoint blockade for cancer treatment. J. Control. Release 2018, 290, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Brandi, G.; Frega, G. Microbiota: Overview and Implication in Immunotherapy-Based Cancer Treatments. Int. J. Mol. Sci. 2019, 20, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- June, C.H.; O’Connor, R.S.; Kawalekar, O.U.; Ghassemi, S.; Milone, M.C. CAR T cell immunotherapy for human cancer. Science 2018, 359, 1361–1365. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, M.H.; Westwood, J.A.; Slaney, C.Y.; Darcy, P.K. Clinical application of genetically modified T cells in cancer therapy. Clin. Transl. Immunol. 2014, 3, e16. [Google Scholar] [CrossRef]
- Shui, L.; Yang, X.; Li, J.; Yi, C.; Sun, Q.; Zhu, H. Gut microbiome as a potential factor for modulating resistance to cancer immunotherapy. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Kump, P.; Wurm, P.; Gröchenig, H.P.; Wenzl, H.; Petritsch, W.; Halwachs, B.; Wagner, M.; Stadlbauer, V.; Eherer, A.; Hoffmann, K.M.; et al. The taxonomic composition of the donor intestinal microbiota is a major factor influencing the efficacy of faecal microbiota transplantation in therapy refractory ulcerative colitis. Aliment. Pharmacol. Ther. 2018, 47, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Ooijevaar, R.; Van Beurden, Y.; Terveer, E.; Goorhuis, A.; Bauer, M.; Keller, J.; Mulder, C.; Kuijper, E. Update of treatment algorithms for Clostridium difficile infection. Clin. Microbiol. Infect. 2018, 24, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Perez, E.; Lee, C.H.; Petrof, E.O. A Practical Method for Preparation of Fecal Microbiota Transplantation. Adv. Struct. Saf. Stud. 2016, 9, 259–267. [Google Scholar] [CrossRef]
- Zuo, T.; Wong, S.H.; Lam, K.; Lui, R.; Cheung, K.; Tang, W.; Ching, J.Y.L.; Chan, P.K.S.; Chan, M.C.-W.; Wu, J.C.Y.; et al. Bacteriophage transfer during faecal microbiota transplantation inClostridiumdifficileinfection is associated with treatment outcome. Gut 2017, 67, 634–643. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Andoh, A.; Sugimoto, M. Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn’s Disease. Digestion 2016, 93, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Spohn, S.N.; Young, V.B. Gastrointestinal Microbial Ecology with Perspectives on Health and Disease. In Physiology of the Gastrointestinal Tract; Elsevier: Amsterdam, The Netherlands, 2018; pp. 737–753. [Google Scholar]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Neuman, H.; Debelius, J.W.; Knight, R.; Koren, O. Microbial endocrinology: The interplay between the microbiota and the endocrine system. FEMS Microbiol. Rev. 2015, 39, 509–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandrini, S.; Aldriwesh, M.; Alruways, M.; Freestone, P. Microbial endocrinology: Host-bacteria communication within the gut microbiome. J. Endocrinol. 2015, 225, R21–R34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceranowicz, P.; Warzecha, Z.; Dembinski, A. Peptidyl hormones of endocrine cells origin in the gut—Their discovery and physiological relevance. J. Physiol. Pharmacol. 2015, 66, 11–27. [Google Scholar] [PubMed]
- Noureldein, M.H.; Eid, A.A. Gut microbiota and mTOR signaling: Insight on a new pathophysiological interaction. Microb. Pathog. 2018, 118, 98–104. [Google Scholar] [CrossRef]
- Kar, S.K.; Jansman, A.J.; Benis, N.; Ramiro-Garcia, J.; Schokker, D.; Kruijt, L.; Stolte, E.H.; Taverne-Thiele, J.J.; Smits, M.A.; Wells, J.M. Dietary protein sources differentially affect microbiota, mTOR activity and transcription of mTOR signaling pathways in the small intestine. PLoS ONE 2017, 12, e0188282. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Jazwinski, S.M. The gut microbiota and healthy aging: A mini-review. Gerontology 2018, 64, 513–520. [Google Scholar] [CrossRef]
- Yeh, C.F.; Chen, Y.H.; Liu, S.F.; Kao, H.L.; Wu, M.S.; Yang, K.C.; Wu, W.K. Mutual Interplay of Host Immune System and Gut Microbiota in the Immunopathology of Atherosclerosis. Int. J. Mol. Sci. 2020, 21, 8729. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.J.; Huffnagle, G.B. The microbiome and regulation of mucosal immunity. Immunology 2014, 142, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Wade, P.A. Crosstalk between the microbiome and epigenome: Messages from bugs. J. Biochem. 2018, 163, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostaff, M.J.; Stange, E.F.; Wehkamp, J. Antimicrobial peptides and gut microbiota in homeostasis and pathology. EMBO Mol. Med. 2013, 5, 1465–1483. [Google Scholar] [CrossRef]
- Jan, G.; Belzacq, A.S.; Haouzi, D.; Rouault, A.; Métivier, D.; Kroemer, G.; Brenner, C. Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 2002, 9, 179–188. [Google Scholar] [CrossRef]
- Wei, W.; Sun, W.; Yu, S.; Yang, Y.; Ai, L. Butyrate production from high-fiber diet protects against lymphoma tumor. Leuk. Lymphoma 2016, 57, 2401–2408. [Google Scholar] [CrossRef]
- Paavonen, J.; Naud, P.; Salmerón, J.; Wheeler, C.M.; Chow, S.N.; Apter, D.; Kitchener, H.; Castellsague, X.; Teixeira, J.C.; Skinner, S.R.; et al. Efficacy of human papillomavirus (HPV)-16/18 AS04-adjuvanted vaccine against cervical infection and precancer caused by oncogenic HPV types (PATRICIA): Final analysis of a double-blind, randomised study in young women. Lancet 2009, 374, 301–314. [Google Scholar] [CrossRef]
- Aranda, F.; Bloy, N.; Pesquet, J.; Petit, B.; Chaba, K.; Sauvat, A.; Kepp, O.; Khadra, N.; Enot, D.; Pfirschke, C.; et al. Immune-dependent antineoplastic effects of cisplatin plus pyridoxine in non-small-cell lung cancer. Oncogene 2015, 34, 3053–3062. [Google Scholar] [CrossRef]
- Konishi, H.; Fujiya, M.; Tanaka, H.; Ueno, N.; Moriichi, K.; Sasajima, J.; Ikuta, K.; Akutsu, H.; Tanabe, H.; Kohgo, Y. Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis. Nat. Commun. 2016, 7, 12365. [Google Scholar] [CrossRef]
- Lenoir, M.; Del Carmen, S.; Cortes-Perez, N.G.; Lozano-Ojalvo, D.; Muñoz-Provencio, D.; Chain, F.; Langella, P.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Lactobacillus casei BL23 regulates Treg and Th17 T-cell populations and reduces DMH-associated colorectal cancer. J. Gastroenterol. 2016, 51, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Shin, J.G.; Kim, E.H.; Kang, H.E.; Yim, I.B.; Kim, J.Y.; Joo, H.G.; Woo, H.J. Immunomodulatory and antitumor effects in vivo by the cytoplasmic fraction of Lactobacillus casei and Bifidobacterium longum. J. Vet. Sci. 2004, 5, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, C.; Millette, M.; Oth, D.; Ruiz, M.T.; Luquet, F.M.; Lacroix, M. Probiotic Lactobacillus acidophilus and L. casei mix sensitize colorectal tumoral cells to 5-fluorouracil-induced apoptosis. Nutr. Cancer 2010, 62, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Takagi, A.; Ikemura, H.; Matsuzaki, T.; Sato, M.; Nomoto, K.; Morotomi, M.; Yokokura, T. Relationship between the in vitro response of dendritic cells to Lactobacillus and prevention of tumorigenesis in the mouse. J. Gastroenterol. 2008, 43, 661–669. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, E.F. The toxins of William, B. Coley and the treatment of bone and soft-tissue sarcomas. IOWA Orthop. J. 2006, 26, 154–158. [Google Scholar] [PubMed]
- Nauts, H.C.; Swift, W.E.; Coley, B.L. The treatment of malignant tumors by bacterial toxins as developed by the late William, B.; Coley, M.D., reviewed in the light of modern research. Cancer Res. 1946, 6, 205–216. [Google Scholar]
- Zbar, B.; Bernstein, I.; Tanaka, T.; Rapp, H.J. Tumor immunity produced by the intradermal inoculation of living tumor cells and living Mycobacterium bovis (strain BCG). Science 1970, 170, 1217–1218. [Google Scholar] [CrossRef]
- Aso, Y.; Akazan, H. Prophylactic effect of a Lactobacillus casei preparation on the recurrence of superficial bladder cancer. BLP Study Group. Urol. Int. 1992, 49, 125–129. [Google Scholar] [CrossRef]
- Hoesl, C.E.; Altwein, J.E. The probiotic approach: An alternative treatment option in urology. Eur. Urol. 2005, 47, 288–296. [Google Scholar] [CrossRef]
- Stebbing, J.; Dalgleish, A.; Gifford-Moore, A.; Martin, A.; Gleeson, C.; Wilson, G.; Brunet, L.R.; Grange, J.; Mudan, S. An intra-patient placebo-controlled phase I trial to evaluate the safety and tolerability of intradermal IMM-101 in melanoma. Ann. Oncol. 2012, 23, 1314–1319. [Google Scholar] [CrossRef]
- Dalgleish, A.G.; Stebbing, J.; Adamson, D.J.; Arif, S.S.; Bidoli, P.; Chang, D.; Cheeseman, S.; Diaz-Beveridge, R.; Fernandez-Martos, C.; Glynne-Jones, R.; et al. Randomised, open-label, phase II study of gemcitabine with and without IMM-101 for advanced pancreatic cancer. Br. J. Cancer. 2016, 115, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F.; et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Nemunaitis, J.; Cunningham, C.; Senzer, N.; Kuhn, J.; Cramm, J.; Litz, C.; Cavagnolo, R.; Cahill, A.; Clairmont, C.; Sznol, M. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene Ther. 2003, 10, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.G.; Masner, M.; Ferreira, F.A.; Hoffman, R.M. Bacterial Therapy of Cancer: Promises, Limitations, and Insights for Future Directions. Front. Microbiol. 2018, 9, 16. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Cancer | Sampling Location | Microbial Increase/Decrease | References |
|---|---|---|---|
| Colorectal | Biopsied tissue and feces materials | Increased: Escherichia coli, Staphylococcus bovis, Fusobacterium nucleatum, Clostridium spp., Streptococcus spp., and Bacteroides. Decreased: Butyrate-producing bacteria, Lactobacillus, Microbacterium, Anoxybacillus, and Akkermansia muciniphila. | [26,27,28] |
| Gall bladder | Bile samples | Increased: Salmonella paratyphi and S. typhi; Bile is typically considered to be bacteria-free but is infected in many cases. | [28,29] |
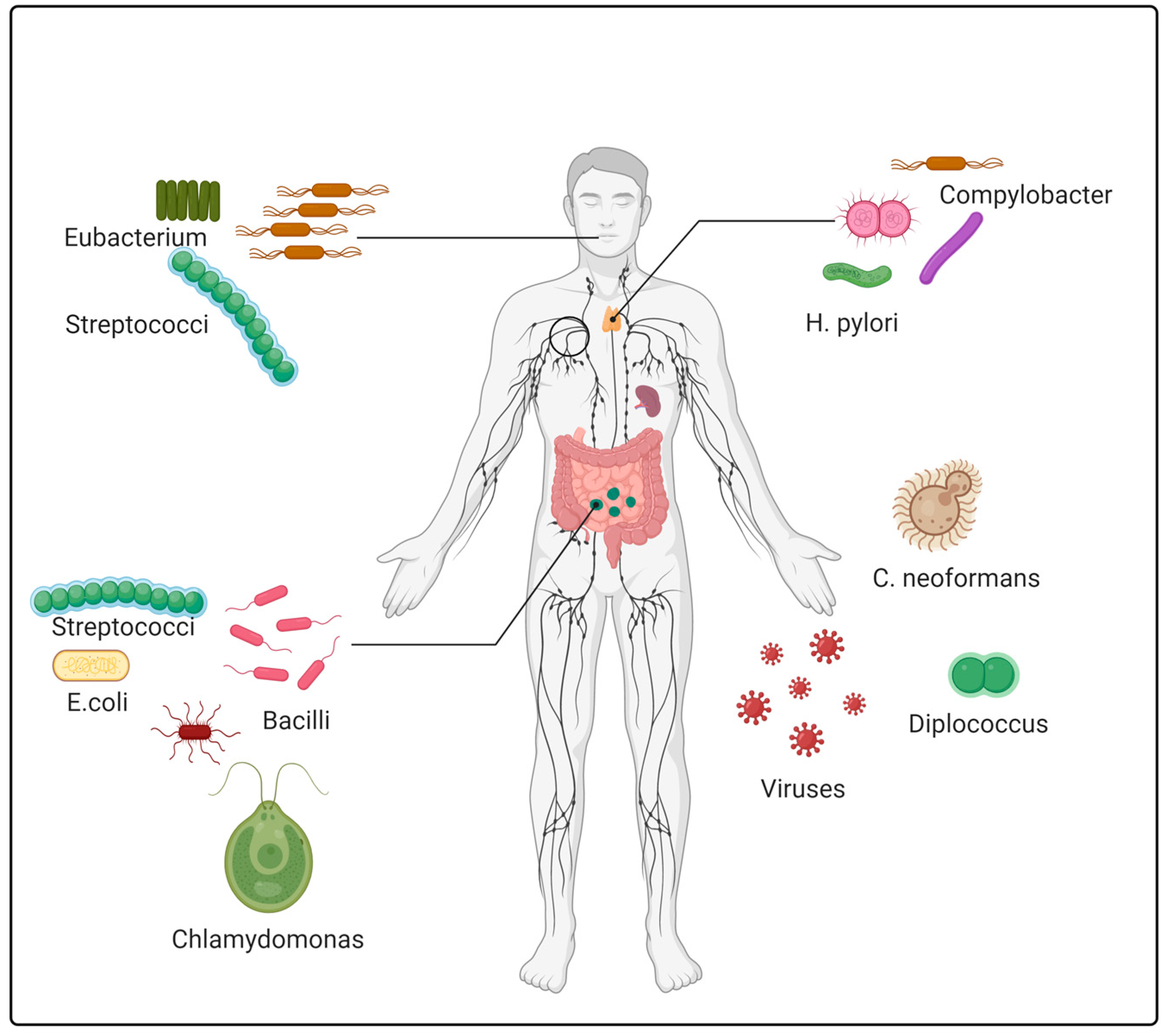
| Esophageal and Barrett’s esophagus | Saliva sample and biopsy tissue | Increased: S. anginosus, Treponema denticola, Campylobacter concisus, C. rectus and S. mitis. Decreased: Helicobacter pylori. | [30,31,32] |
| Mouth carcinoma | Saliva culture | Increased: Eubacterium sabureum, Leptotrichia buccalis, C. ochracea, Capnocytophaga gingivalis and Streptococcus mitis | [33,34,35] |
| Microbes | Type(s) of Cancer |
|---|---|
| Human herpes virus 8 | Kaposi’ssarcoma |
| Type 1 human T-cell lymphotropic virus (HTLV-1) | T-cell lymphoma, leukemia (adult) |
| Hb-B virus | Hepatocellular carcinoma |
| Hb-Cvirus | Lymphoma, hepatocellular carcinoma |
| HIV (human immunodeficiency virus) | Kaposi’s sarcoma, lymphomas |
| EBV (Epstein–Barr virus) | Nasopharyngeal carcinoma, lymphomas |
| HPV (human papilloma virus) | Oropharyngeal carcinoma, anogenital carcinomas |
| Helicobacter pylori | Esophageal adenocarcinoma, gastric lymphoma, gastric adenocarcinoma |
| Cancer | Study Model | Outcome/s | References |
|---|---|---|---|
| Murine Studies | |||
| Breast | Germ-free rats treated with DMAB. | Reduced tumors growth in germ-free rats. | [50] |
| Lung | Germ-free rats treated with NHMI. | Fewer tumors in male germ-free rats. No observed changes in female germ-free rats. | [51] |
| Gastric | 1. INS-GAS mice (gnotobiotic) infected with Helicobacter pylori. | Fewer tumors in germ-free mice. | [52] |
| 2. Antibiotic-treated INS-GAS mice infected with Helicobacter pylori. | Fewer tumors in mice treated with antibiotics. | [53] | |
| Liver | 1. Germ-free mice treated with (DEN) and CCl4. | Fewer tumors in germ-free mice. | [53] |
| 2. An antibiotic cocktail was administered to DEN and CCl4-treated mice. | Fewer tumors in antibiotic-treated mice. | [54] | |
| 3. Rifaximin administered to DEN and CCl4-treated mice. | Fewer tumors in rifaximin-treated mice. | [54] | |
| 4. Neomycin administeredto DEN-treated rats. | Fewer tumors in neomycin-treated rats. | [54] | |
| 5. Vancomycin administeredto DMBA-treated miceon a high-fatdiet. | Fewer tumors in vancomycin-treated mice. | [55] | |
| 6. An antibiotic cocktail administered to DMBA-treated mice on a high-fat diet. | Fewer tumors in antibiotic-treated mice. | [55] | |
| Colorectal | 1. Germ-free mice (ApcMin/+). | Fewer tumors in germ-free mice. | [56] |
| 2. Gnotobiotic mice (AOM in IL-10−/−). | Fewer tumors in germ-free mice. | [57] | |
| 3. Mice (ApcMin/+ Cdx2-Cre) treated with an antibiotic mixture. | Fewer tumors in antibiotic-treated mice. | [58] | |
| 4. Mice (Nod1−/−) treated with an antibiotic mixture. | Fewer tumors in antibiotic-treated mice. | [59] | |
| 5. Mice (DSS and AOM) treated with an antibiotic mixture. | Fewer tumors in antibiotic-treated mice. | [60] | |
| 6. Mice (Nod2−/−) were transplanted with wild-type microbiota. | Fewer tumors after transplant. | [60] | |
| Type of Cancer | Role of Microbes/Mechanism of Carcinogenesis | Evidence/Proof | References |
|---|---|---|---|
| Gastric lymphoma of the gastric MALT, IPSID, MALT lymphoma of the skin, adnexal ocular lymphoma | Chronic infection with Helicobacter pylori Patients with chronic infection with H. pylori, Campylobacter jejuni, Borreliaburgdorferi, or Chlamydia psittaci | Epidemiology support Reduction by H. pylori Eradication Antibiotic treatment | [63,64,65,66,67,68,69,70,71,72,73] |
| Esophageal | Decreased risk in patients who have H. pylori infection | Epidemiology support | [70,74] |
| Gallbladder | Chronic infection with Salmonella enteric subsp. enterica serovar Typhi | Epidemiology support | [75,76] |
| Breast | Increased T regulatory cell-mediated inflammation | Cancer promoted in ApcMin/+ mice infected with Helicobacter hepaticus | [77] |
| Liver | Chronic hepatitis | Cancer facilitated in mice infected with H. hepaticus | [78] |
| Colorectal | TNF-mediated and NO-mediated | Cancer supported in Rag2−/− mice infected with H. hepaticus | [79] |
| Colorectal |
| Cancer reduction by antibiotics and in germ-free mice; transmission of dysbiotic microbiota triggers cancer development | [80,81,82,83,84] |
| Liver |
| Cancer reduction by treatment with antibiotics and in germ-free mice Cancer increased by treatment with LPS and DCA | [85,86] |
| Lung | Increased bacterial infection in COPD | Decreased cancer in germ-free animals The promotion of cancer by LPS and infections | [87,88,89,90,91] |
| Pancreatic | LPS-TLR4-mediated increase | LPS treatment increases cancer development | [92,93,94] |
| Global NCT Number | Type of Malignancy | Aim | Intervention | Results/Outcome Measures | Place |
|---|---|---|---|---|---|
| 03290651 | Breast cancer | To determine if oral antibiotics can change the breast flora | Probiotics Natural Health Product-RepHresh Pro-B | Change in breast microbiota, inflammatory markers | Canada |
| 03341143 | Melanoma | To establish concurrent use of FMT and pembrolizumab in patients with PD-1-resistant melanoma | FMT (donor responder to PD-1 therapy) with pembrolizumab | ORR, change in T cell composition and function, change in innate and adaptive immune subsets | USA |
| 00936572 | Colorectal cancer | To investigate the effect of probiotics on gut microflora and the immune and inflammatory response | Probiotics (La1, BB536) | To perform morphological and microbiological evaluation of the colonic microflora, GI function | Italy |
| 03072641 | Colorectal cancer | To reactivate the tumor-suppressor genes using probiotics | ProBionClinica (Bifidobacteriumlactis, L. acidophilus) | Changes in microbiota composition and DNA methylation | Sweden |
| 01609660 | Colorectal cancer | To assess the impact of probiotics on patients undergoing colorectal resections | Saccharomyces boulardii | To measure mucosal cytokines, SCFA postoperative complications, and hospital LOS | Brazil |
| 00197873 | Colorectal cancer | To prevent chemotherapy-induced diarrhea | L. rhamnosus supplementation | Effect on treatment-related toxicity other than diarrhea | Finland |
| 02269150 | Malignancies requiring allo-HSCT | To assess the utility of FMT in prevention of CDI in patients who underwent allo-HSCT | Auto-FMT | CDI | USA |
| 02928523 | Acute myeloid leukemia | To use FMT to prevent complications associated with dysbiosis in patients undergoing intensive treatment | Auto-FMT | Dysbiosis correction, eradication of multidrug resistant bacteria, definition of dysbiosis, biosignature | France |
| 03552458 | Head-and-neck cancer | To assess the role of probiotics in preventing oral mucositis | Lactobacillus reuteri Oral Solution (BioGaia) | Oral mucositis severity, oral bacterial genetics, and transcriptional analysis | Singapore |
| Accession or Trial Number | Intervention | Targeted Patient Population | Finding(s) |
|---|---|---|---|
| NCT03072641 | Irregular probiotics supplements (ProBionClinicaB. lactis BI-04, L. acidophilus Inulin+ NCFM) Intake | CRC patients ages18+ | Primary: change in fecal and tumor microbiota. Secondary: Changes in epigenetics patterns of tumor tissue. |
| NCT01895530 | Randomized probiotic (S. Boulardii) administration | CRC patients ages 18+ undergoing elective CRC resection | Primary: cytokine expression in colonic mucosa (via qPCR). Secondary: post-operative complications. |
| NCT03358511 | Single-arm probiotics (Primal Defense Ultra multi-strain probiotics formula) | Post-menopausal breast cancer patients (stages I–III) | Primary: change in mean number of CD8+ cells. |
| NCT03353402 | Single-arm FMT (colonoscopy or gastroscopy) from patient donors who responded to immunotherapy | Metastatic melanoma patients ages 18+ who previously failed standard therapies | Primary: safety (AEs associated with FMT), engraftment of FMT. Secondary: changes in immune cell populations and activity, objective response rate. |
| NCT02928523 | Single-arm autologous FMT (frozen inoculum) | Acute myeloid leukemia patients ages 18–65 treated with intensive chemotherapy and antibiotics | Primary: diversity of the gut microbiome, multi-drug-resistant bacteria eradication. Secondary: signature of dysbiosis. of the gut microbiome. |
| NCT02079662 | Randomized intensive lifestyle change (diet, exercise, psychosocial) | Stages II and III breast cancer patients treated at MDACC ages 18+ | Primary: disease-free survival (DFS). Secondary: change in fecal and oral microbiome (via 16S profiling). |
| NCT02843425 | Addition of ½ cup beans per day to regular diet in a crossover design | All cancer patients treated at MDACC | Primary: change in fecal microbiome profile. |
| Main Outcome | Data Source | Carcinoma/Treatment | References |
|---|---|---|---|
| Immunological therapy Commensal type of microbiota needed for standard response to treatment | Rodent—mouse | Several models of carcinoma/anti-IL-10R antibody + CpG oligonucleotide and oxaliplatin (platinum-chemotherapy) | [161] |
| Irradiation of whole body with disruption of the intestine barrier and enhanced response of T-lymphocyte cell-mediated treatment via mechanisms dependent upon TLR4 signaling/microbe translocation/LPS | Rodent—mouse | Melanoma/Adoptive T-lymphocyte transfer of cells | [162] |
| Firmicutes and Faecalibacterium presence in baseline samples of stool liked with ICB response; abundance of Bacteroides linked with low ICB response | Human | Melanoma (Metastatic stage)/CTLA-4 inhibitor | [163] |
| Abundance of Blautia linked with overall increased survival rate and decreased GVHD risk | Human | Hematologic cancers/Allo-HSCT | [164] |
| Bacteroides abundance was associated with resistance to ICB-induced colitis | Human | Melanoma (Metastatic stage)/CTLA-4 inhibitor | [165] |
| Abundance of Bifidobacterium linked with enhanced automatic immunity against tumor and ICB response | Rodent—mouse | Melanoma/Anti-PD-L1 inhibitor | [166] |
| Abundance of Bacteroides liked with ICB response | Rodent—human | Melanoma (Metastatic stage)/CTLA-4 inhibitor | [167] |
| Abundance of Eubacterium limosum linked with reduced chances of progression of disease or relapse | Rodent—human | Blood cancer/Allo-HSCT | [168] |
| Faecalibacterium prausnitzii, Bacteroides caccae, Holdemaniafiliformis, Bacteroides thetaiotaomicron, and Doreaformicigenerans linked with ICB response | Human | Melanoma (Metastatic stage)/CTLA-4 inhibitor; PD-1 inhibitor | [169] |
| Large number of bacteria in the sample of baseline stool found to be enriched differentially between strong ICB response patients vs. poor ICB response patients | Human | Mouse-melanoma (Metastatic)/PD-1 inhibitor | [170] |
| Clostridiales, high microbiome richness and abundance of Faecalibacterium, Ruminococcaceae with ICB response in baseline stool samples | Mouse; Human | Melanoma (Metastatic stage)/PD-1 inhibitor | [171] |
| Abundance of A. muciniphila in the samples of baseline stool found to be linked with ICB response | Mouse; Human | lung carcinoma of non-small cell; Renal cancer/PD-1 inhibitor | [172] |
| Chemotherapy with immunostimulatory properties Existence of intratumoral Gammaproteobacteria linked with gemcitabine chemotherapy resistance | Mouse; Human | Adenocarcinoma (Ductal Pancreatic)/Gemcitabine immunomodulatory chemotherapy treatment | [173] |
| Abundance of Akkermansia muciniphila in the samples of baseline stool found to be liked with ICB response | Rodent-Mouse | Several models of carcinoma/immunomodulatory chemotherapy Cyclophosphamide | [174] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhry, H. The Microbiome and Its Implications in Cancer Immunotherapy. Molecules 2021, 26, 206. https://doi.org/10.3390/molecules26010206
Choudhry H. The Microbiome and Its Implications in Cancer Immunotherapy. Molecules. 2021; 26(1):206. https://doi.org/10.3390/molecules26010206
Chicago/Turabian StyleChoudhry, Hani. 2021. "The Microbiome and Its Implications in Cancer Immunotherapy" Molecules 26, no. 1: 206. https://doi.org/10.3390/molecules26010206
APA StyleChoudhry, H. (2021). The Microbiome and Its Implications in Cancer Immunotherapy. Molecules, 26(1), 206. https://doi.org/10.3390/molecules26010206