
Phenolics from Medicinal and Aromatic Plants: Characterisation and Potential as Biostimulants and Bioprotectants

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Phenolic Composition of Medicinal and Aromatic Plants
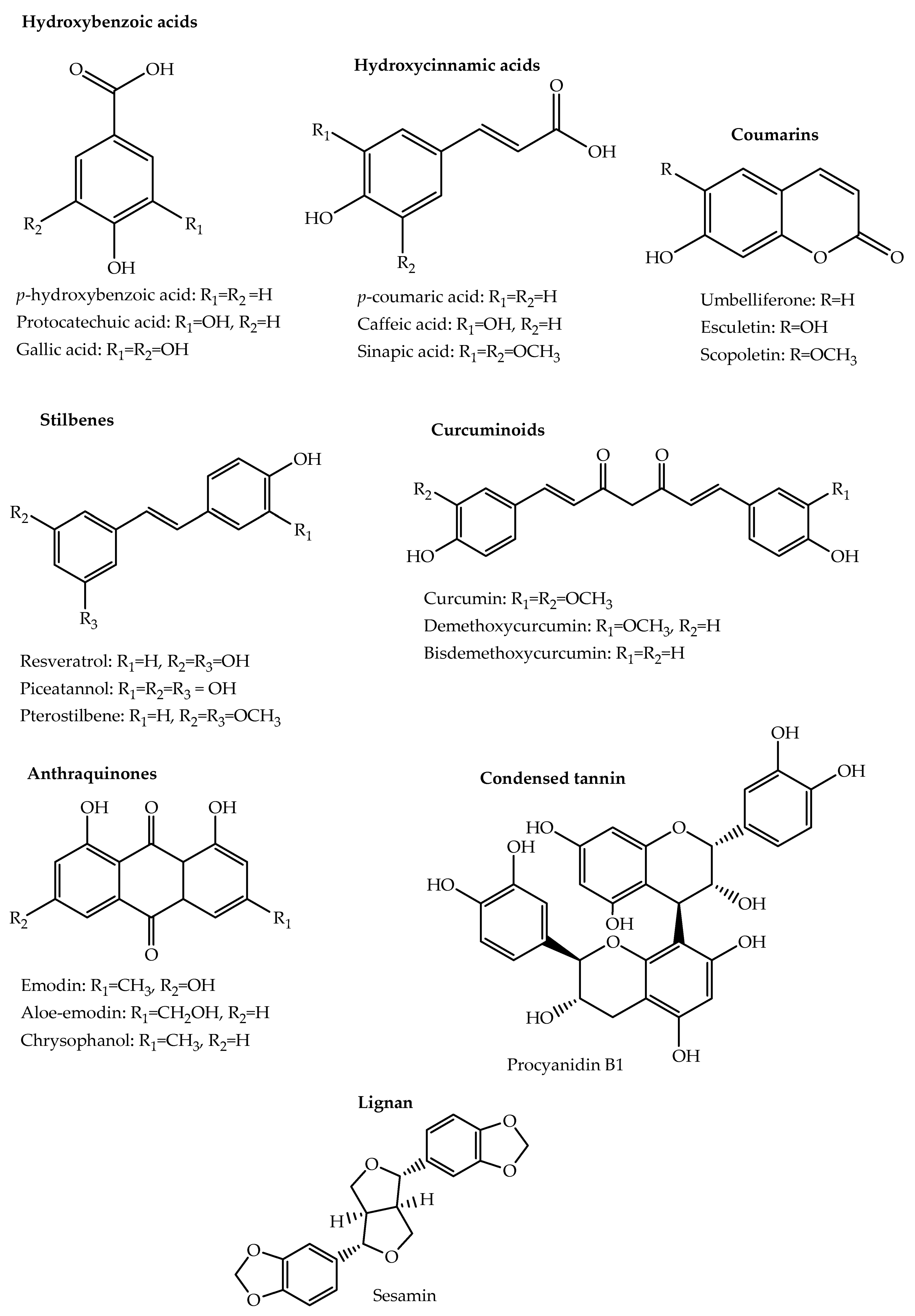
2.1. Phenolic Acids and Their Derivatives
2.2. Stilbenes
2.3. Coumarins
2.4. Lignans
2.5. Quinones
2.6. Curcuminoids
2.7. Flavonoids
2.8. Tannins
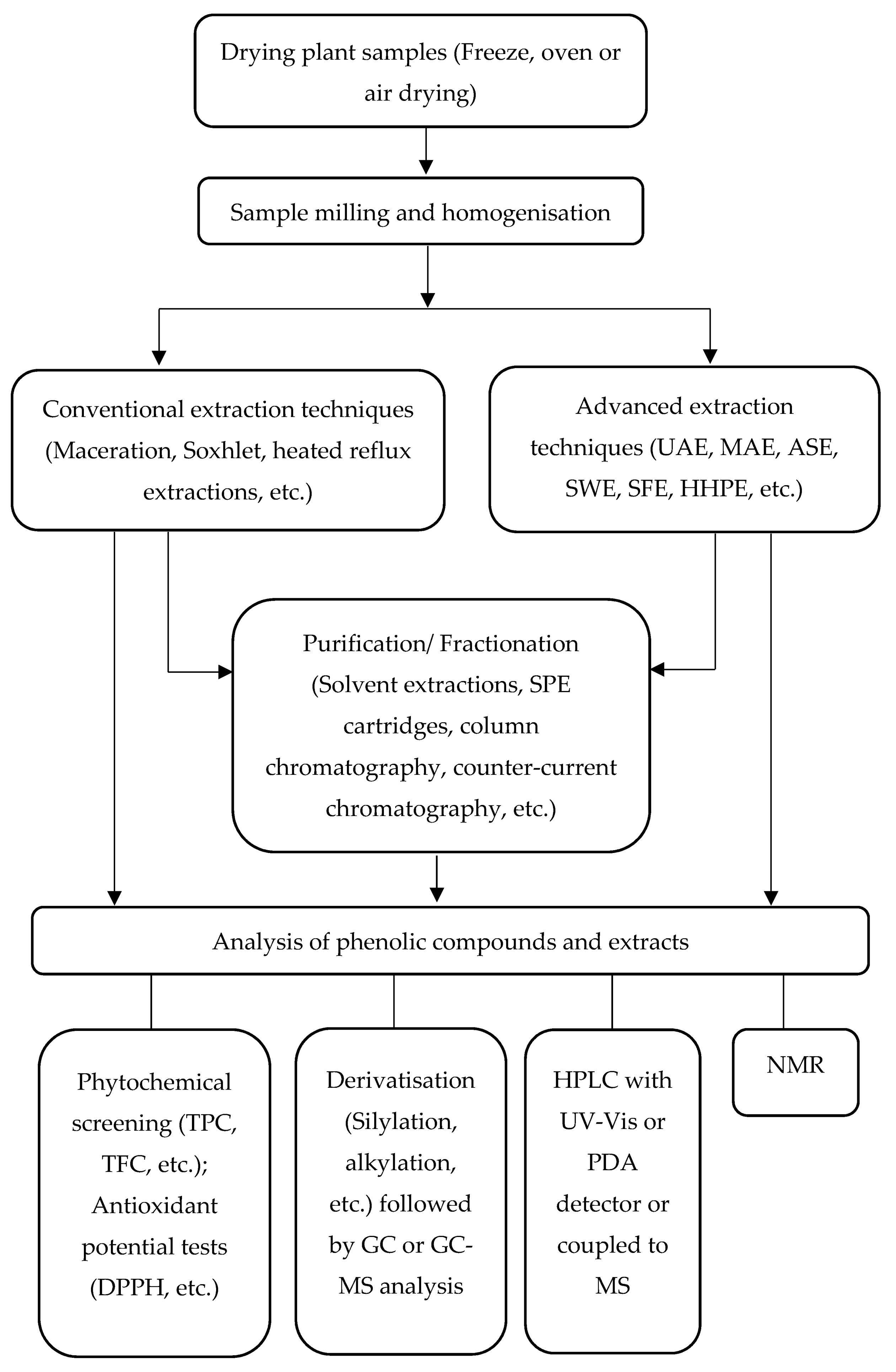
3. Extraction
4. Phytochemical Screening
5. Analytical Methods
5.1. GC, HPLC and Their Hyphenation with MS
5.2. Nuclear Magnetic Resonance Spectroscopy, NMR
5.2.1. 1D 1H NMR
5.2.2. 2D 1H-1H COSY
5.2.3. 2D 1H-1H TOCSY
5.2.4. 2D 1H-1H NOESY
5.2.5. 2D 1H-13C HSQC
5.2.6. 2D 1H-13C HMBC
6. Metabolomic Analysis versus Conventional Natural Product Analysis
7. Potential for Use of Medicinal and Aromatic Plants as Biostimulants and Bioprotectants in Agriculture
7.1. Biostimulant Potential
7.1.1. Regulation of Phytohormone Activity
7.1.2. Modulation of Enzymes
7.1.3. Enhancement of Useful Metabolites
7.2. Bioprotectant Potential
7.2.1. Antimicrobial Tests
7.2.2. Insecticidal Activity
7.2.3. Nematicidal Activity
7.2.4. Herbicidal Activity
8. Conclusions and Suggestions for Future Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Madende, M.; Hayes, M. Fish by-product use as biostimulants: An overview of the current state of the art, including relevant legislation and regulations within the EU and USA. Molecules 2020, 25, 1122. [Google Scholar] [CrossRef] [Green Version]
- Rafiee, H.; Naghdi Badi, H.; Mehrafarin, A.; Qaderi, A.; Zarinpanjeh, N.; Sekara, A.; Zand, E. Application of plant biostimulants as new approach to improve the biological responses of medicinal plants—A critical review. J. Med. Plants 2016, 15, 6–39. [Google Scholar]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Lengai, G.M.W.; Muthomi, J.W.; Mbega, E.R. Phytochemical activity and role of botanical pesticides in pest management for sustainable agricultural crop production. Sci. Afr. 2020, 7, e00239. [Google Scholar] [CrossRef]
- Hikal, W.M.; Baeshen, R.S.; Said-Al Ahl, H.A.H. Botanical insecticide as simple extractives for pest control. Cogent Biol. 2017, 3, 1404274. [Google Scholar] [CrossRef]
- Hussein, R.A.; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants. In Herbal Medicine; Builders, P.F., Ed.; IntechOpen: London, UK, 2018; pp. 11–30. [Google Scholar]
- Jain, C.; Khatana, S.; Vijayvergia, R. Bioactivity of secondary metabolites of various plants: A Review. Int. J. Pharm. Sci. Res. 2019, 10, 494–504. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S.B. Defense potential of secondary metabolites in medicinal plants under UV-B stress. J. Photochem. Photobiol. B Biol. 2019, 193, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Kerala, India, 2006; Volume 661, pp. 23–67. [Google Scholar]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The roles of plant phenolics in defence and communication during Agrobacterium and Rhizobium infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubió, L.; Motilva, M.J.; Romero, M.P. Recent Advances in Biologically Active Compounds in Herbs and Spices: A Review of the Most Effective Antioxidant and Anti-Inflammatory Active Principles. Crit. Rev. Food Sci. Nutr. 2013, 53, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Aromatic plants as a source of bioactive compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M. The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. Sativa) plants. Int. J. Plant Physiol. Biochem. 2013, 5, 42–49. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Tinti, A.; Nardi, S. Biological Activity of Vegetal Extracts Containing Phenols on Plant Metabolism. Molecules 2016, 21, 205. [Google Scholar] [CrossRef]
- Czerniewicz, P.; Chrzanowski, G.; Sytykiewicz, H.; Sprawka, I.; Leszczynski, B. Aphidicidal and deterrent activity of phenolic acid extracts from some herbal plants towards Myzus persicae Sulz. and Rhopalosiphum padi L. Fresenius Environ. Bull. 2016, 25, 5714–5721. [Google Scholar]
- Laquale, S.; Avato, P.; Argentieri, M.P.; Candido, V.; Perniola, M.; D’Addabbo, T. Nematicidal activity of Echinacea species on the root-knot nematode Meloidogyne incognita. J. Pest Sci. 2020, 93, 1397–1410. [Google Scholar] [CrossRef]
- Sodaeizadeh, H.; Rafieiolhossaini, M.; Havlík, J.; van Damme, P. Allelopathic activity of different plant parts of Peganum harmala L. and identification of their growth inhibitors substances. Plant Growth Regul. 2009, 59, 227–236. [Google Scholar] [CrossRef]
- Wink, M. Modes of Action of Herbal Medicines and Plant Secondary Metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef]
- De la Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Chapter 12—Phenolic Compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 253–271. [Google Scholar]
- Teponno, R.B.; Kusari, S.; Spiteller, M. Recent advances in research on lignans and neolignans. Nat. Prod. Rep. 2016, 33, 1044–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Manguro, L.O.A.; Lemmen, P. Phenolics of Moringa oleifera leaves. Nat. Prod. Res. 2007, 21, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.Z.; Sun, M.; Corke, H. Antioxidant capacity of 26 spice extracts and characterization of their phenolic constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar] [CrossRef] [PubMed]
- Koksal, E.; Bursal, E.; Gulcin, I.; Korkmaz, M.; Caglayan, C.; Goren, A.C.; Alwasel, S.H. Antioxidant activity and polyphenol content of Turkish thyme (Thymus vulgaris) monitored by liquid chromatography and tandem mass spectrometry. Int. J. Food Prop. 2017, 20, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Vastano, B.C.; Chen, Y.; Zhu, N.Q.; Ho, C.T.; Zhou, Z.Y.; Rosen, R.T. Isolation and identification of stilbenes in two varieties of Polygonum cuspidatum. J. Agric. Food 2000, 48, 253–256. [Google Scholar] [CrossRef]
- Zhao, X.H.; Han, F.; Li, Y.L.; Yue, H.L. Preparative Isolation and Purification of Three Stilbene Glycosides from the Tibetan Medicinal Plant Rheum tanguticum Maxim. Ex Balf. by High-speed Counter-current Chromatography. Phytochem. Anal. 2013, 24, 171–175. [Google Scholar] [CrossRef]
- Thuong, P.T.; Hung, T.M.; Ngoc, T.M.; Ha, D.T.; Min, B.S.; Kwack, S.J.; Kang, T.S.; Choi, J.S.; Bae, K. Antioxidant Activities of Coumarins from Korean Medicinal Plants and their Structure-Activity Relationships. Phytother. Res. 2010, 24, 101–106. [Google Scholar] [CrossRef]
- Ahmad, A.; Misra, L.N. Isolation of herniarin and other constituents from Matricaria chamomilla flowers. Int. J. Pharmacogn. 1997, 35, 121–125. [Google Scholar] [CrossRef]
- Morikawa, T.; Luo, F.L.; Manse, Y.; Sugita, H.; Saeki, S.; Chaipech, S.; Pongpiriyadacha, Y.; Muraoka, O.; Ninomiya, K. Geranylated Coumarins From Thai Medicinal Plant Mammea siamensis With Testosterone 5 alpha-Reductase Inhibitory Activity. Front. Chem. 2020, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Tiwari, N.; Shanker, K.; Verma, R.K.; Gupta, A.K.; Gupta, M.M. Two new lignans from Phyllanthus amarus. J. Asian Nat. Prod. Res. 2009, 11, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Yang, Z.Y.; Yao, X.C.; Wang, H.; Han, N.; Liu, Z.H.; Wang, Y.; Yang, J.Y.; Yin, J. Dibenzocyclooctadiene lignans from Schisandra chinensis and their inhibitory activity on NO production in lipopolysaccharide-activated microglia cells. Phytochemistry 2014, 104, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Baranyai, B.; Bäcker, C.; Reich, C.; Lindequist, U. The production of 7-methyljuglone, plumbagin and quercetin in wild and cultivated Drosera rotundifolia and Drosera intermedia. Mires Peat 2016, 18, 1–8. [Google Scholar] [CrossRef]
- Tian, K.; Zhang, H.G.; Chen, X.G.; Hu, Z.D. Determination of five anthraquinones in medicinal plants by capillary zone electrophoresis with beta-cyclodextrin addition. J. Chromatogr. A 2006, 1123, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Haque, A.; Saleem, K. Separation and identification of curcuminoids in turmeric powder by HPLC using phenyl column. Anal. Methods 2014, 6, 2526–2536. [Google Scholar] [CrossRef]
- Taamalli, A.; Arraez-Roman, D.; Abaza, L.; Iswaldi, I.; Fernandez-Gutierrez, A.; Zarrouk, M.; Segura-Carretero, A. LC-MS-based metabolite profiling of methanolic extracts from the medicinal and aromatic species Mentha pulegium and Origanum majorana. Phytochem. Anal. 2015, 26, 320–330. [Google Scholar] [CrossRef]
- Rodrigues, F.; Almeida, I.; Sarmento, B.; Amaral, M.H.; Oliveira, M. Study of the isoflavone content of different extracts of Medicago spp. as potential active ingredient. Ind. Crops Prod. 2014, 57, 110–115. [Google Scholar] [CrossRef]
- Stevenson, P.C.; Kite, G.C.; Lewis, G.P.; Forest, F.; Nyirenda, S.P.; Belmain, S.R.; Sileshi, G.W.; Veitch, N.C. Distinct chemotypes of Tephrosia vogelii and implications for their use in pest control and soil enrichment. Phytochemistry 2012, 78, 135–146. [Google Scholar] [CrossRef]
- Russo, M.; Fanali, C.; Tripodo, G.; Dugo, P.; Muleo, R.; Dugo, L.; De Gara, L.; Mondello, L. Analysis of phenolic compounds in different parts of pomegranate (Punica granatum) fruit by HPLC-PDA-ESI/MS and evaluation of their antioxidant activity: Application to different Italian varieties. Anal. Bioanal. Chem. 2018, 410, 3507–3520. [Google Scholar] [CrossRef]
- Niesen, D.B.; Hessler, C.; Seeram, N.P. Beyond resveratrol: A review of natural stilbenoids identified from 2009-2013. J. Berry Res. 2013, 3, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on Natural Coumarin Lead Compounds for Their Pharmacological Activity. BioMed Res. Int. 2013, 14. [Google Scholar] [CrossRef] [Green Version]
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An Important Class of Phytochemicals. In Phytochemicals—Isolation, Characterisation and Role in Human Health; Rao, A.V., Rao, L.G., Eds.; IntechOpen: London, UK, 2015; pp. 113–140. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Garcia, C.; Sanchez-Quesada, C.; Toledo, E.; Delgado-Rodriguez, M.; Gaforio, J.J. Naturally Lignan-Rich Foods: A Dietary Tool for Health Promotion? Molecules 2019, 24, 917. [Google Scholar] [CrossRef] [Green Version]
- Ochwang’I, D.O.; Kimwele, C.N.; Oduma, J.A.; Gathumbi, P.K.; Kiama, S.G.; Efferth, T. Phytochemical Screening of Medicinal Plants of the Kakamega Country, Kenya Commonly Used against Cancer. Med. Aromat. Plants 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Eyong, K.O.; Kuete, V.; Efferth, T. Quinones and Benzophenones from the Medicinal Plants of Africa. In Medicinal Plant Research in Africa; Kuete, V., Ed.; Elsevier: Oxford, UK, 2013; pp. 351–391. [Google Scholar]
- Babula, P.; Adam, V.; Havel, L.; Kizek, R. Noteworthy Secondary Metabolites Naphthoquinones—Their Occurrence, Pharmacological Properties and Analysis. Curr. Pharm. Anal. 2009, 5, 47–68. [Google Scholar] [CrossRef]
- Itokawa, H.; Shi, Q.; Akiyama, T.; Morris-Natschke, S.L.; Lee, K.H. Recent advances in the investigation of curcuminoids. Chin. Med. 2008, 3, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amalraj, A.; Pius, A.; Gopi, S. Biological activities of curcuminoids, other biomolecules from turmeric and their derivatives—A review. J. Tradit. Complementary Med. 2017, 7, 205–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, E.L.; Maia, B.H.L.N.S.; Ferriani, A.P.; Teixeira, S.D. Flavonoids: Classification, Biosynthesis and Chemical Ecology. In Flavonoid—From Biosynthesis to Human Health; Justino, G., Ed.; IntechOpen: London, UK, 2017; pp. 3–16. [Google Scholar]
- Fraga-Corral, M.; Garcia-Oliveira, P.; Pereira, A.G.; Lourenco-Lopes, C.; Jimenez-Lopez, C.; Prieto, M.A.; Simal-Gandara, J. Technological Application of Tannin-Based Extracts. Molecules 2020, 25, 614. [Google Scholar] [CrossRef] [Green Version]
- Bele, A.A.; Jadhav, V.M.; Kadam, V.J. Potential of Tannnins: A Review. Asian J. Plant Sci. 2010, 9, 209–214. [Google Scholar] [CrossRef]
- Chang, Z.; Zhang, Q.; Liang, W.; Zhou, K.; Jian, P.; She, G.; Zhang, L. A Comprehensive Review of the Structure Elucidation of Tannins from Terminalia Linn. Evid. Based Complementary Altern. Med. 2019, 2019, 8623909. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Khoddami, A.; Wilkes, M.A.; Roberts, T.H. Techniques for Analysis of Plant Phenolic Compounds. Molecules 2013, 18, 2328–2375. [Google Scholar] [CrossRef]
- Mushtaq, M.Y.; Choi, Y.H.; Verpoorte, R.; Wilson, E.G. Extraction for Metabolomics: Access to The Metabolome. Phytochem. Anal. 2014, 25, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Orphanides, A.; Goulas, V.; Gekas, V. Effect of drying method on the phenolic content and antioxidant capacity of spearmint. Czech J. Food Sci. 2013, 31, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Hameed, S.; Imran, A.; Nisa, M.U.; Arshad, M.S.; Saeed, F.; Arshad, M.U.; Khan, M.A. Characterization of extracted phenolics from black cumin (Nigella sativa Linn), coriander seed (Coriandrum sativum L.), and fenugreek seed (Trigonella foenum-graecum). Int. J. Food Prop. 2019, 22, 714–726. [Google Scholar] [CrossRef] [Green Version]
- Kouassi, E.K.; Coulibaly, I.; Rodica, P.; Pintea, A.; Ouattara, S.; Odagiu, A. HPLC Phenolic Compounds Analysis and Antifungal Activity of extracts from Cymbopogon citratus (DC) Stapf. against Fusarium graminearum and Fusarium oxysporum sp tulipae. J. Sci. Res. Rep. 2017, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rebey, I.B.; Bourgou, S.; Debez, I.B.; Karoui, I.J.; Sellami, I.H.; Msaada, K.; Limam, F.; Marzouk, B. Effects of Extraction Solvents and Provenances on Phenolic Contents and Antioxidant Activities of Cumin (Cuminum cyminum L.) Seeds. Food Bioprocess Technol. 2012, 5, 2827–2836. [Google Scholar] [CrossRef]
- Gull, T.; Sultana, B.; Anwar, F.; Nouman, W.; Mehmood, T.; Sher, M. Characterization of phenolics in different parts of selected Capparis species harvested in low and high rainfall season. J. Food Meas. Charact. 2018, 12, 1539–1547. [Google Scholar] [CrossRef]
- Oreopoulou, A.; Tsimogiannis, D.; Oreopoulou, V. Extraction of Polyphenols From Aromatic and Medicinal Plants: An Overview of the Methods and the Effect of Extraction Parameters. In Polyphenols in Plants, 2nd ed.; Watson, R.R., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 243–259. [Google Scholar]
- Arceusz, A.; Wesolowski, M.; Konieczynski, P. Methods for Extraction and Determination of Phenolic Acids in Medicinal Plants: A Review. Nat. Prod. Commun. 2013, 8, 1821–1829. [Google Scholar] [CrossRef] [Green Version]
- Azwanida, N.N. Review on the Extraction Methods Use in Medicinal Plants, Principle, Strength and Limitation. Med. Aromat. Plants 2015, 4. [Google Scholar] [CrossRef]
- Cong-Cong, X.; Bing, W.; Yi-Qiong, P.; Jian-Sheng, T.; Tong, Z. Advances in extraction and analysis of phenolic compounds from plant materials. Chin. J. Nat. Med. 2017, 15, 721–731. [Google Scholar]
- Baldino, L.; Scognamiglio, M.; Reverchon, E. Supercritical fluid technologies applied to the extraction of compounds of industrial interest from Cannabis sativa L. and to their pharmaceutical formulations: A review. J. Supercrit. Fluids 2020, 165, 104960. [Google Scholar] [CrossRef]
- de Souza, C.G.; Rodrigues, T.H.S.; e Silva, L.M.A.; Ribeiro, P.R.V.; de Brito, E.S. Sequential extraction of flavonoids and pectin from yellow passion fruit rind using pressurized solvent or ultrasound. J. Sci. Food Agric. 2018, 98, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Garmus, T.T.; Paviani, L.C.; Queiroga, C.L.; Cabral, F.A. Extraction of phenolic compounds from pepper-rosmarin (Lippia sidoides Cham.) leaves by sequential extraction in fixed bed extractor using supercritical CO2, ethanol and water as solvents. J. Supercrit. Fluids 2015, 99, 68–75. [Google Scholar] [CrossRef]
- Joshi, N.; Bhatt, S.; Dyani, S.; Nain, J. Phytochemical screening of secondary metabolites of Argemone mexicana Linn. Flowers. Int. J. Curr. Pharm. Res. 2013, 5, 144–147. [Google Scholar]
- Patel, M.; Verma, R.; Srivastav, P. Antioxidant activity of Eclipta alba extract. J. Med. Plants Stud. 2016, 4, 92–98. [Google Scholar]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef]
- Nitiema, L.W.; Sombie, P.A.E.D.; Koala, M.; Fiore, A.D. Phytochemical Composition and Antioxidant Activity of Balanites aegyptiaca, Securidaca longepedunculata and Acacia gourmaensis Used against Seed-borne Fungi in Burkina Faso. Curr. J. Appl. Sci. Technol. 2020, 39, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Rana, Z.H.; Alam, M.K.; Akhtaruzzaman, M. Nutritional Composition, Total Phenolic Content, Antioxidant and alpha-Amylase Inhibitory Activities of Different Fractions of Selected Wild Edible Plants. Antioxidants 2019, 8, 203. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Lopez, R.I.; Leon-Felix, J.; Angulo-Escalante, M.A.; Gutierrez-Dorado, R.; Muy-Rangel, M.D.; Heredia, J.B. Nutritional and Phenolic Characterization of Moringa oleifera leaves grown in Sinaloa, Mexico. Pak. J. Bot. 2017, 49, 161–168. [Google Scholar]
- Khlifi, D.; Sghaier, R.M.; Amouri, S.; Laouini, D.; Hamdi, M.; Bouajila, J. Composition and anti-oxidant, anti-cancer and anti-inflammatory activities of Artemisia herba-alba, Ruta chalpensis L. and Peganum harmala L. Food Chem. Toxicol. 2013, 55, 202–208. [Google Scholar] [CrossRef]
- Rohloff, J. Analysis of Phenolic and Cyclic Compounds in Plants Using Derivatization Techniques in Combination with GC-MS-Based Metabolite Profiling. Molecules 2015, 20, 3431–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kafkas, N.E.; Kosar, M.; Oz, A.T.; Mitchell, A.E. Advanced Analytical Methods for Phenolics in Fruits. J. Food Qual. 2018, 2018, 3836064. [Google Scholar] [CrossRef]
- Proestos, C.; Kapsokefalou, M.; Komaitis, M. Analysis of naturally occurring phenolic compounds in aromatic plants by RP-HPLC and GC-MS after silylation. J. Food Qual. 2008, 31, 402–414. [Google Scholar] [CrossRef] [Green Version]
- Marques, T.R.; Caetano, A.A.; Alves, D.S.; Ramos, V.D.; Simao, A.A.; Carvalho, G.A.; Correa, A.D. Malpighia emarginata DC. bagasse acetone extract: Phenolic compounds and their effect on Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Chil. J. Agric. Res. 2016, 76, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.M.C.; Seo, D.J.; Kim, K.Y.; Park, R.D.; Kim, D.H.; Han, Y.S.; Kim, T.H.; Jung, W.J. Nematicidal activity of 3,4-dihydroxybenzoic acid purified from Terminalia nigrovenulosa bark against Meloidogyne incognita. Microb. Pathog. 2013, 59–60, 52–59. [Google Scholar] [CrossRef]
- Benayad, Z.; Gomez-Cordoves, C.; Es-Safi, N.E. Characterization of Flavonoid Glycosides from Fenugreek (Trigonella foenum-graecum) Crude Seeds by HPLC-DAD-ESI/MS Analysis. Int. J. Mol. Sci. 2014, 15, 20668–20685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amessis-Ouchemoukh, N.; Abu-Reidah, I.M.; Quirantes-Pine, R.; Madani, K.; Segura-Carretero, A. Phytochemical profiling, in vitro evaluation of total phenolic contents and antioxidant properties of Marrubium vulgare (horehound) leaves of plants growing in Algeria. Ind. Crops Prod. 2014, 61, 120–129. [Google Scholar] [CrossRef]
- Oldoni, T.L.C.; Merlin, N.; Karling, M.; Carpes, S.T.; Alencar, S.M.d.; Morales, R.G.F.; Silva, E.A.d.; Pilau, E.J. Bioguided extraction of phenolic compounds and UHPLC-ESI-Q-TOF-MS/MS characterization of extracts of Moringa oleifera leaves collected in Brazil. Food Res. Int. 2019, 125, 108647. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Ø.M.; Fossen, T. Characterization of Anthocyanins by NMR. Curr. Protoc. Food Anal. Chem. 2003, 9, F1.4.1–F1.4.23. [Google Scholar] [CrossRef]
- Hatzakis, E. Nuclear Magnetic Resonance (NMR) Spectroscopy in Food Science: A Comprehensive Review. Compr. Rev. Food. Sci. Food Saf. 2019, 18, 189–220. [Google Scholar] [CrossRef] [Green Version]
- Dona, A.C.; Kyriakides, M.; Scott, F.; Shephard, E.A.; Varshavi, D.; Veselkov, K.; Everett, J.R. A guide to the identification of metabolites in NMR-based metabonomics/metabolomics experiments. Comp. Struct. Biotechnol. J. 2016, 14, 135–153. [Google Scholar] [CrossRef] [Green Version]
- Abedini, A.; Roumy, V.; Mahieux, S.; Biabiany, M.; Standaert-Vitse, A.; Riviere, C.; Sahpaz, S.; Bailleul, F.; Neut, C.; Hennebelle, T. Rosmarinic Acid and Its Methyl Ester as Antimicrobial Components of the Hydromethanolic Extract of Hyptis atrorubens Poit. (Lamiaceae). Evid. Based Complementary Altern. Med. 2013, 2013, 604536. [Google Scholar] [CrossRef] [Green Version]
- Rayyan, S.; Fossen, T.; Andersen, Ø.M. Flavone C-Glycosides from Seeds of Fenugreek, Trigonella foenum-graecum L. J. Agric. Food Chem. 2010, 58, 7211–7217. [Google Scholar] [CrossRef]
- Khalafallah, A.K.; Suleiman, S.A.; Yousef, A.H.; El-kanzi, N.A.A.; Mohamed, A. Prenylated flavonoids from Tephrosia apollinea. Chin. Chem. Lett. 2009, 20, 1465–1468. [Google Scholar] [CrossRef]
- Zhao, H.Y.; Fan, L.; Zhou, L.; Han, J.; Wang, R.R.; Guo, D.A. 8-(3,3-dimethylallyl)-substituted flavonoid glycosides from the aerial parts of Epimedium koreanum. Helv. Chim. Acta 2007, 90, 2186–2195. [Google Scholar] [CrossRef]
- Li, J.; Adelakun, T.A.; Wang, S.J.; Ruan, J.Y.; Yang, S.C.; Li, X.X.; Zhang, Y.; Wang, T. Inhibitory Effects of Constituents from the Aerial Parts of Rosmarinus officinalis L. on Triglyceride Accumulation. Molecules 2017, 22, 110. [Google Scholar] [CrossRef] [Green Version]
- Krzyzanowska-Kowalczyk, J.; Pecio, L.; Moldoch, J.; Ludwiczuk, A.; Kowalczyk, M. Novel Phenolic Constituents of Pulmonaria officinalis L. LC-MS/MS Comparison of Spring and Autumn Metabolite Profiles. Molecules 2018, 23, 2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akoury, E. Isolation and structural elucidation of rosmarinic acid by nuclear magnetic resonance spectroscopy. Am. Res. J. Chem. 2017, 2, 17–23. [Google Scholar] [CrossRef]
- Kabbash, A.; Shoeib, N. Chemical and Biological Investigation of Some Secondary Metabolites in Atriplex halimus Growing in Egypt. Nat. Prod. Commun. 2012, 7, 1465–1468. [Google Scholar] [CrossRef] [Green Version]
- Exarchou, V.; Troganis, A.; Gerothanassis, I.P.; Tsimidou, M.; Boskou, D. Identification and quantification of caffeic and rosmarinic acid in complex plant extracts by the use of variable-temperature two-dimensional nuclear magnetic resonance spectroscopy. J. Agric. Food Chem. 2001, 49, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Oh, D.-G.; Singh, D.; Lee, J.S.; Lee, S.; Lee, C.H. Exploring the metabolomic diversity of plant species across spatial (leaf and stem) components and phylogenic groups. BMC Plant Biol. 2020, 20, 39. [Google Scholar] [CrossRef]
- Demarque, D.P.; Dusi, R.G.; de Sousa, F.D.M.; Grossi, S.M.; Silvério, M.R.S.; Lopes, N.P.; Espindola, L.S. Mass spectrometry-based metabolomics approach in the isolation of bioactive natural products. Sci. Rep. 2020, 10, 1051. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; de Souza, L.P.; Serag, A.; Fernie, A.R.; Farag, M.A.; Ezzat, S.M.; Alseekh, S. Metabolomics in the Context of Plant Natural Products Research: From Sample Preparation to Metabolite Analysis. Metabolites 2020, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.J.; Ran, J.L.; Zhang, A.H.; Sun, H.; Tan, G.L.; Han, Y.; Liu, L. Novel applications of mass spectrometry-based metabolomics in herbal medicines and its active ingredients: Current evidence. Mass Spectrom. Rev. 2019, 38, 380–402. [Google Scholar] [CrossRef]
- Leiss, K.A.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. An overview of NMR-based metabolomics to identify secondary plant compounds involved in host plant resistance. Phytochem. Rev. 2011, 10, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Bentley, J.; Moore, J.P.; Farrant, J.M. Metabolomic Profiling of the Desiccation-Tolerant Medicinal Shrub Myrothamnus flabellifolia Indicates Phenolic Variability Across Its Natural Habitat: Implications for Tea and Cosmetics Production. Molecules 2019, 24, 1240. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.A.; Ali, S.E.; Hodaya, R.H.; El-Seedi, H.R.; Sultani, H.N.; Laub, A.; Eissa, T.E.; Abou-Zaid, F.O.F.; Wessjohann, L.A. Phytochemical Profiles and Antimicrobial Activities of Allium cepa Red cv. and A. sativum Subjected to Different Drying Methods: A Comparative MS-Based Metabolomics. Molecules 2017, 22, 761. [Google Scholar] [CrossRef]
- Wu, K.X.; Liu, J.; Liu, Y.; Guo, X.R.; Mu, L.Q.; Hu, X.H.; Tang, Z.H. A Comparative Metabolomics Analysis Reveals the Tissue-Specific Phenolic Profiling in Two Acanthopanax Species. Molecules 2018, 23, 2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piasecka, A.; Kachlicki, P.; Stobiecki, M. Analytical Methods for Detection of Plant Metabolomes Changes in Response to Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalla, M.H. Some Phenolic Compounds Enhance Nodulation and Nitrogen Fixation in a Soybean/Bradyrhizobium japonicum System. Phyton-Ann. REI Bot. 1994, 33, 249–256. [Google Scholar]
- Hayat, S.; Ahmad, H.; Ali, M.; Ren, K.L.; Cheng, Z.H. Aqueous garlic extract stimulates growth and antioxidant enzymes activity of tomato (Solanum lycopersicum). Sci. Hortic. 2018, 240, 139–146. [Google Scholar] [CrossRef]
- Zida, E.P.; Sereme, P.; Leth, V.; Sankara, P. Effect of Aqueous Extracts of Acacia gourmaensis A. Chev and Eclipta alba (L.) Hassk. on Seed Health, Seedling Vigour and Grain Yield of Sorghum and Pearl Millet. Asian J. Plant Pathol. 2008, 2, 40–47. [Google Scholar] [CrossRef]
- Thanaa, S.h.M.; Kassim, N.E.; AbouRayya, M.S.; Abdalla, A.M. Influence of Foliar Application with Moringa (Moringa oleifera L.) Leaf Extract on Yield and Fruit Quality of Hollywood Plum Cultivar. J. Hortic. 2017, 4, 1–7. [Google Scholar] [CrossRef]
- Mkindi, A.G.; Tembo, Y.L.B.; Mbega, E.R.; Smith, A.K.; Farrell, I.W.; Ndakidemi, P.A.; Stevenson, P.C.; Belmain, S.R. Extracts of Common Pesticidal Plants Increase Plant Growth and Yield in Common Bean Plants. Plants 2020, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayange, C.D.M.; Njera, D.; Nyirenda, S.P.; Mwamlima, L. Effectiveness of Tephrosia vogelii and Tephrosia candida Extracts against Common Bean Aphid (Aphis fabae) in Malawi. Adv. Agric. 2019, 2019, 6704834. [Google Scholar] [CrossRef] [Green Version]
- Alao, F.O.; Adebayo, T.A. Comparative efficacy of Tephrosia vogelii and Moringa oleifera against insect pests of watermelon (Citrullus lanatus Thumb.). Int. Lett. Nat. Sci. 2015, 35, 71–78. [Google Scholar]
- Ahmed, M.F.; Khalequzzaman, K.M.; Islam, M.N.; Anam, M.K.; Islam, M.T. Effect of Plant Extracts Against Bipolaris oryzae of Rice under In vitro Conditions. Pak. J. Biol. Sci. 2002, 5, 442–445. [Google Scholar] [CrossRef]
- Odewole, A.F.; Adebayo, T.A.; Babarinde, S.A.; Awolokun, G.S. Insecticidal activity of aqueous indigenous plant extracts against insect pests associated with cucumber (Cucumis sativus L.) in Southern Guinea Savannah Zone of Nigeria. Arch. Phytopathol. Plant Prot. 2020, 53, 230–246. [Google Scholar] [CrossRef]
- Zida, P.E.; Neya, B.J.; Soalla, R.W.; Sereme, P.; Lund, O.S. Seed treatment with Eclipta alba plant extract and application of fertilizer confers a combined effect on the yield of sorghum. Int. J. Agron. Agric. Res. 2018, 13, 96–102. [Google Scholar]
- De Klerk, G.J.; Guan, H.Y.; Huisman, P.; Marinova, S. Effects of phenolic compounds on adventitious root formation and oxidative decarboxylation of applied indoleacetic acid in Malus ‘Jork 9’. Plant Growth Regul. 2011, 63, 175–185. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.A.T.; Dobranszki, J.; Ross, S. Phloroglucinol in plant tissue culture. In Vitro Cell. Dev. Biol.-Plant 2013, 49, 1–16. [Google Scholar] [CrossRef]
- Ranade, S.; David, S.B. Quinones As Plant-Growth Regulators. Plant Growth Regul. 1985, 3, 3–13. [Google Scholar] [CrossRef]
- Wuyts, N.; Swennen, R.; De Waele, D. Effects of plant phenylpropanoid pathway products and selected terpenoids and alkaloids on the behaviour of the plant-parasitic nematodes Radopholus similis, Pratylenchus penetrans and Meloidogyne incognita. Nematology 2006, 8, 89–101. [Google Scholar] [CrossRef]
- Cetin-Karaca, H.; Newman, M.C. Antimicrobial Efficacy of Natural Phenolic Compounds against Gram Positive Foodborne Pathogens. J. Food Res. 2015, 4, 14. [Google Scholar] [CrossRef]
- Maddox, C.E.; Laur, L.M.; Tian, L. Antibacterial Activity of Phenolic Compounds Against the Phytopathogen Xylella fastidiosa. Curr. Microbiol. 2010, 60, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, D.C.; Wearing, A.H.; Coates, L.; Terry, L. Effects of phosphonate and salicylic acid treatments on anthracnose disease development and ripening of ‘Kensington Pride’ mango fruit. Aust. J. Exp. Agric. 2001, 41, 805–813. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J.; Al-Dakheel, A.J. Biochemical, physiological and isotopic responses to natural product p-hydroxybenzoic acid in Cocksfoot (Dactylis glomerata L.). Plant Growth Regul. 2015, 75, 783–792. [Google Scholar] [CrossRef]
- Mahajan, R.; Kaur, D.J.; Bajaj, K.L. Nematicidal Activity of Phenolic Compounds against Meloidogyne incognita. Nematol. Mediterr. 1992, 20, 217–219. [Google Scholar]
- Mahajan, R.; Singh, P.; Bajaj, K. L Nematicidal activity of some phenolic compounds against Meloidogyne incognita. Revue Nematol. 1985, 8, 161–164. [Google Scholar]
- Chong, K.P.; Rossall, S.; Atong, M. In Vitro Antimicrobial Activity and Fungitoxicity of Syringic Acid, Caffeic Acid and 4-hydroxybenzoic Acid against Ganoderma boninense. J. Agric. Sci. 2009, 1, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, M. Inhibitory effects of spice extracts on the growth of Aspergillus parasiticus NRRL2999 strain. Z. Lebensm. Unters. Forsch. A Food Res. Technol. 1998, 207, 253–255. [Google Scholar] [CrossRef]
- Fadel, F.; Ben Hmamou, D.; Salghi, R.; Chebli, B.; Benali, O.; Zarrouk, A.; Ebenso, E.E.; Chakir, A.; Hammouti, B. Antifungal Activity and Anti-Corrosion Inhibition of Origanum compactum Extracts. Int. J. Electrochem. Sci. 2013, 8, 11019–11032. [Google Scholar]
- Srivastava, A.K.; Lal, B. Studies on biofungicidal properties of leaf extract of some plants. Indian Phytopathol. 1997, 50, 408–411. [Google Scholar]
- Ahmad, F.; Rather, M.A.; Siddiqui, M.A. Nematicidal Activity of Leaf Extracts from Lantana camara L. against Meloidogyne incognita (Kofoid and White) Chitwood and its use to Manage Roots Infection of Solanum melongena L. Braz. Arch. Biol. Technol. 2010, 53, 543–548. [Google Scholar] [CrossRef]
- Jyotsna, J.; Das, S.; Kumar, B. Efficacy of Aqueous Leaf Extract of Medicinal Plants against Blast and Brown Spot Disease of Rice. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 4138–4144. [Google Scholar] [CrossRef]
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar] [CrossRef] [PubMed]
- Terry, L.A.; Joyce, D.C. Elicitors of induced disease resistance in postharvest horticultural crops: A brief review. Postharvest Biol. Technol. 2004, 32, 1–13. [Google Scholar] [CrossRef]
- Taguri, T.; Tanaka, T.; Kouno, I. Antibacterial spectrum of plant polyphenols and extracts depending upon hydroxyphenyl structure. Biol. Pharm. Bull. 2006, 29, 2226–2235. [Google Scholar] [CrossRef] [Green Version]
- Taguri, T.; Tanaka, T.; Kouno, I. Antimicrobial activity of 10 different plant polyphenols against bacteria causing food-borne disease. Biol. Pharm. Bull. 2004, 27, 1965–1969. [Google Scholar] [CrossRef] [Green Version]
- Tomczyk, A.; Suszko, M. The role of phenols in the influence of herbal extracts from Salvia officinalis L. and Matricaria chamomilla L. on two-spotted spider mite Tetranychus urticae Koch. Biol. Lett. 2011, 48, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Castillo, L.; Gonzalez-Coloma, A.; Gonzalez, A.; Diaz, M.; Santos, E.; Alonso-Paz, E.; Bassagoda, M.J.; Rossini, C. Screening of Uruguayan plants for deterrent activity against insects. Ind. Crops Prod. 2009, 29, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Gupta, M.M.; Prajapati, V.; Tripathi, A.K.; Kumar, S. Sesamin a potent antifeedant principle from Piper mullesua. Phytother. Res. 2001, 15, 70–72. [Google Scholar] [CrossRef]
- Ohri, P.; Pannu, S.K. Effect of phenolic compounds on nematodes- A review. J. Appl. Nat. Sci. 2010, 2, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Asif, M.; Tariq, M.; Rehman, B.; Parihar, K.; Siddiqui, M.A. Phytochemical Investigation, Nematostatic and Nematicidal Potential of Weeds Extract Against the Root-knot Nematode, Meloidogyne incognita In Vitro. Asian J. Biol. Sci. 2017, 10, 38–46. [Google Scholar] [CrossRef]
- Korayem, A.M.; Hasabo, S.A.; Ameen, H.H. Effects and mode of action of some plant extracts on certain plant parasitic nematodes. Anz. Schadlingskd. Pflanzenschutz Umweltschutz 1993, 66, 32–36. [Google Scholar] [CrossRef]
- Sultana, N.; Akhter, M.; Khatoon, Z. Nematicidal natural products from the aerial parts of Rubus niveus. Nat. Prod. Res. 2010, 24, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Wang, Q.A.; Ruan, X.A.; Pan, C.D.; Jiang, D.A. Phenolics and Plant Allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodaeizadeh, H.; Rafieiolhossaini, M.; Van Damme, P. Herbicidal activity of a medicinal plant, Peganum harmala L., and decomposition dynamics of its phytotoxins in the soil. Ind. Crops Prod. 2010, 31, 385–394. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Phenolic Compound | Phenolic Subcategory | Medicinal/Aromatic Plant | Reference |
|---|---|---|---|
| Phenolic acids and derivatives | |||
| Syringic acid, gallic acid | Hydroxybenzoic acid | Moringa oleifera | [27] |
| Gallic acid, vanillic acid 3,4-dihydroxybenzoic acid, 4-hydroxybenzoic acid, syringic acid | Peganum harmala | [22] | |
| Caffeic acid, p-coumaric acid | Hydroxycinnamic acid | Origanum vulgare L., Salvia officinalis L., Thymus vulgaris L., Ocimum basilicum L. | [28] |
| Vanillin | Hydroxybenzoic acid derivative | T. vulgaris | [29] |
| Rosmarinic acid | Hydroxycinnamic acid derivative | Rosmarinus officinalis L., Mentha canadensis L. | [28] |
| Stilbenes | |||
| Piceatannol glucoside, resveratroloside, piceid | Stilbene glycosides | Polygonum cuspidatum | [30] |
| Trans-rhapontin, cis-rhapontin and trans-desoxyrhaponticin | Stilbene glycosides | Rheum tanguticum Maxim. ex Balf. | [31] |
| Coumarins | |||
| Scopoletin, fraxetin, aesculetin, fraxin, aesculin | Coumarins, coumarin glycosides | Fraxinus rhynchophylla. | [32] |
| Herniarin | Coumarin | Matricaria chamomilla | [33] |
| Kayeassamin I, mammeasin E, mammeasin E | Geranylated coumarins | Mammea siamensis | [34] |
| Lignans | |||
| Phyllanthin, hypophyllanthin, niranthin, nirtetralin, 5-demethoxy niranthin, virgatusin, heliobuphthalmin lactone and bursehernin | Lignans | Phyllanthus amarus | [35] |
| Schisanchinin A, schisanchinin B, schisanchinin C, schisanchinin D | Dibenzocyclooctadiene lignans | Schisandra chinensis | [36] |
| Quinones | |||
| 7-methlyjuglone | Naphthoquinone | Drosera rotundifolia | [37] |
| Aloe-emodin, emodin, chrysophanol, physcoin and rhein | Anthraquinones | Rheumpalmatum and Rheum hotaoense | [38] |
| Curcuminoids | |||
| Curcumin, demethoxycurcumin and bis-demethoxycurcumin | Curcuminoids | Curcuma longa | [39] |
| Flavonoids | |||
| Luteolin, apigenin, apigenin-O-glucuronide, luteolin-O-glycoside, orientin, apigenin-6,8-di-C-hexoside | Flavones and flavone glycosides | Origanum majorana | [40] |
| Daidzein, glycitein, genistein, formononetin, prunetin, biochanin A and daidzin, genistin | Isoflavone aglycones and isoflavone glycosides | Medicago spp. | [41] |
| Kaempferol 3-O-glucoside, isorhamnetin 3-O-galactoside | Flavonol glycosides | Tephrosia vogelii | [42] |
| Kaempferol 3-O-rhamnoside, rutin and quercetin 3-O-glucoside | M.oleifera | [27] | |
| Naringenin-O-rhamnoglucoside, hesperidin, isosakuranetin-O-rutinoside | Flavanone glycosides | Mentha pulegium | [40] |
| Taxifolin, taxifolin methyl ether, dihydrokaempferide | Flavanonols | O. majorana | [40] |
| Gallocatechin, catechin | Flavan-3-ols | M. pulegium | [40] |
| Tannins | |||
| Granatin A, punicalagin, peduncalagin I, ellagic acid, ellagic acid glucoside, ellagic acid pentoside, punigluconin | Ellagitannins | Punica granatum | [43] |
| Medicinal/Aromatic Plant | Common Name | Plant Part | Solvent | Composition | Reference | |
|---|---|---|---|---|---|---|
| TPC | TFC | |||||
| Acacia gourmaensis 1 | Stem bark | 70% Ethanol | 807.58 | 271.39 | [75] | |
| Eclipta alba 1 | False daisy | Leaves | 42.5 mL Methanol + 7.5 mL 1N HCl | 55.32 | 31.55 | [76] |
| Roots | 8.45 | 4.88 | ||||
| Whole plant | 50% Ethanol | 98.39 | 86.53 | [73] | ||
| Moringa oleifera 1 | Moringa | Leaves | Methanol | 76.63 | 60.26 | [77] |
| Capparis spinosa 2 | Caper bush | Stem bark | 80% Methanol | 408.4 | 157.3 | [64] |
| Shoot | 229.2 | 250.8 | ||||
| Fruit | 535.8 | 283.3 | ||||
| Flowers | 416.0 | 235.6 | ||||
| Roots | 531.9 | 348.6 | ||||
| Coriandrum sativum 3 | Coriander | Seeds | 70% Methanol | 289.3 | 179.6 | [61] |
| Water | 250.4 | 162.6 | ||||
| Thymus vulgaris 4 | Common thyme | Leaves | Water | 256 | 44.2 | [29] |
| Ethanol | 158 | 36.3 | ||||
| Cuminum cyminum 5 | Cumin | Seeds (Tunisian) | 80% Acetone | 18.60 | 5.91 | [63] |
| Seeds (Indian) | 14.15 | 4.19 | ||||
| Cymbopogon citratus (DC) Stapf. 1 | Lemongrass | Leaves | Methanol | 118.14 | [62] | |
| Ethanol | 35.43 | |||||
| Mentha pulegium 5 | Pennyroyal | Aerial parts | Methanol | 18.28 | 13.76 | [40] |
| Peganum harmala 6 | African Rue | Seeds | 75% Methanol | 93.39 | 1.60 | [78] |
| Medicinal/Aromatic Plant | Common Names | Plant Part | Application Method | Tested Concentrations | Biostimulated Parameters | Reference |
|---|---|---|---|---|---|---|
| Acacia gourmaensis | Bark | Seed immersion in extract | 25% w/v | Seedling emergence, growth, vigour and weight in sorghum and pearl millet. | [110] | |
| Eclipta alba | False daisy | Whole plant | Seed immersion in extract | 25% w/v | Grain yield, seedling emergence, growth, vigour and weight in pearl millet and sorghum | [110] |
| Moringa oleifera | Moringa, horse-radish tree | Leaves | Foliar spray | 1:32 v/v from 1000% w/v stock solution | Growth and yield in Cauliflower | [76] |
| Leaves | Foliar spray | 4%, 5%, 6% from 10% w/v stock solution | Yield, colour, weight and firmness of plums | [111] | ||
| Leaves and twigs | Foliar spray | 1, 2, 3% leaf and 1, 2, 3% twig extracts | Plant height, fresh herb weight and dry herb weight in rocket | [18] | ||
| Crataegus monogyna Jacq. | Hawthorn | Leaves | 0.1, 1.0 mL/L of commercial extract | Dry weight of both roots and leaves of the maize plants | [19] | |
| Tephrosia vogelii | Vogel’s tephrosia, fish-poison bean | Leaves | Foliar spray and soil drench | 10% w/v | Plant height, number of leaves and branches, leaf area, stem width, leaf greenness in beans | [112] |
| Leaves | Plant spraying | 0.5, 2, 5% w/v | Yield (number of pods, pod length, shelled bean weight and seeds) in beans | [113] | ||
| Leaves | Plant spraying | 5, 10, 20% from 50% w/v stock solution | Yield of watermelon | [114] | ||
| Allium sativum | Garlic | Clove | Seed soaking | 100, 50, 25, 12.5% w/v | Percentage germination as well as root and shoot length of seedlings and plant vigour in rice | [115] |
| Bulbs | Fertigation and foliar application | 50, 100, 200 µg/mL | Plant height, leaf area, stem diameter, fresh and dry weight in tomatoes | [109] | ||
| Zingiber officinale | Ginger | Rhizome | Foliar application | 10 and 15% from 50% w/v stock solution | Fruit yield in cucumber | [116] |
| Phenolic Compound | Solvent | Effective Concentration | Bioprotectant Effect | References |
|---|---|---|---|---|
| Ferulic acid | Water | LC50, 120 µg/mL | Strong repellent, nematistatic and nematicidal activity against Radopholus similis | [121] |
| Curcumin | 95% Ethanol | MIC < 5 ppm | Inhibition of in vitro growth of Clostridium perfringens, Clostridium butyricum and Clostridium sporogenes | [122] |
| Ellagic acid | 95% Ethanol | MIC < 5 ppm | Inhibition of in vitro growth of Listeria monocytogenes | [122] |
| Catechol, coumarin, gallic acid, caffeic acid, p-coumaric acid, ferulic acid | Water | MIC, 100–2000 µM | Inhibition of in vitro growth of Xylella fastidiosa | [123] |
| Catechin, naringenin, quercetin, resveratrol, rutin, sinapic acid | DMSO | MIC, 200–2000 µM | Inhibition of in vitro growth of X. fastidiosa | [123] |
| Salicylic acid | LC50, 46 µg/mL | Attractant, motility and hatch inhibition and nematicidal effects on Meloidogyne incognita | [121] | |
| 2 mg/mL | Reduction of the severity of anthracnose disease caused by Colletotrichum gloeosporioides in mangoes | [124] | ||
| 4-hydroxybenzoic acid | 20% Methanol | 0.1 to 1.5 mM | Herbicidal activity against Dactylis glomerata | [125] |
| Pyrogallol, ethylgallate, gentistic acid, juglone, apigenin 7-O-glucoside and 7-hydroxycoumarin | Water | 1100 ppm | Nematicidal effects on M. incognita | [126,127] |
| Syringic acid | 50% Acetone | 1 mg/mL | Inhibition of radial growth of Ganoderma boninense in vitro | [128] |
| Medicinal/Aromatic Plant | Common Name | Plant Part | Extract Solvent | Effective Concentration | Bioprotectant Effect | References |
|---|---|---|---|---|---|---|
| Acacia gourmaensis | Bark | Water | 25 g/100 mL | Reduced the incidence of Phoma sorghina, Curvularia lunata and Fusarium moniliforme in pearl millet seeds, and Phoma sorghina and Colletotrichum graminicola in sorghum seeds | [110] | |
| Azadirachta indica | Neem tree | Leaf | Water | 100 g/100 mL | Reduced incidence of seed borne Bipolaris oryzae in rice | [115] |
| Cymbopogon citratus (DC) Stapf. | Lemon grass | Leaves | Methanol | MIC, 25 μg/mL | Inhibition of in vitro growth of Fusarium graminearum | [62] |
| MIC, 12.5 μg/mL | Inhibition of in vitro growth of Fusarium oxysporum sp tulipae | |||||
| Thymbra spicata L. | Black thyme, Mediterranean thyme | Leaves and flowers | Methanol | 2% | Complete inhibition of in vitro growth of Aspergillus parasiticus | [129] |
| Origanum compactum | Aerial parts | Water, 80% methanol | 25 g/L | Complete inhibition of radial growth of Penicillium digitatum | [130] | |
| Lantana camara L. | Lantana, tickberry | Leaves | Water | 25 g/100 mL | Complete inhibition of spore germination of Curvularia tuberculata and protected pear fruits from rot caused by the pathogen | [131] |
| 25 g/75 ml | Immobilised root knot nematodes, M. incognita | [132] | ||||
| Ocimum basilicum L. | Basil | Leaves | Water | 25 g/100 mL | Inhibition of spore germination of Alternaria alternata and protected pear fruits from rot caused by the pathogen | [131] |
| Moringa olieifera | Moringa, horse-radish tree | Leaves | Water | 20% v/v | Reduced populations of adult Phyllotreta cruciferae insects in watermelon | [114] |
| Tephrosia vogelii | Vogel’s tephrosia, fish-poison bean | Leaves | Water | 20% v/v | Reduced populations of adult Diabrotica undecimpunctata and Dacus cucurbitea insects in water mellon | [114] |
| 5% w/v | Reduction of aphid infestation in beans | [113] | ||||
| Mentha piperita L. | Peppermint | Leaves and flowers | Water containing 0.05% Tween 80 and 4% ethanol | 0.2% | Feeding deterrent of peach potato aphid, Myzus persicae and the bird-cherry oat aphid, Rhopalosiphum padi L. on pea and winter wheat; decreased fecundity and increased larval development time | [20] |
| Echinacea angustifolia | Narrow-leaf coneflower | Roots, aerial parts | Water | LC50 352 µg/mL, 487 µg/mL | Caused juvenile mortality and inhibition of egg hatching in M. incognita nematodes | [21] |
| Peganum harmala | African Rue | Leaves, stems, roots | Water | Herbicidal activity against Avena fatua and Convolvulus arvensis | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kisiriko, M.; Anastasiadi, M.; Terry, L.A.; Yasri, A.; Beale, M.H.; Ward, J.L. Phenolics from Medicinal and Aromatic Plants: Characterisation and Potential as Biostimulants and Bioprotectants. Molecules 2021, 26, 6343. https://doi.org/10.3390/molecules26216343
Kisiriko M, Anastasiadi M, Terry LA, Yasri A, Beale MH, Ward JL. Phenolics from Medicinal and Aromatic Plants: Characterisation and Potential as Biostimulants and Bioprotectants. Molecules. 2021; 26(21):6343. https://doi.org/10.3390/molecules26216343
Chicago/Turabian StyleKisiriko, Musa, Maria Anastasiadi, Leon Alexander Terry, Abdelaziz Yasri, Michael Henry Beale, and Jane Louise Ward. 2021. "Phenolics from Medicinal and Aromatic Plants: Characterisation and Potential as Biostimulants and Bioprotectants" Molecules 26, no. 21: 6343. https://doi.org/10.3390/molecules26216343
APA StyleKisiriko, M., Anastasiadi, M., Terry, L. A., Yasri, A., Beale, M. H., & Ward, J. L. (2021). Phenolics from Medicinal and Aromatic Plants: Characterisation and Potential as Biostimulants and Bioprotectants. Molecules, 26(21), 6343. https://doi.org/10.3390/molecules26216343