
Enhanced Oxytetracycline Production by Streptomyces rimosus in Submerged Co-Cultures with Streptomyces noursei

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
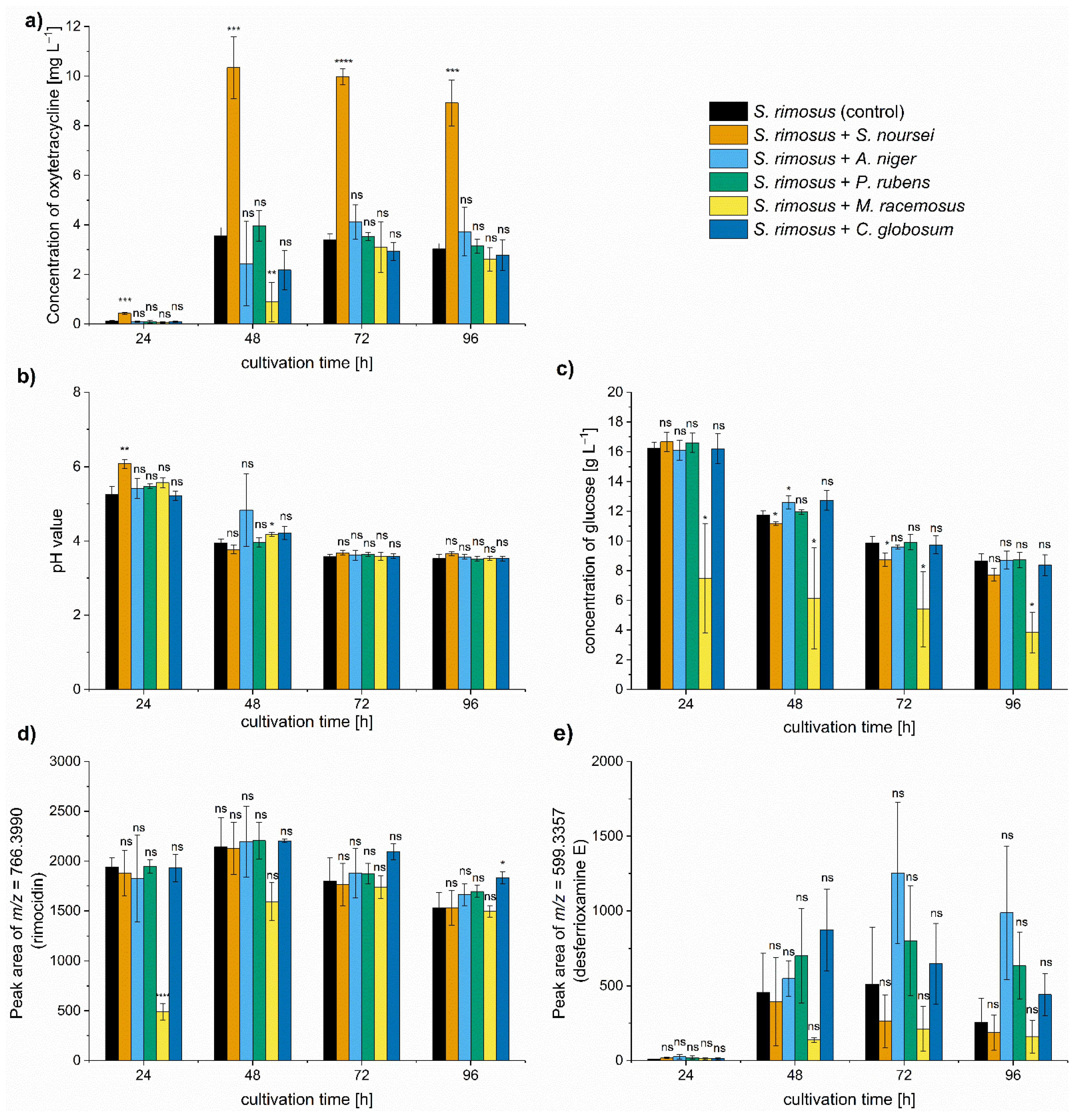
2. Results
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Cultivation
4.3. Chemical Analysis
4.4. Morphological Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bérdy, J. Bioactive microbial metabolites: A personal view. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.A.; Banat, A.M.; Abdelhameed, A.M.; Banat, I.M. Streptomyces from traditional medicine: Sources of new innovations in antibiotic discovery. J. Med. Microbiol. 2020, 69, 1040–1048. [Google Scholar] [CrossRef]
- Xia, H.; Li, X.; Li, Z.; Zhan, X.; Mao, X.; Li, Y. The Application of Regulatory Cascades in Streptomyces: Yield Enhancement and Metabolite Mining. Front. Microbiol. 2020, 11, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamehiro, N.; Hosaka, T.; Xu, J.; Hu, H.; Otake, N.; Ochi, K. Innovative Approach for Improvement of an Antibiotic-Overproducing Industrial Strain of Streptomyces albus. Appl. Environ. Microbiol. 2003, 69, 6412–6417. [Google Scholar] [CrossRef] [Green Version]
- Baltz, R.H. Genetic manipulation of secondary metabolite biosynthesis for improved production in Streptomyces and other actinomycetes. J. Ind. Microbiol. Biotechnol. 2016, 43, 343–370. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zheng, G.; Chen, J.; Ge, M.; Jiang, W.; Lu, Y. Multiplexed site-specific genome engineering for overproducing bioactive secondary metabolites in actinomycetes. Metab. Eng. 2017, 40, 80–92. [Google Scholar] [CrossRef]
- Saudagar, P.S.; Singhal, R.S. Optimization of nutritional requirements and feeding strategies for clavulanic acid production by Streptomyces clavuligerus. Bioresour. Technol. 2007, 98, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, Q.; Liu, Y.; Yin, H.; Yang, W.; Wang, Y.; Liu, Y.; Li, Y.; Pang, S.; Liu, W.; et al. Development of a pyrF-based counterselectable system for targeted gene deletion in Streptomyces rimosus. J. Zhejiang Univ. Sci. B 2021, 22, 383–396. [Google Scholar] [CrossRef]
- Ni, H.; Mohsin, A.; Guo, M.; Chu, J.; Zhuang, Y. Two-component system AfrQ1Q2 involved in oxytetracycline biosynthesis of Streptomyces rimosus M4018 in a medium-dependent manner. J. Biosci. Bioeng. 2020, 129, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, S.; Li, Z.; Zhang, J.; Fan, K.; Tan, G.; Ai, G.; Lam, S.M.; Shui, G.; Yang, Z.; et al. Harnessing the intracellular triacylglycerols for titer improvement of polyketides in Streptomyces. Nat. Biotechnol. 2020, 38, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Petković, H.; Lukežič, T.; Šušković, J. Biosynthesis of Oxytetracycline by Streptomyces rimosus: Past, Present and Future Directions in the Development of Tetracycline Antibiotics. Food Technol. Biotechnol. 2017, 55, 3. [Google Scholar] [CrossRef]
- Liao, Z.; Song, Z.; Xu, J.; Ma, Z.; Bechthold, A.; Yu, X. Identification of a gene from Streptomyces rimosus M527 negatively affecting rimocidin biosynthesis and morphological differentiation. Appl. Microbiol. Biotechnol. 2020, 104, 10191–10202. [Google Scholar] [CrossRef]
- Song, Z.; Ma, Z.; Bechthold, A.; Yu, X. Effects of addition of elicitors on rimocidin biosynthesis in Streptomyces rimosus M527. Appl. Microbiol. Biotechnol. 2020, 104, 4445–4455. [Google Scholar] [CrossRef] [PubMed]
- Pethick, F.E.; MacFadyen, A.C.; Tang, Z.; Sangal, V.; Liu, T.T.; Chu, J.; Kosec, G.; Petkovic, H.; Guo, M.; Kirby, R.; et al. Draft genome sequence of the oxytetracycline-producing bacterium Streptomyces rimosus ATCC 10970. Genome Announc. 2013, 1, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, M.; Rückert, C.; Gläser, L.; Beganovic, S.; Luzhetskyy, A.; Kalinowski, J.; Wittmann, C. Microparticles enhance the formation of seven major classes of natural products in native and metabolically engineered actinobacteria through accelerated morphological development. Biotechnol. Bioeng. 2021, 118, 3076–3093. [Google Scholar] [CrossRef]
- Kowalska, A.; Boruta, T.; Bizukojć, M. Morphological evolution of various fungal species in the presence and absence of aluminum oxide microparticles: Comparative and quantitative insights into microparticle-enhanced cultivation (MPEC). Microbiologyopen 2018, 7, e00603. [Google Scholar] [CrossRef]
- Kowalska, A.; Boruta, T.; Bizukojć, M. Performance of fungal microparticle-enhanced cultivations in stirred tank bioreactors depends on species and number of process stages. Biochem. Eng. J. 2020, 161, 107696. [Google Scholar] [CrossRef]
- Wucherpfennig, T.; Hestler, T.; Krull, R. Morphology engineering—Osmolality and its effect on Aspergillus niger morphology and productivity. Microb. Cell Fact. 2011, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, S.; Onaka, H.; Abe, I. Activation of silent biosynthetic pathways and discovery of novel secondary metabolites in actinomycetes by co-culture with mycolic acid-containing bacteria. J. Ind. Microbiol. Biotechnol. 2019, 46, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, M.; Iwami, M.; Yamashita, M.; Komori, T.; Umehara, K.; Imanaka, H. Biphenomycin A production by a mixed culture. Appl. Environ. Microbiol. 1992, 58, 3879–3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavituna, F.; Luti, K.J.K.; Gu, L. In Search of the E. coli Compounds that Change the Antibiotic Production Pattern of Streptomyces coelicolor During Inter-species Interaction. Enzyme Microb. Technol. 2016, 90, 45–52. [Google Scholar] [CrossRef]
- Luti, K.J.K.; Mavituna, F. Elicitation of Streptomyces coelicolor with E. coli in a bioreactor enhances undecylprodigiosin production. Biochem. Eng. J. 2011, 53, 281–285. [Google Scholar] [CrossRef]
- Böl, M.; Schrinner, K.; Tesche, S.; Krull, R. Challenges of influencing cellular morphology by morphology engineering techniques and mechanical induced stress on filamentous pellet systems—A critical review. Eng. Life Sci. 2021, 21, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Boruta, T.; Górnicka, A.; Grzybowska, I.; Stefaniak, I.; Bizukojć, M. Exploring the extremes: Applying high concentration of yeast extract leads to drastic morphological changes and elimination of (+)-geodin and asterric acid production in Aspergillus terreus submerged cultures. Biotechnol. Lett. 2021, 43, 61–71. [Google Scholar] [CrossRef]
- Boruta, T.; Milczarek, I.; Bizukojc, M. Evaluating the outcomes of submerged co-cultivation: Production of lovastatin and other secondary metabolites by Aspergillus terreus in fungal co-cultures. Appl. Microbiol. Biotechnol. 2019, 103, 5593–5605. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Ríos, D.; López-Agudelo, V.A.; Ramírez-Malule, H.; Neubauer, P.; Junne, S.; Ochoa, S.; Ríos-Estepa, R. A genome-scale insight into the effect of shear stress during the fed-batch production of clavulanic acid by Streptomyces clavuligerus. Microorganisms 2020, 8, 1255. [Google Scholar] [CrossRef]
- Esnault, C.; Dulermo, T.; Smirnov, A.; Askora, A.; David, M.; Deniset-Besseau, A.; Holland, I.B.; Virolle, M.J. Strong antibiotic production is correlated with highly active oxidative metabolism in Streptomyces coelicolor M145. Sci. Rep. 2017, 7, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan-Oropeza, A.; Henry, C.; Lejeune, C.; David, M.; Virolle, M.J. Expression of genes of the Pho regulon is altered in Streptomyces coelicolor. Sci. Rep. 2020, 10, 8492. [Google Scholar] [CrossRef]
- Gonciarz, J.; Kowalska, A.; Bizukojc, M. Application of microparticle-enhanced cultivation to increase the access of oxygen to Aspergillus terreus ATCC 20542 mycelium and intensify lovastatin biosynthesis in batch and continuous fed-batch stirred tank bioreactors. Biochem. Eng. J. 2016, 109, 178–188. [Google Scholar] [CrossRef]
- Elsayed, E.A.; Omar, H.G.; El-Enshasy, H.A. Development of fed-batch cultivation strategy for efficient oxytetracycline production by Streptomyces rimosus at semi-industrial scale. Braz. Arch. Biol. Technol. 2015, 58, 676–685. [Google Scholar] [CrossRef]
- Bizukojc, M.; Pawlak, M.; Boruta, T.; Gonciarz, J. Effect of pH on biosynthesis of lovastatin and other secondary metabolites by Aspergillus terreus ATCC 20542. J. Biotechnol. 2012, 162, 253–261. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boruta, T.; Ścigaczewska, A. Enhanced Oxytetracycline Production by Streptomyces rimosus in Submerged Co-Cultures with Streptomyces noursei. Molecules 2021, 26, 6036. https://doi.org/10.3390/molecules26196036
Boruta T, Ścigaczewska A. Enhanced Oxytetracycline Production by Streptomyces rimosus in Submerged Co-Cultures with Streptomyces noursei. Molecules. 2021; 26(19):6036. https://doi.org/10.3390/molecules26196036
Chicago/Turabian StyleBoruta, Tomasz, and Anna Ścigaczewska. 2021. "Enhanced Oxytetracycline Production by Streptomyces rimosus in Submerged Co-Cultures with Streptomyces noursei" Molecules 26, no. 19: 6036. https://doi.org/10.3390/molecules26196036
APA StyleBoruta, T., & Ścigaczewska, A. (2021). Enhanced Oxytetracycline Production by Streptomyces rimosus in Submerged Co-Cultures with Streptomyces noursei. Molecules, 26(19), 6036. https://doi.org/10.3390/molecules26196036