1. Introduction
Mushrooms have great prospects in medicine and nutraceutical production. In addition to having good organoleptic properties and high nutritional values, many mushrooms were reported to have a myriad of pharmacological activities [
1,
2,
3]. Among the most widely cultivated mushrooms are
Lentinula edodes (shiitake mushroom) and
Pleurotus ostreatus (oyster mushroom). Shiitake comes in second to
Agaricus bisporus as the most consumed edible mushroom worldwide [
4]. This mushroom has promising antibacterial, antifungal, antiviral, hepatoprotective, antihyperglycemic and immunomodulatory effects [
5,
6,
7]. It is rich in bioactive molecules, the most studied of which is “lentinan”, a polysaccharide with an effect against bacteria, viruses and tumors, in addition to “lentinacin”, which showed help in controlling dyslipidemia and hyperglycemia [
8].
Pleurotus ostreatus, the most common species of the genus
Pleurotus [
9], also known as the oyster mushroom, is a wood decomposer and has a broad range of biological activities [
10]. In comparison to other therapeutic mushrooms, oyster mushrooms are becoming more popular as health promoters [
11]. The presence of a large number of nutritious components such as lectins, polysaccharides, vitamins and minerals in oyster mushrooms makes them able to possess potential anticancer, antioxidant, antidiabetic, antimicrobial and anti-hypercholesterolemic properties [
12,
13]. In comparison to other edible mushrooms, oyster and shiitake mushrooms have a brief growing period and can be harvested throughout the year [
14]. As a result of their ease of cultivation and testing, high nutritional values and promising medicinal benefits, these mushrooms have tremendous potential in the food and pharmaceutical industries [
4,
15]. Indeed, higher Basidiomycetes can be promising multifunctional food sources.
Many bioactive compounds, including sugars, physiologically active proteins, un-saturated fatty acids, phenolics (phenolic acids and polyphenols), flavonoids, terpenoids, glycoproteins, polyketides, steroids and alkaloids, were found in these two mushrooms [
16,
17]. These compounds act solely or synergistically to bring about the broad pharmacological actions of these fungi. Shiitake and oyster mushrooms are reported to have antibacterial, antiviral, antihypertensive, immunomodulatory and antioxidant activities [
1,
18,
19]. Seo et al. [
20] summarized the mechanisms of the antiviral activity of mushroom biomolecules, indicating that the reduction in viral infection is mainly through interfering with the uptake of the virus into host cells, its replication and its protein and enzyme synthesis, in addition to stimulating the host immune response [
21]. Antioxidants are important molecules in the face of reactive oxygen species which are behind many health problems. Since many red flags were raised against the use of synthetic antioxidants, the study of natural ones has become mandatory [
22]. The antitumor effect of these mushroom products was related to biomolecules including glucans, ergosterol, proteoglucans and amino acids (arginine and glutamine) [
23]. The postulated mechanism underlying this effect may include the stimulation of T-lymphocytes, suppression of neovascularization and induction of cancer cell death, in addition to triggering the immune response against cancer cells [
24,
25].
Proteomics has proven itself as one of the valuable tools in bioresearch, particularly agricultural research [
26]. Lindequist et al. [
27] were the first group to shed light on the importance of employing proteomics in edible mushroom research. They emphasized the need for using “omics” for the study of fungal bioactive molecules. Proteomics has been used scarcely in edible mushroom research vis à vis pathogenic fungi [
28]. Accordingly, the aims of this study were to analyze the proteome of the two edible mushrooms,
Pleurotus ostreatus and
Lentinula edodes, and to investigate their potential antiviral, antitumor and antioxidant activities.
3. Discussion
White rot fungi have stirred the scientific community’s interest due to their medicinal properties, which include immune system modulation, hypoglycemic and antithrombotic activity and antihypertensive, anti-inflammatory, antimicrobial and antitumor properties, as well as the ability to lower blood cholesterol levels [
24,
36,
37,
38]. The two white rot fungi
P. ostreatus and
L. edodes are two of the most highly consumed edible mushrooms in many countries [
39]. A multitude of bioactive proteins are produced by these two mushrooms, and proteome analysis is currently the most effective tool for protein profiling [
40]. In the current study, proteome analysis showed the expression of bioactive proteins including the Rab GDP dissociation inhibitor, thioredoxin reductase, serine proteinase, superoxide dismutase and lectin in the two test mushrooms. The Rab GDP dissociation inhibitor (Rab GDI) regulates the function of Rab GTPases which play a pivotal role in membrane trafficking in tumor cells. Accordingly, using Rab GDI is a promising anticancer strategy [
29].
Serine proteinase contributes to the antiviral activity of edible mushrooms [
30,
31]. Moreover, Yap et al. (2018) reported a strong selective cytotoxicity of serine proteinase in the face of a human breast adenocarcinoma cell line (MCF7) and suggested that the mechanism involves the collaborative effect of both extrinsic and intrinsic cell death mechanisms, in addition to the stimulation of caspase-8 and -9 and inhibition of Bcl-2 [
41].
The intake of antioxidants is an auspicious prophylactic strategy against reactive oxygen species (ROS)-mediated pathophysiology [
22]. Our results show the expression of enzymes of the antioxidant defense system by
Pleurotus ostreatus and
Lentinula edodes. These enzymes include superoxide dismutase, catalase and glutathione peroxidase, which counterbalance the production of ROS [
32]. Superoxide dismutase starts by converting superoxides into hydrogen peroxide, which is, in turn, converted by catalase and glutathione peroxidase into water [
33].
The expression of lectin was also detected in the two mushrooms. Many researchers reported the remarkably diverse biological profiles of lectin with a vast range of activities encompassing anti-inflammatory, antidepressant, anticonceptive and vasodilatory activities [
34,
35,
42]. Lectins are carbohydrate-binding proteins with a variety of cellular functions including in vitro and in vivo suppression of tumor growth via the selective binding to tumor cell membranes or their receptors, resulting in the activation of protein kinases, or modulation of immune responses through interleukin production [
43]. Additionally, mushroom lectins also have a role in triggering different cell death pathways, including apoptosis, necrosis and/or autophagy [
42].
Our data show the expression of both ostreolysin and pleurotolysin in
Pleurotus ostreatus. These are pore-forming proteins with highly selective anticancer activities [
44,
45]. Ostreolysin is a 15 kDa cytolytic protein with the ability to permeabilize erythrocytes and other cells at sub-micromolar concentrations. It acts via a colloid osmotic mechanism and induces the formation of wide membrane pores [
46,
47]. Nimri et al. produced recombinant pleurotolysin with potent antitumor activity against human and mouse colon tumor cells [
48].
In
L. edodes, valosin-containing protein and latcripin were expressed. Valosin-containing protein was previously reported to have antioxidant properties which could help in neuronal syndromes such as Alzheimer’s disease, lateral sclerosis and dementia [
49,
50,
51]. Latcripin is a potential anticancer agent [
50]. Riaz Ud Din et al. (2020) investigated the anticancer mechanism of latcripin against breast cancer cell lines. They reported its ability to induce cell death as well as autophagy, in addition to its inhibitory effect on migration and invasion [
52].
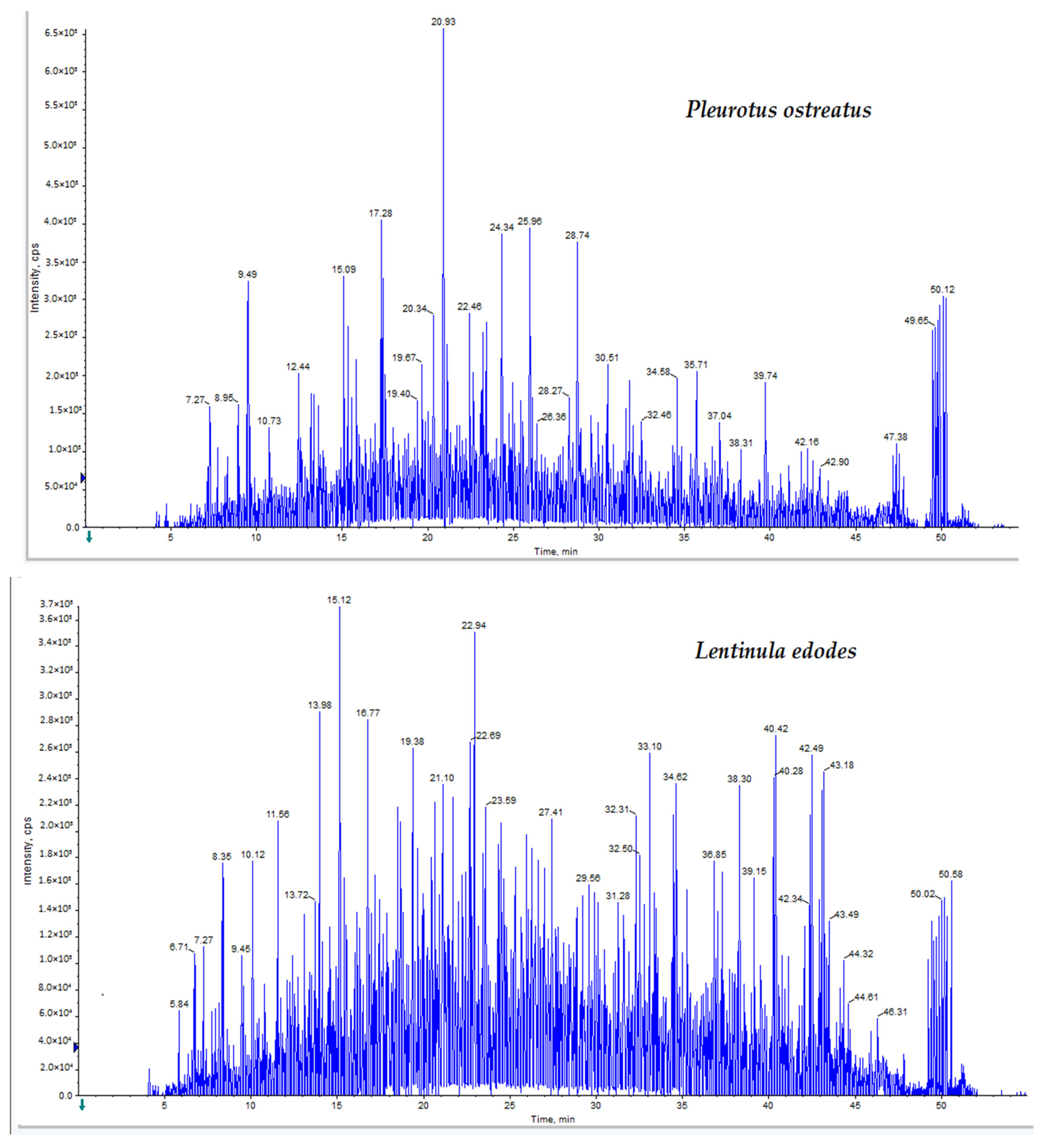
A range of bioactive compounds were quantified in
P. ostreatus and
L. edodes extracts. TPNs of 19.37 ± 0.39 and 24.14 ± 1.01 mg/g of extract for
P. ostreatus and
L. edodes, respectively, were detected in the current study. Those values are higher than those recorded by Rahimah et al., who used the ammonium and Shinoda tests to quantify the total flavonoid content of
P. ostreatus as 6.67 mg/g of extract [
53]. Additionally, our results are higher than those found by Montibus et al., who assayed the TPC in
L. edodes extracts using the Folin–Ciocalteu method and found 0.8–1.5% dry weight (dw) in the caps and 0.8–1.1% dw in the stipes [
54].
Catechin was detected in both
P. ostreatus and
L. edodes extracts. Catechin is a plant secondary phenolic metabolite, with potent free radical scavenging properties [
55]. The
L. edodes extract was found to contain the flavonoid quercetin. The antioxidant effect of quercetin is mediated via the regulation of glutathione levels and an increase in the production of antioxidant enzymes including glutathione transferase and aldo-keto reductase [
56]. In addition, quercetin is reported to have antitumor activity through interrupting the cell cycle and promoting apoptosis [
57,
58]. Lee et al. reported that quercetin causes cell arrest in the PC3, Du145 and U937 cancer cell lines [
59]. Our results also reveal that kaempferol and apigenin were found in
P. ostreatus. Kaempferol and apigenin have been proven to have antioxidant and anti-proliferative activities [
60,
61,
62]. Glutamic acid was found to be the most abundant amino acid in both mushroom extracts. In the same vein, Chirinang et al. analyzed the amino acid content of
P. ostreatus and
P. sajor-caju and found that the glutamic acid content was the highest, followed by aspartic acid and then arginine [
63].
P. ostreatus and L. edodes showed promising antiviral activities against adenovirus and
herpes simplex virus-II, with the SI reaching 4.5 for the
P. ostreatus extract against adenovirus. The SI was used to evaluate the efficacy and safety of the extracts as it estimates the window between the cytotoxic and antiviral activities. The higher the SI, the safer and more efficient the compound [
64,
65]. Related data were recently shown by Urbancikova et al., where pleuran (insoluble β-1,3/1,6-D-glucan isolated from
P. ostreatus)-based supplements significantly shortened the duration of
herpes simplex virus-I symptoms, with a lower severity of respiratory symptoms, in
herpes simplex virus-I-positive patients than the placebo group, without significant side effects, proposing pleuran for possible future use in the treatment of acute
herpes simplex virus-I [
66].
The antiviral effect of mushrooms is mostly due to interfering with viral uptake, replication, enzyme activity and functioning peptides as well as potentiating the host immune system [
67]. Seo and Choi (2021) proposed that the β-glucan in the polysaccharide fraction of
P. ostreatus and
L. edodes may be responsible for its antiherpetic effect through pre- and post-treatment effects [
20]. The anti-HIV effect of the aqueous extracts of
P. ostreatus and
L. edodes may be caused by inhibition of the reverse transcriptase enzyme by ubiquitin-like protein and lentin, respectively [
68,
69].
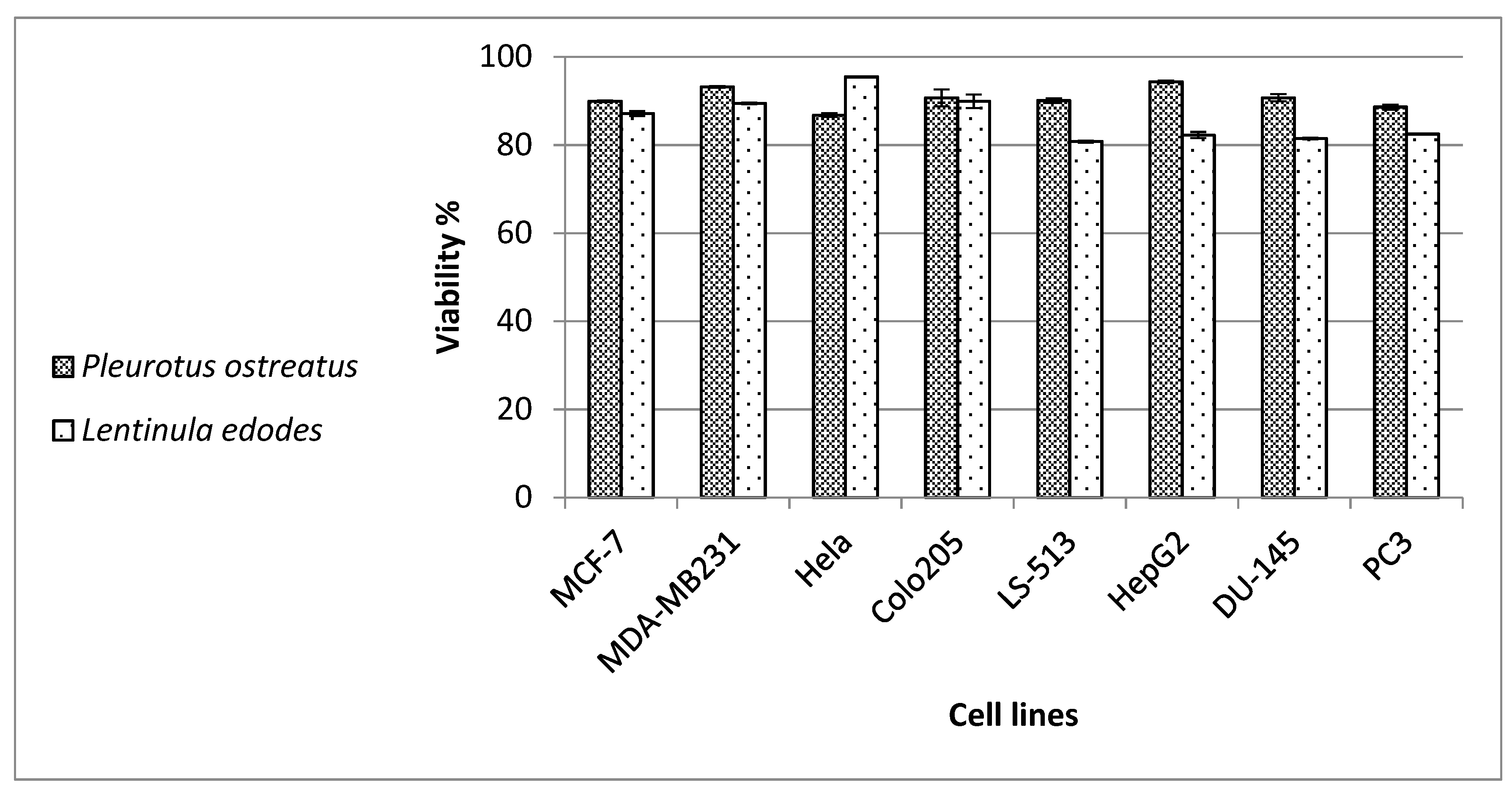
The aqueous extracts of our mushrooms were shown to inhibit the viability of the tested cancer cell lines by approximately 20%. We also saw similar effects against leukemia and lymphoma cell lines, with a decrease in viability to 66% with the
L. edodes extract against THP1 leukemia cells, and to 70.6% with the
P. ostreatus extract against CCRF-CEM leukemia cells. Aqueous mycelial and fruit body extracts of
L. edodes were previously reported to exert anti-proliferative and apoptotic actions on MCF-7 breast cancer cells [
70]. The ethyl acetate fraction and β-glucan of
L. edodes were mostly responsible for these actions [
71,
72]. In the same context, the
P. ostreatus extract was able to show anti-proliferative activity toward MCF-7 and MDA-MB-231 breast adenocarcinoma cells [
73], and the 6-linked glucans of the extract potentiated the natural killer cytotoxicity against breast and lung cancer cells [
74]. Recently, Jakopovic et al. found that medicinal mushroom preparations consisting of 6 and 10 mushrooms including
L. edodes and
P. ostreatus exhibited significant anti-proliferative and pro-apoptotic effects on colorectal (HCT-116, SW620) tumor cell lines [
75]. On the contrary, the authors noticed that the effect on a human fibroblast cell line (WI-38) was proliferative, showing the specificity of these mushroom preparations towards tumor cell lines. Similarly, we detected minimal cytotoxicity excreted by our mushrooms against normal human PBMCs. This finding also suggests the safe use of our mushrooms.
The cytotoxicity of mushrooms has been attributed to a wide range of molecules including α- and β-glucans, proteins, glycoproteins, fatty acids, nucleoside antagonists, terpenoids and phenolic compounds [
76]. Abdalla et al. (2012) proposed that mushroom extracts suppressed breast cancer cell proliferation by inhibiting aromatase activity [
77]. Imam et al. (2021) isolated an indole-3-lactic acid from
L. edodes that inhibited the division of lung adenocarcinoma cells [
78]. Yukawa et al. (2012) suggested that the direct apoptotic effect of
L. edodes mycelia on HepG2 cells is through activation of the caspase-3 and -8 death receptor pathways [
79]. Wu et al. (2011) studied the cytotoxic effect of the protein extract of
P. ostreatus on human colorectal adenocarcinoma (SW480) cells and a human monocytic leukemia, THP-1 (cells), and reported generation of ROS, exhaustion of glutathione, alteration of mitochondrial membrane potential and disintegration of oligonucleosomal DNA, resulting in apoptosis of SW480 cells [
80].
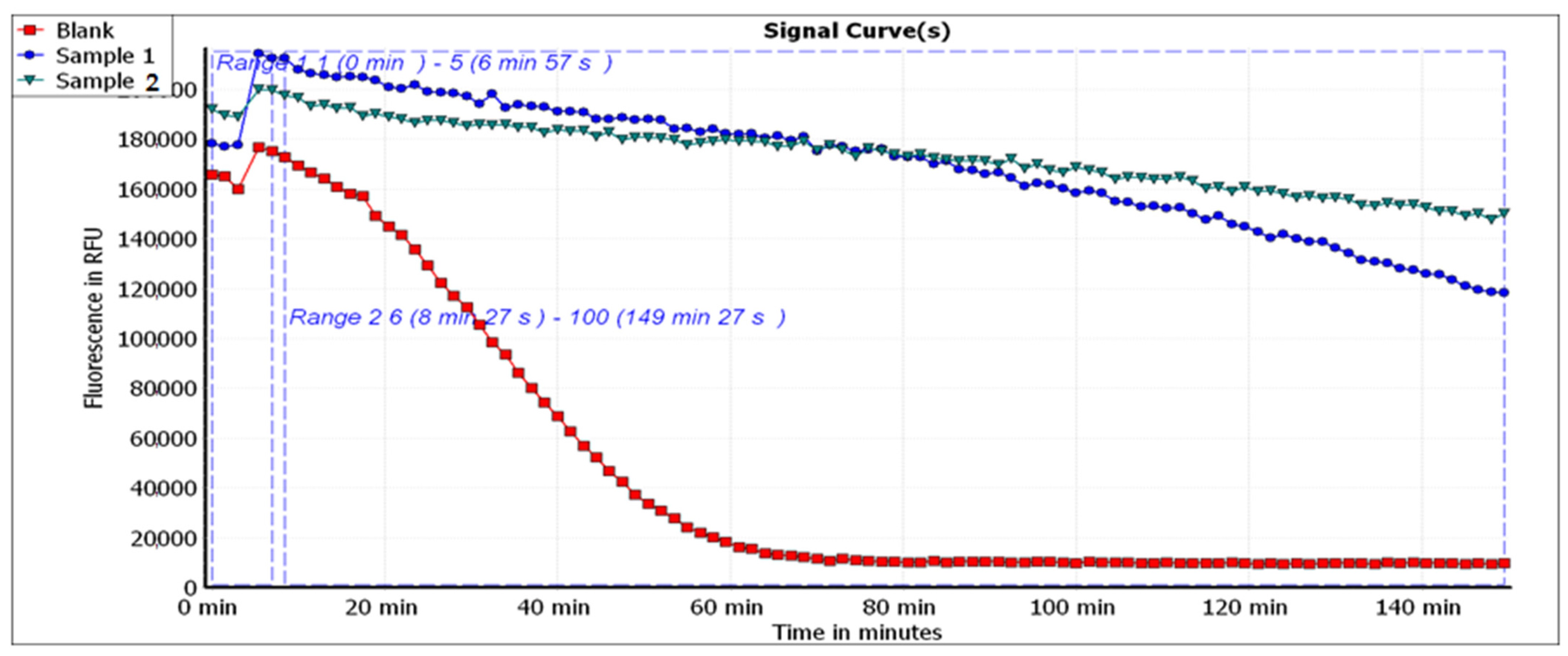
As antioxidants do not act through a single mechanism, evaluation of antioxidant capacity is usually conducted through more than one method [
81]. In this study, three assessment methods were used, namely, DPPH, ABTS and ORAC assays. The SET (single electron transfer) antioxidant mechanism was evaluated through the DPPH assay, while the ABTS and ORAC assays were used for HAT (hydrogen atom transfer) reactions [
81]. The ORAC assay provides more precise accurate estimates as it combines the inhibition time and the degree of inhibition in a single term [
82]. While the phosphomolybdate assay is commonly used to express the total antioxidant capacity (TAC), all three methods used in this manuscript have been reported to evaluate the TAC. Munteanu and Apetrei (2021), in their review on the methods used in determining antioxidant activity, described the ABTS radical scavenging assay as a “simple and convenient method used to measure the total antioxidant capacity (TAC)” [
83]. In their comparative study, Csepregi et al. (2016) compared four methods used for evaluation of the TAC, which were Trolox equivalent antioxidant capacity (TEAC), the ferric reducing antioxidant potential (FRAP), the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay and Folin–Ciocalteu reagent reactivity (FCR) [
84]. In 2013, Ou et al. introduced ORAC as a novel method for evaluation of the TAC [
85]. In the same vein, Rubio et al. (2016) classified the ORAC assay as being among the direct methods for the determination of the TAC, together with TEAC [
86]. The results of the ABTS and ORAC assays show that the
P. ostreatus and
L. edode extracts have antioxidant capacities that are higher than those of the standard antioxidant, Trolox.
The antioxidant effect of the two mushrooms may be attributed to the presence of many bioactive components including flavonoids, phenolics, bioactive peptides and vitamin C [
87]. Gaber et al. compared the ABTS, DPPH, ferric reducing antioxidant power (FRAP) and ORAC assays for assessing the antioxidant capacity of guava extracts and reported a positive correlation between antioxidant powers, as determined by the four methods, and the vitamin C content and TPN. On the other hand, they recorded a negative correlation with the carotenoid content [
88]. DPPH (1,1-Diphenyl-2-picryl-hydrazyl) is a stable free radical which has an unpaired valence electron at one atom of the nitrogen bridge; thus, scavenging of the DPPH radical is the basis of the popular DPPH antioxidant assay [
89]. The 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonate) radical cation (ABTS) loses its blue color when reduced by an antioxidant, and the color alteration can be quantified spectrophotometrically [
90]. The ORAC assay measures the oxidative degradation of the fluorescence probe by the free radicals which are interrupted by the antioxidant substance. Accordingly, the stronger the antioxidant capacity, the shorter the time needed to quench the fluorescence [
91]. Additionally, since the tested extracts showed promising antioxidant effects when tested through the above-mentioned methods, it can be concluded that they act through free radical scavenging. However, the tested mushrooms contained considerable total flavonoid contents, and as flavonoids have been linked to the activation of the transcriptional factor Nrf2 (2017), this can be added as a putative mechanism [
92].



 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}