
Neuroprotective Activities of Crossyne flava Bulbs and Amaryllidaceae Alkaloids: Implications for Parkinson’s Disease

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Identification of Compounds
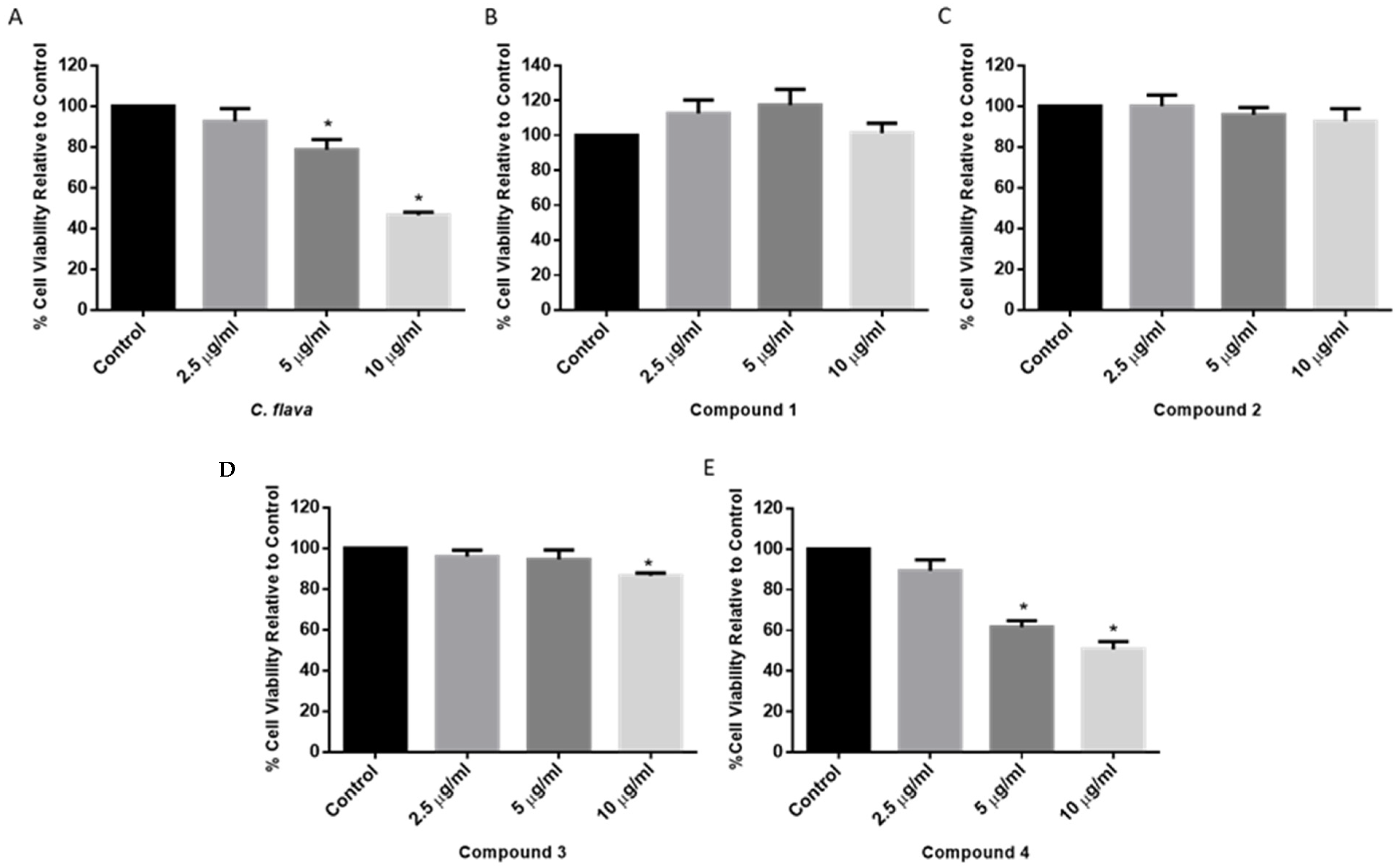
2.2. Dose Response of C. flava and Compounds
2.3. C. flava and Compounds Mitigated MPP+-Induced Toxicity
2.4. C. flava Prevented MPP+-Induced Alterations in Cell Morphology
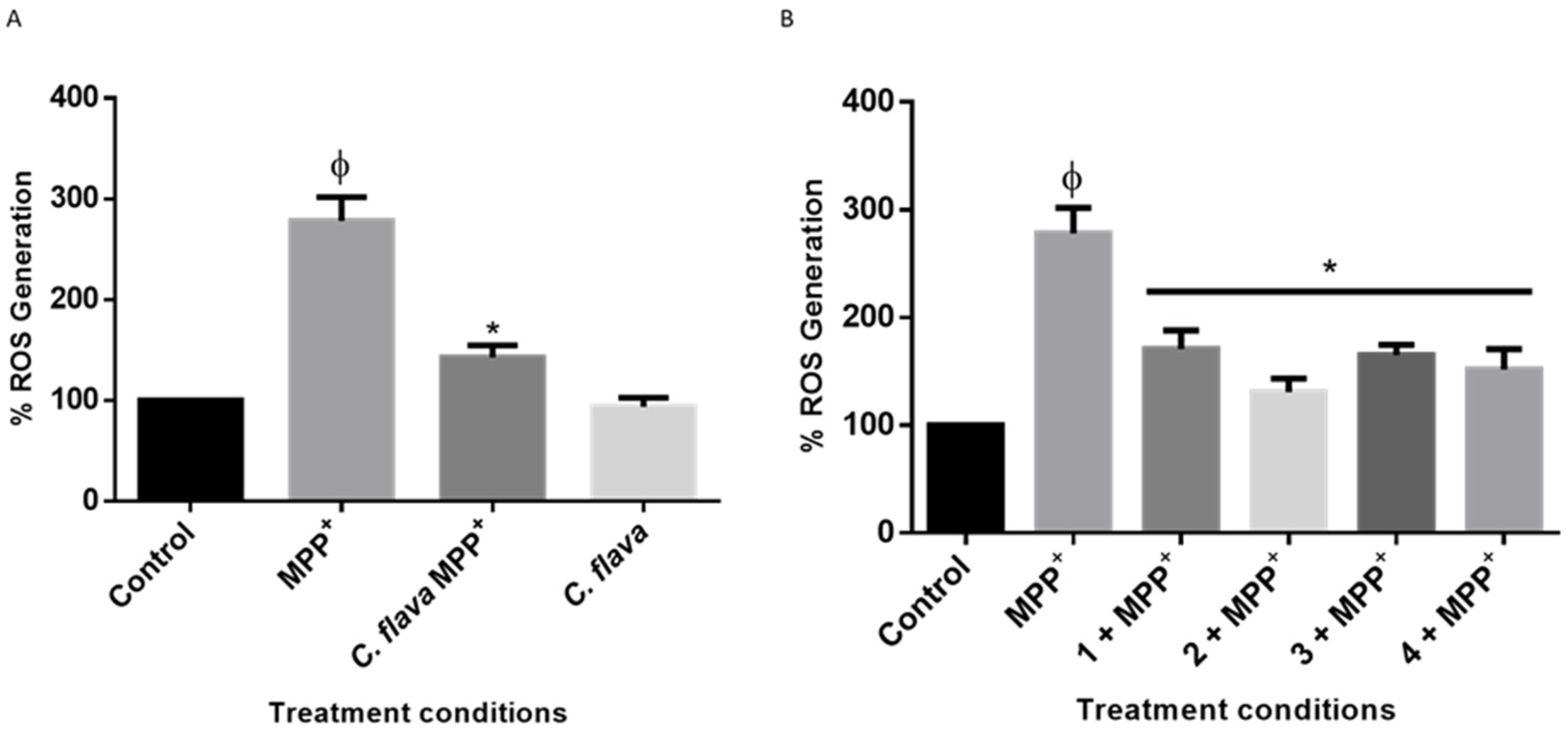
2.5. C. flava and Compounds Mitigate MPP+-Induced ROS Generation
2.6. C. flava and Compounds Attenuated MPP+-Induced Loss of ATP
2.7. C. flava and Compounds Inhibited MPP+-Induced Apoptosis
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Collection and Identification of the Plant Material
4.3. Preparation of Plant Extract and Isolation of Compounds
4.4. Cell Culture and Maintenance
4.5. Treatments
4.6. Cell Viability Assays
4.7. Cell Morphology
4.8. Reactive Oxygen Species (ROS) Assay
4.9. Adenosine Triphosphate Assay
4.10. Caspase 3/7 Apoptosis Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ferreira, S.A.; Romero-Ramos, M. Microglia response during Parkinson’s disease: Alpha-synuclein intervention. Front. Cell. Neurosci. 2018, 12, 247. [Google Scholar] [CrossRef] [Green Version]
- Massano, J.; Bhatia, K.P. Clinical approach to Parkinson’s disease: Features, diagnosis, and principles of management. Cold Spring Harb. Perspect. Med. 2012, 2, a008870. [Google Scholar] [CrossRef]
- Kaidery, N.A.; Thomas, B. Current perspective of mitochondrial biology in Parkinson’s disease. Neurochem. Int. 2018, 117, 91–113. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Gao, J.; Wang, J.; Xie, A. Prion-like mechanisms in Parkinson’s disease. Front. Neurosci. 2019, 13, 552. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Manna, J.; Dunbar, G.L. Current understanding of the molecular mechanisms in Parkinson’s disease: Targets for potential treatments. Transl. Neurodegener. 2017, 6, 28. [Google Scholar]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Parkinson’s Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [Green Version]
- Kieburtz, K.; Wunderle, K.B. Parkinson’s disease: Evidence for environmental risk factors. Mov. Disord. Off. J. Mov. Disord. Soc. 2013, 28, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ritz, B. The search for environmental causes of Parkinson’s disease: Moving forward. J. Parkinson’s Dis. 2018, 8, S9–S17. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.K.; Yankee, E.L. A review on Parkinson’s disease treatment. Neuroimmunol. Neuroinflamm. 2021, 8. [Google Scholar] [CrossRef]
- Zahoor, I.S.A.; Haq, E. Pharmacological Treatment of Parkinson’s Disease. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; Stoker, T.B., Greenland, J.C., Eds.; Codon Publications: Brisbane, Australia, 2018; Chapter 7. [Google Scholar]
- Surguchov, A.; Bernal, L.; Surguchev, A.A. Phytochemicals as Regulators of Genes Involved in Synucleinopathies. Biomolecules 2021, 11, 624. [Google Scholar] [CrossRef]
- Rabiei, Z.; Solati, K.; Amini-Khoei, H. Phytotherapy in treatment of Parkinson’s disease: A review. Pharm. Biol. 2019, 57, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhao, D.; Bezard, E. Traditional Chinese medicine for Parkinson’s disease: A review of Chinese literature. Behav. Pharmacol. 2006, 17, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.E.; Hess, C.W.; Fox, S.H. Motor Complications of Dopaminergic Medications in Parkinson’s Disease. Semin. Neurol. 2017, 37, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Fennell, C.W.; van Staden, J. Crinum species in traditional and modern medicine. J. Ethnopharmacol. 2001, 78, 15–26. [Google Scholar] [CrossRef]
- Koekemoer, M.; Steyn, H.M.; Bester, S.P. Guide to Plant Families of Southern Africa; South African National Biodiversity Institute: Pretoria, South Africa, 2013. [Google Scholar]
- Van Goietsenoven, G.; Andolfi, A.; Lallemand, B.; Cimmino, A.; Lamoral-Theys, D.; Gras, T.; Abou-Donia, A.; Dubois, J.; Lefranc, F.; Mathieu, V.; et al. Amaryllidaceae Alkaloids Belonging to Different Structural Subgroups Display Activity against Apoptosis-Resistant Cancer Cells. J. Nat. Prod. 2010, 73, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qu, C.; Gao, O.; Hu, X.; Hong, X. Biological and pharmacological activities of Amaryllidaceae alkaloids. RSC Adv. 2015, 5, 16562–16574. [Google Scholar] [CrossRef]
- Naidoo, D.; Roy, A.; Slavětínská, L.P.; Chukwujekwu, J.C.; Gupta, S.; Van Staden, J. New role for crinamine as a potent, safe and selective inhibitor of human monoamine oxidase B: In vitro and in silico pharmacology and modeling. J. Ethnopharmacol. 2020, 248, 112305. [Google Scholar] [CrossRef]
- Sibanyoni, M.N.; Chaudhary, S.K.; Chen, W.; Adhami, H.-R.; Combrinck, S.; Maharaj, V.; Schuster, D.; Viljoen, A. Isolation, in vitro evaluation and molecular docking of acetylcholinesterase inhibitors from South African Amaryllidaceae. Fitoterapia 2020, 146, 104650. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Teoh, H.L. Galanthamine from snowdrop—the development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef]
- Viladomat, F.; Bastida, J.; Codina, C.; Campbell, W.E.; Mathee, S. Alkaloids from Boophane flava. Phytochemistry 1995, 40, 307–311. [Google Scholar] [CrossRef]
- Viladomat, F.; Codina, C.; Bastida, J.; Mathee, S.; Campbell, W.E. Further alkaloids from Brunsvigia josephinae. Phytochemistry 1995, 40, 961–965. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, chemotaxonomy and chemoecology of Amaryllidaceae alkaloids. In The Alkaloids: Chemistry and Biology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 83, pp. 113–185. [Google Scholar]
- Cedron, J.C.; Oberti, J.C.; Estevez-Braun, A.; Ravelo, A.G.; Del Arco-Aguilar, M.; Lopez, M. Pancratium canariense as an important source of Amaryllidaceae alkaloids. J. Nat. Prod. 2009, 72, 112–116. [Google Scholar] [CrossRef]
- Liu, W.-B.; Zhou, J.; Qu, Y.; Li, X.; Lu, C.-T.; Xie, K.-L.; Sun, X.-L.; Fei, Z. Neuroprotective effect of osthole on MPP+-induced cytotoxicity in PC12 cells via inhibition of mitochondrial dysfunction and ROS production. Neurochem. Int. 2010, 57, 206–215. [Google Scholar] [CrossRef]
- Abushouk, A.I.; Negida, A.; Ahmed, H.; Abdel-Daim, M.M. Neuroprotective mechanisms of plant extracts against MPTP induced neurotoxicity: Future applications in Parkinson’s disease. Biomed. Pharmacother. 2017, 85, 635–645. [Google Scholar] [CrossRef]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Requejo-Aguilar, R.; Bolaños, J.P. Mitochondrial control of cell bioenergetics in Parkinson’s disease. Free Radic. Biol. Med. 2016, 100, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, P.; Szalai, P.; Olesen, C.; Praetorius, H.A.; Nissen, P.; Christensen, S.B.; Engedal, N.; Møller, J.V. Inhibition of the sarco/endoplasmic reticulum (ER) Ca2+-ATPase by thapsigargin analogs induces cell death via ER Ca2+ depletion and the unfolded protein response. J. Biol. Chem. 2017, 292, 19656–19673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, Y.-i.; Kuranaga, E. Caspase-dependent non-apoptotic processes in development. Cell Death Differ. 2017, 24, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Thornberry, N.A.; Rano, T.A.; Peterson, E.P.; Rasper, D.M.; Timkey, T.; Garcia-Calvo, M.; Houtzager, V.M.; Nordstrom, P.A.; Roy, S.; Vaillancourt, J.P. A combinatorial approach defines specificities of members of the caspase family and granzyme B Functional relationships established for key mediators of apoptosis. J. Biol. Chem. 1997, 272, 17907–17911. [Google Scholar] [CrossRef] [Green Version]
- Bressenot, A.; Marchal, S.; Bezdetnaya, L.; Garrier, J.; Guillemin, F.; Plénat, F. Assessment of apoptosis by immunohistochemistry to active caspase-3, active caspase-7, or cleaved PARP in monolayer cells and spheroid and subcutaneous xenografts of human carcinoma. J. Histochem. Cytochem. 2009, 57, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.; Stamelou, M.; Goetz, C.; Poewe, W.; Lang, A.; Weintraub, D.; Burn, D.; Halliday, G.M.; Bezard, E.; Przedborski, S. Past, present, and future of Parkinson’s disease: A special essay on the 200th Anniversary of the Shaking Palsy. Mov. Disord. 2017, 32, 1264–1310. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.; Gordon, G.; Hand, A.; Walker, R.W.; Fisher, J.M. 200 Years of Parkinson’s disease: What have we learnt from James Parkinson? Age Ageing 2018, 47, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voon, V.; Napier, T.C.; Frank, M.J.; Sgambato-Faure, V.; Grace, A.A.; Rodriguez-Oroz, M.; Obeso, J.; Bezard, E.; Fernagut, P.-O. Impulse control disorders and levodopa-induced dyskinesias in Parkinson’s disease: An update. Lancet Neurol. 2017, 16, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Espay, A.J.; Morgante, F.; Merola, A.; Fasano, A.; Marsili, L.; Fox, S.H.; Bezard, E.; Picconi, B.; Calabresi, P.; Lang, A.E. Levodopa-induced dyskinesia in Parkinson disease: Current and evolving concepts. Ann. Neurol. 2018, 84, 797–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.J.; Dallapiazza, R.F.; De Vloo, P.; Lozano, A.M. Current surgical treatments for Parkinson’s disease and potential therapeutic targets. Neural Regen. Res. 2018, 13, 1342. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.J.; van Staden, J. Cytotoxicity studies of lycorine alkaloids of the Amaryllidaceae. Nat. Prod. Commun. 2014, 9, 1934578X1400900834. [Google Scholar] [CrossRef]
- Adewusi, E.A.; Fouche, G.; Steenkamp, V. Cytotoxicity and acetylcholinesterase inhibitory activity of an isolated crinine alkaloid from Boophane disticha (Amaryllidaceae). J. Ethnopharmacol. 2012, 143, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.J.; van Staden, J. Pharmacological and toxicological insights to the South African Amaryllidaceae. Food Chem. Toxicol. 2013, 62, 262–275. [Google Scholar] [CrossRef]
- Jin, A.; Li, X.; Zhu, Y.-Y.; Yu, H.-Y.; Pi, H.-F.; Zhang, P.; Ruan, H.-L. Four new compounds from the bulbs of Lycoris aurea with neuroprotective effects against CoCl2 and H2O2-induced SH-SY5Y cell injuries. Arch. Pharmacal Res. 2014, 37, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Omoruyi, S.I.; Delport, J.; Kangwa, T.S.; Ibrakaw, A.S.; Cupido, C.N.; Ekpo, O.E.; Hussein, A.A. In vitro neuroprotective potential of Clivia miniata and Nerine humilis (Amaryllidaceae) in MPP+-induced neuronal toxicity in SH-SY5Y neuroblastoma cells. S. Afr. J. Bot. 2020, 136, 110–117. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Amaryllidaceae alkaloids: Absolute configuration and biological activity. Chirality 2017, 29, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Qu, D.; Zhang, K.-M.; Cang, X.-X.; Kou, Z.-N.; Xiao, W.; Zhu, J.-B. Phytochemical and biological investigations of Amaryllidaceae alkaloids: A review. J. Asian Nat. Prod. Res. 2017, 19, 53–100. [Google Scholar] [CrossRef]
- Hulcová, D.; Breiterová, K.; Siatka, T.; Klímová, K.; Davani, L.; Šafratová, M.; Hošťálková, A.; De Simone, A.; Andrisano, V.; Cahlíková, L. Amaryllidaceae alkaloids as potential glycogen synthase kinase-3β inhibitors. Molecules 2018, 23, 719. [Google Scholar] [CrossRef] [Green Version]
- Sandager, M.; Nielsen, N.D.; Stafford, G.I.; van Staden, J.; Jäger, A.K. Alkaloids from Boophane disticha with affinity to the serotonin transporter in rat brain. J. Ethnopharmacol. 2005, 98, 367–370. [Google Scholar] [CrossRef]
- Elgorashi, E.E.; Stafford, G.I.; Jäger, A.K.; van Staden, J. Inhibition of [3H] citalopram binding to the rat brain serotonin transporter by Amaryllidaceae alkaloids. Planta Med. 2006, 72, 470–473. [Google Scholar] [CrossRef]
- Neergaard, J.S.; Andersen, J.; Pedersen, M.E.; Stafford, G.I.; Staden, J.V.; Jäger, A.K. Alkaloids from Boophone disticha with affinity to the serotonin transporter. S. Afr. J. Bot. 2009, 75, 371–374. [Google Scholar] [CrossRef] [Green Version]
- Crews, L.; Masliah, E. Molecular mechanisms of neurodegeneration in Alzheimer’s disease. Hum. Mol. Genet. 2010, 19, R12–R20. [Google Scholar] [CrossRef]
- Huang, W.-J.; Zhang, X.; Chen, W.-W. Role of oxidative stress in Alzheimer’s disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgorashi, E.E.; Stafford, G.I.; Van Staden, J. Acetylcholinesterase enzyme inhibitory effects of Amaryllidaceae alkaloids. Planta Med. 2004, 70, 260–262. [Google Scholar] [PubMed]
- Moon, H.E.; Paek, S.H. Mitochondrial Dysfunction in Parkinson’s Disease. Exp. Neurobiol. 2015, 24, 103–116. [Google Scholar] [CrossRef]
- Burté, F.; De Girolamo, L.A.; Hargreaves, A.J.; Billett, E.E. Alterations in the mitochondrial proteome of neuroblastoma cells in response to complex 1 inhibition. J. Proteome Res. 2011, 10, 1974–1986. [Google Scholar] [CrossRef]
- Piao, Y.; Kim, H.G.; Oh, M.S.; Pak, Y.K. Overexpression of TFAM, NRF-1 and myr-AKT protects the MPP+-induced mitochondrial dysfunctions in neuronal cells. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 577–585. [Google Scholar] [CrossRef]
- Jeong, K.H.; Jeon, M.-T.; Kim, H.D.; Jung, U.J.; Jang, M.C.; Chu, J.W.; Yang, S.J.; Choi, I.Y.; Choi, M.-S.; Kim, S.R. Nobiletin protects dopaminergic neurons in the 1-methyl-4-phenylpyridinium-treated rat model of Parkinson’s disease. J. Med. Food 2015, 18, 409–414. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, J.-Y.; Chin, M.H.; Schepmoes, A.A.; Petyuk, V.A.; Weitz, K.K.; Petritis, B.O.; Monroe, M.E.; Camp, D.G.; Wood, S.A. Region-specific protein abundance changes in the brain of MPTP-induced Parkinson’s disease mouse model. J. Proteome Res. 2010, 9, 1496–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepule, K.H.; Cordier, W.; Steenkamp, P.; Nell, M.; Steenkamp, V. The ability of three African herbal remedies to offer protection against an in vitro model of Parkinson’s disease. S. Afr. J. Bot. 2019, 126, 121–131. [Google Scholar] [CrossRef]
- Ito, K.; Eguchi, Y.; Imagawa, Y.; Akai, S.; Mochizuki, H.; Tsujimoto, Y. MPP+ induces necrostatin-1- and ferrostatin-1-sensitive necrotic death of neuronal SH-SY5Y cells. Cell Death Discov. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julien, O.; Wells, J.A. Caspases and their substrates. Cell Death Differ. 2017, 24, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Waldmeier, P.C.; Tatton, W.G. Interrupting apoptosis in neurodegenerative disease: Potential for effective therapy? Drug Discov. Today 2004, 9, 210–218. [Google Scholar] [CrossRef]
- Egunlusi, A.O.; Malan, S.F.; Omoruyi, S.I.; Ekpo, O.E.; Palchykov, V.A.; Joubert, J. Open and rearranged norbornane derived polycyclic cage molecules as potential neuroprotective agents through attenuation of MPP+-and Calcium overload-induced excitotoxicity in neuroblastoma SH-SY5Y cells. Eur. J. Med. Chem. 2020, 204, 112617. [Google Scholar] [CrossRef] [PubMed]
- Ibrakaw, A.S.; Omoruyi, S.I.; Ekpo, O.E.; Hussein, A.A. Neuroprotective activities of Boophone haemanthoides (Amaryllidaceae) extract and its chemical constituents. Molecules 2020, 25, 5376. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pancratinine B (1) | Bufanidrine (2) | Buphanisine (3) | Epibuphanisine (4) | |||||
|---|---|---|---|---|---|---|---|---|
| No | δC | δ, J (Hz) | δC | δ, J (Hz) | δC | δ, J (Hz) | δC | δ, J (Hz) |
| 1 | 133.4 d | 6.03 br d, 10.4 | 132.9 d | 6.58 d, 10.2 | 132.9 d, | 6.55 d, 9.8 | 132.1 d | 6.58 d, 10.4 |
| 2 | 131.7 d | 5.70 br d, 10.4 | 125.3 d | 5.95 dd, 10.2, 5.2 | 125.4 d, | 5.95 dd, 9.8, 5.2 | 125.6 d | 5.96 dd, 10.4, 5.4 |
| 3 | 72.3 d | 3.94 br dd, 5.0, 10.8 | 72.6 d | 3.82 * | 77.2 d | 3.81 * | 72.3 d | 3.80 ddd, 5.4, 4.3, 1.7 |
| 4 | 30.9 t | 1.50 ddd, 4.6, 11.0, 12.8 | 28.7 t | 2.08 m | 28.7 t, | 2.07 ddd, 13.3, 4.3, 2.0 | 28.2 t | 2.20 * |
| 2.35 br d, 12.9 | 1.59 dt, 13.7, 4.0 | 1.58 td, 13.3, 4.3 | 1.62 td, 13.4, 4.2 | |||||
| 4a | 67.5 d | 2.95 m | 62.6 d | 3.31 dd, 13.7, 4.0 | 63.1 d | 3.43 # | 63.3 d | 3.32 ** |
| 6 | 61.8 t | 3.83 d, 16.8 | 58.6 t | 4.24 d, 17.6 | 62.3 t | 4.45 d, 16.7 | 61.8 t | 4.38 d, 16.7 |
| 4.26 d, 16.8 | 3.81 * d, 17.6 | 3.83 * d, 16.7 | 3.75 d, 16.7 | |||||
| 6a | 125.4 s | 117.3 s | 126.2 s | 125.6 s | ||||
| 7 | 106.8 d | 6.51 s | 139.4 s | 106.9 s | 6.45 s | 106.9 d | 6.46 s | |
| 8 | 145.9 s | 133.4 s | 145.7 s | 145.9 s | ||||
| 9 | 147.4 s | 148.0 s | 146.0 s | 146.4 s | ||||
| 10 | 109.9 d | 6.63 s | 96.6 d | 6.81 s | 102.9 d | 6.82 s | 103.0 d | 6.81 s |
| 10a | 129.4 s | 139.3 s | 138.4 s | 137.7 s | ||||
| 10b | 44.3 s | 44.3 s | 44.5 s | |||||
| 11 (exo) | 49.2 t | 2.64 d, 2.2 | 44.0 t | 2.15, ddd, 12.7, 9.8, 4.8 | 44.2 t | 2.15 ddd, 12.4, 9.2, 4.3 | 43.6 t | 1.89 ddd, 10.6, 10.6, 5.2 |
| 11 (endo) | 1.90 ddd, 11.8, 10.6, 5.8 | 1.90 ddd, 12.4, 11.0, 6.0 | 2.20 * | |||||
| 11b | 82.5 s | |||||||
| 12 (exo) | 54.7 t | 2.82 d, 11.8 | 53.6 t | 3.37 ** | 53.4 t | 3.45 # | 53.3 t | 3.46 |
| 12 (endo) | 2.97 dd, 2.2, 11.8 | 2.87, ddd, 14.0, 9.0, 6.0 | 2.93 ddd, 12.0, 9.0, 6.0 | 2.94 ddd, 14.0, 9.0, 6.0 | ||||
| OCH2O | 100.9 t | 5.92 s | 100.5 t | 5.85, 5.83 d/each, 1.40 | 100.7 t | 5.88, 5.87 br s/each | 100.9 t | 5.86, 5.85 d, 1.3 |
| OMe-3 | 56.3 q | 3.42 s | 56.4 q | 3.35 ** s | 56.5 q | 3.34 s | 56.6 q | 3.34 ** s |
| OMe-7 | 59.1 q | 3.96 s | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omoruyi, S.I.; Ibrakaw, A.S.; Ekpo, O.E.; Boatwright, J.S.; Cupido, C.N.; Hussein, A.A. Neuroprotective Activities of Crossyne flava Bulbs and Amaryllidaceae Alkaloids: Implications for Parkinson’s Disease. Molecules 2021, 26, 3990. https://doi.org/10.3390/molecules26133990
Omoruyi SI, Ibrakaw AS, Ekpo OE, Boatwright JS, Cupido CN, Hussein AA. Neuroprotective Activities of Crossyne flava Bulbs and Amaryllidaceae Alkaloids: Implications for Parkinson’s Disease. Molecules. 2021; 26(13):3990. https://doi.org/10.3390/molecules26133990
Chicago/Turabian StyleOmoruyi, Sylvester I., Abobaker S. Ibrakaw, Okobi E. Ekpo, James S. Boatwright, Christopher N. Cupido, and Ahmed A. Hussein. 2021. "Neuroprotective Activities of Crossyne flava Bulbs and Amaryllidaceae Alkaloids: Implications for Parkinson’s Disease" Molecules 26, no. 13: 3990. https://doi.org/10.3390/molecules26133990