
Zinc as a Drug for Wilson’s Disease, Non-Alcoholic Liver Disease and COVID-19-Related Liver Injury

 ,
, 
Abstract
:1. Introduction
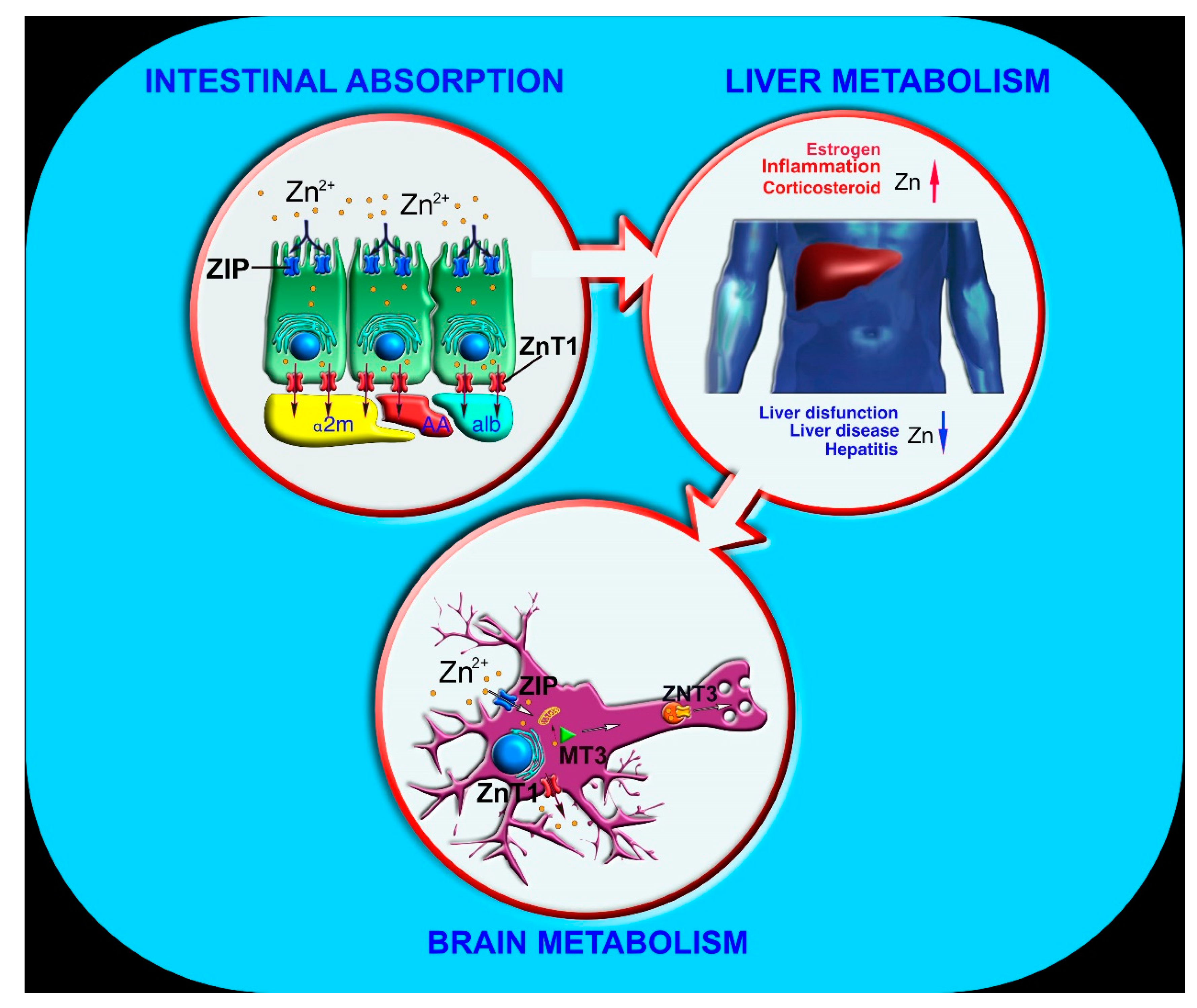
2. Zinc Absorption and Metabolism
3. Zinc Binding Proteins and Zinc Biological Role
4. Zinc Pharmacodynamics and Clinical Trials
5. Zinc in the Therapy of Wilson’s Disease
6. Zinc in the Therapy of Non-Alcoholic Liver Disease (NAFLD)
7. Zinc in the Prevention of COVID-19-Related Liver Injury
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grochowski, C.; Blicharska, E.; Krukow, P.; Jonak, K.; Maciejewski, M.; Szczepanek, D.; Jonak, K.; Flieger, J.; Maciejewski, R. Analysis of trace elements in human brain: Its aim, methods, and concentration levels. Front. Chem. 2019, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Maares, M.; Haase, H. A guide to human zinc absorption: General overview and recent advances of in vitro intestinal models. Nutrients 2020, 12, 762. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.J. Physiology of zinc: General aspects. In Zinc in Human Biology; Mills, C.F., Ed.; Springer: London, UK, 1989; pp. 1–14. [Google Scholar]
- Coni, P.; Ravarino, A.; Farci, A.M.G.; Callea, F.; Van Eyken, P.; Sciot, R.; Ambu, R.; Marras, A.; Costa, V.; Faa, G. Zinc content and distribution in the newborn liver. J. Pediatric Gastroenterol. Nutr. 1996, 23, 125–129. [Google Scholar] [CrossRef]
- Al-Harthi, S.; Lachowicz, J.I.; Nowakowski, M.E.; Jaremko, M.; Jaremko, Ł. Towards the functional high-resolution coordination chemistry of blood plasma human serum albumin. J. Inorg. Biochem. 2019, 198, 110716. [Google Scholar] [CrossRef]
- Livingstone, C. Zinc: Physiology, deficiency, and parenteral nutrition. Nutr. Clin. Pract. 2015, 30, 371–382. [Google Scholar] [CrossRef]
- Leoni, G.; Rosato, A.; Perozzi, G.; Murgia, C. Zinc proteome interaction network as a model to identify nutrient-affected pathways in human pathologies. Genes Nutr. 2014, 9, 436. [Google Scholar] [CrossRef] [Green Version]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef]
- WHO. The World Health Report 2002: Reducing Risks, Promoting Healthy Life; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144. [Google Scholar]
- Wessels, I.; Rink, L. Micronutrients in autoimmune diseases: Possible therapeutic benefits of zinc and vitamin D. J. Nutr. Biochem. 2020, 77, 108240. [Google Scholar] [CrossRef]
- Haase, H.; Schomburg, L. You’d Better Zinc—Trace Element Homeostasis in Infection and Inflammation. Nutrients 2019, 11, 2078. [Google Scholar] [CrossRef] [Green Version]
- Ackland, M.L.; Michalczyk, A. Zinc deficiency and its inherited disorders—A review. Genes Nutr. 2006, 1, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Zamai, L. The Yin and Yang of ACE/ACE2 Pathways: The Rationale for the Use of Renin-Angiotensin System Inhibitors in COVID-19 Patients. Cells 2020, 9, 1704. [Google Scholar] [CrossRef]
- Derwand, R.; Scholz, M. Does zinc supplementation enhance the clinical efficacy of chloroquine/hydroxychloroquine to win today’s battle against COVID-19? Med. Hypotheses 2020, 142, 109815. [Google Scholar] [CrossRef]
- Clegg, M.; Keen, C.; Hurley, L.S. Biochemical pathologies of zinc deficiency. In Zinc in Human Biology; Springer: Berlin/Heidelberg, Germany, 1989; pp. 129–145. [Google Scholar]
- Prakash, A.; Bharti, K.; Majeed, A.B.A. Zinc: Indications in brain disorders. Fundam. Clin. Pharmacol. 2015, 29, 131–149. [Google Scholar] [CrossRef]
- Li, D.-D.; Zhang, W.; Wang, Z.-Y.; Zhao, P. Serum copper, zinc, and iron levels in patients with Alzheimer’s disease: A meta-analysis of case-control studies. Front. Aging Neurosci. 2017, 9, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emwas, A.-H.; Alghrably, M.; Dhahri, M.; Sharfalddin, A.; Alsiary, R.; Jaremko, M.; Faa, G.; Campagna, M.; Congiu, T.; Piras, M. Living with the enemy: From protein-misfolding pathologies we know, to those we want to know. Ageing Res. Rev. 2021, 70, 101391. [Google Scholar] [CrossRef]
- Barman, S.; Srinivasan, K. Diabetes and zinc dyshomeostasis: Can zinc supplementation mitigate diabetic complications? Crit. Rev. Food Sci. Nutr. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Sun, W.; Fu, Y.; Miao, L.; Cai, L. Zinc homeostasis in the metabolic syndrome and diabetes. Front. Med. 2013, 7, 31–52. [Google Scholar] [CrossRef]
- Krebs, N.F. Overview of zinc absorption and excretion in the human gastrointestinal tract. J. Nutr. 2000, 130, 1374S–1377S. [Google Scholar] [CrossRef] [Green Version]
- Sandström, B.; Fairweather-Tait, S.; Hurrell, R.; Van Dokkum, W. Methods for studying mineral and trace element absorption in humans using stable isotopes. Nutr. Res. Rev. 1993, 6, 71–95. [Google Scholar] [CrossRef]
- Steinhardt, H.J.; Adibi, S.A. Interaction between transport of zinc and other solutes in human intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 1984, 247, G176–G182. [Google Scholar] [CrossRef]
- Lee, H.H.; Prasad, A.S.; Brewer, G.J.; Owyang, C. Zinc absorption in human small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 1989, 256, G87–G91. [Google Scholar] [CrossRef]
- Küry, S.; Dréno, B.; Bézieau, S.; Giraudet, S.; Kharfi, M.; Kamoun, R.; Moisan, J.-P. Identification of SLC39A4, a gene involved in acrodermatitis enteropathica. Nat. Genet. 2002, 31, 239–240. [Google Scholar] [PubMed]
- Wang, K.; Zhou, B.; Kuo, Y.-M.; Zemansky, J.; Gitschier, J. A novel member of a zinc transporter family is defective in acrodermatitis enteropathica. Am. J. Hum. Genet. 2002, 71, 66–73. [Google Scholar] [CrossRef] [Green Version]
- McMahon, R.J.; Cousins, R.J. Regulation of the zinc transporter ZnT-1 by dietary zinc. Proc. Natl. Acad. Sci. USA 1998, 95, 4841–4846. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Kim, B.-E.; Petris, M.J.; Eide, D.J. The mammalian Zip5 protein is a zinc transporter that localizes to the basolateral surface of polarized cells. J. Biol. Chem. 2004, 279, 51433–51441. [Google Scholar] [CrossRef] [Green Version]
- Guthrie, G.J.; Aydemir, T.B.; Troche, C.; Martin, A.B.; Chang, S.-M.; Cousins, R.J. Influence of ZIP14 (slc39A14) on intestinal zinc processing and barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G171–G178. [Google Scholar] [CrossRef] [Green Version]
- Cragg, R.A.; Christie, G.R.; Phillips, S.R.; Russi, R.M.; Küry, S.B.; Mathers, J.C.; Taylor, P.M.; Ford, D. A novel zinc-regulated human zinc transporter, hZTL1, is localized to the enterocyte apical membrane. J. Biol. Chem. 2002, 277, 22789–22797. [Google Scholar] [CrossRef] [Green Version]
- Cragg, R.; Phillips, S.; Piper, J.; Varma, J.; Campbell, F.; Mathers, J.; Ford, D. Homeostatic regulation of zinc transporters in the human small intestine by dietary zinc supplementation. Gut 2005, 54, 469–478. [Google Scholar]
- Valentine, R.A.; Jackson, K.A.; Christie, G.R.; Mathers, J.C.; Taylor, P.M.; Ford, D. ZnT5 variant B is a bidirectional zinc transporter and mediates zinc uptake in human intestinal Caco-2 cells. J. Biol. Chem. 2007, 282, 14389–14393. [Google Scholar] [CrossRef] [Green Version]
- Fukunaka, A.; Fujitani, Y. Role of zinc homeostasis in the pathogenesis of diabetes and obesity. Int. J. Mol. Sci. 2018, 19, 476. [Google Scholar] [CrossRef] [Green Version]
- Scott, B.J.; Bradwell, A.R. Identification of the serum binding proteins for iron, zinc, cadmium, nickel, and calcium. Clin. Chem. 1983, 29, 629–633. [Google Scholar] [CrossRef]
- Reyes, J.G. Zinc transport in mammalian cells. Am. J. Physiol. Cell Physiol. 1996, 270, C401–C410. [Google Scholar] [CrossRef]
- Kimura, T.; Kambe, T. The functions of metallothionein and ZIP and ZnT transporters: An overview and perspective. Int. J. Mol. Sci. 2016, 17, 336. [Google Scholar] [CrossRef] [Green Version]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Hill, G.M.; Brewer, G.J.; Prasad, A.S.; Hydrick, C.R.; Hartmann, D.E. Treatment of Wilson’s disease with zinc. I. Oral zinc therapy regimens. Hepatology 1987, 7, 522–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessells, K.R.; Jorgensen, J.M.; Hess, S.Y.; Woodhouse, L.R.; Peerson, J.M.; Brown, K.H. Plasma zinc concentration responds rapidly to the initiation and discontinuation of short-term zinc supplementation in healthy men. J. Nutr. 2010, 140, 2128–2133. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.H.; Wuehler, S.E.; Peerson, J.M. The importance of zinc in human nutrition and estimation of the global prevalence of zinc deficiency. Food Nutr. Bull. 2001, 22, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Colvin, R.A.; Davis, N.; Nipper, R.W.; Carter, P.A. Zinc transport in the brain: Routes of zinc influx and efflux in neurons. J. Nutr. 2000, 130, 1484S–1487S. [Google Scholar] [CrossRef]
- Myers, S.A.; Nield, A.; Myers, M. Zinc transporters, mechanisms of action and therapeutic utility: Implications for type 2 diabetes mellitus. J. Nutr. Metab. 2012, 2012, 173712. [Google Scholar] [CrossRef] [Green Version]
- Kochańczyk, T.; Drozd, A.; Krężel, A. Relationship between the architecture of zinc coordination and zinc binding affinity in proteins–insights into zinc regulation. Metallomics 2015, 7, 244–257. [Google Scholar] [CrossRef]
- Osredkar, J.; Sustar, N. Copper and zinc, biological role and significance of copper/zinc imbalance. J. Clin. Toxicol. S 2011, 3, 0495. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, A.P.; Abubakar, M.B.; Malami, I.; Ibrahim, K.G.; Abubakar, B.; Bello, M.B.; Qusty, N.; Elazab, S.T.; Imam, M.U.; Alexiou, A. Zinc Metalloproteins in Epigenetics and Their Crosstalk. Life 2021, 11, 186. [Google Scholar] [CrossRef]
- Brito, S.; Lee, M.-G.; Bin, B.-H.; Lee, J.-S. Zinc and its transporters in epigenetics. Mol. Cells 2020, 43, 323. [Google Scholar]
- Kumar, N.; Satarker, S.; Nampoothiri, M. Zinc as a plausible epigenetic modulator of glioblastoma multiforme. Eur. J. Pharmacol. 2020, 887, 173549. [Google Scholar]
- Available online: www.drugbank.ca (accessed on 1 July 2021).
- Mohammad, M.K.; Zhou, Z.; Cave, M.; Barve, A.; McClain, C.J. Zinc and liver disease. Nutr. Clin. Pract. 2012, 27, 8–20. [Google Scholar] [CrossRef]
- Berger, M.M.; Baines, M.; Raffoul, W.; Benathan, M.; Chiolero, R.L.; Reeves, C.; Revelly, J.-P.; Cayeux, M.-C.; Sénéchaud, I.; Shenkin, A. Trace element supplementation after major burns modulates antioxidant status and clinical course by way of increased tissue trace element concentrations. Am. J. Clin. Nutr. 2007, 85, 1293–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, M.M.; Binnert, C.; Chiolero, R.L.; Taylor, W.; Raffoul, W.; Cayeux, M.-C.; Benathan, M.; Shenkin, A.; Tappy, L. Trace element supplementation after major burns increases burned skin trace element concentrations and modulates local protein metabolism but not whole-body substrate metabolism. Am. J. Clin. Nutr. 2007, 85, 1301–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, M.M.; Spertini, F.; Shenkin, A.; Wardle, C.; Wiesner, L.; Schindler, C.; Chiolero, R.L. Trace element supplementation modulates pulmonary infection rates after major burns: A double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 1998, 68, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Cvijanovich, N.Z.; King, J.C.; Flori, H.R.; Gildengorin, G.; Vinks, A.A.; Wong, H.R. Safety and dose escalation study of intravenous zinc supplementation in pediatric critical illness. J. Parenter. Enter. Nutr. 2016, 40, 860–868. [Google Scholar] [CrossRef] [Green Version]
- Hemilä, H. Zinc lozenges and the common cold: A meta-analysis comparing zinc acetate and zinc gluconate, and the role of zinc dosage. JRSM Open 2017, 8, 2054270417694291. [Google Scholar] [CrossRef]
- Brown, M.A.; Thom, J.V.; Orth, G.L.; Cova, P.; Juarez, J. Food poisoning involving zinc contamination. Arch. Environ. Health Int. J. 1964, 8, 657–660. [Google Scholar] [CrossRef]
- Fosmire, G.J. Zinc toxicity. Am. J. Clin. Nutr. 1990, 51, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Nriagu, J. Zinc Toxicity in Humans; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–7. [Google Scholar]
- Yuzbasiyan-Gurkan, V.A.; Brewer, G.J.; Vander, A.J.; Guenther, M.J.; Prasad, A.S. Net renal tubular reabsorption of zinc in healthy man and impaired handling in sickle cell anemia. Am. J. Hematol. 1989, 31, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.; Chung, H.-E.; Yu, J.; Lee, J.-A.; Kim, T.-H.; Oh, J.-M.; Lee, W.-J.; Paek, S.-M.; Lee, J.K.; Jeong, J. Pharmacokinetics, tissue distribution, and excretion of zinc oxide nanoparticles. Int. J. Nanomed. 2012, 7, 3081. [Google Scholar]
- Available online: https://www.nlm.nih.gov/toxnet/index.html (accessed on 1 July 2021).
- Glasgow, J. Malabsorption syndrome. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Academic Press: Cambridge, MA, USA, 2003. [Google Scholar]
- D’Amico, F.; Baumgart, D.C.; Danese, S.; Peyrin-Biroulet, L. Diarrhea during COVID-19 infection: Pathogenesis, epidemiology, prevention, and management. Clin. Gastroenterol. Hepatol. 2020, 18, 1663–1672. [Google Scholar] [CrossRef]
- Pilloni, L.; Lecca, S.; Coni, P.; Demelia, L.; Pilleri, G.; Spiga, E.; Faa, G.; Ambu, R. Wilson’s disease with late onset. Dig. Liver Dis. 2000, 32, 180. [Google Scholar] [CrossRef]
- Lecca, S.; Pilloni, L.; Ambu, R.; Flore, C.; Callea, F.; Faa, G. Multiple histochemical methods in the diagnosis of Wilson’s disease. (Presentation of 74 cases and review of the literature). Pathologica 1998, 90, 771–775. [Google Scholar] [PubMed]
- Chang, I.J.; Hahn, S.H. The genetics of Wilson disease. Handb. Clin. Neurol. 2017, 142, 19–34. [Google Scholar]
- Fanni, D.; Fanos, V.; Gerosa, C.; Piras, M.; Dessi, A.; Atzei, A.; Van, E.P.; Gibo, Y.; Faa, G. Effects of iron and copper overload on the human liver: An ultrastructural study. Curr. Med. Chem. 2014, 21, 3768–3774. [Google Scholar] [CrossRef]
- Gerosa, C.; Fanni, D.; Congiu, T.; Piras, M.; Cau, F.; Moi, M.; Faa, G. Liver pathology in Wilson’s disease: From copper overload to cirrhosis. J. Inorg. Biochem. 2019, 193, 106–111. [Google Scholar] [CrossRef]
- US EPA. Estimation Program Interface (EPI) Suite. Ver. 4.1. November 2012. Available online: https://www2.epa.gov/tsca-screening-tools (accessed on 1 July 2021).
- Lech, T.; Sadlik, J. Copper concentration in body tissues and fluids in normal subjects of southern Poland. Biol. Trace Elem. Res. 2007, 118, 10–15. [Google Scholar] [CrossRef]
- Horoupian, D.; Sternlieb, I.; Scheinberg, I. Neuropathological findings in penicillamine-treated patients with Wilson’s disease. Clin. Neuropathol. 1988, 7, 62–67. [Google Scholar]
- Walshe, J. Penicillamine, a new oral therapy for Wilson’s disease. Am. J. Med. 1956, 21, 487–495. [Google Scholar] [CrossRef]
- Walshe, J. Treatment of Wilson’s disease with trientine (triethylene tetramine) dihydrochloride. Lancet 1982, 319, 643–647. [Google Scholar] [CrossRef]
- Hoogenraad, T.; Koevoet, R.; de Ruyter Korver, E. Oral zinc sulphate as long-term treatment in Wilson’s disease (hepatolenticular degeneration). Eur. Neurol. 1979, 18, 205–211. [Google Scholar] [CrossRef]
- Yuzbasiyan-Gurkan, V.; Grider, A.; Nostrant, T.; Cousins, R.J.; Brewer, G.J. Treatment of Wilson’s disease with zinc: X. Intestinal metallothionein induction. J. Lab. Clin. Med. 1992, 120, 380–386. [Google Scholar]
- Schouwink, G. De Hepato-Cerebral Degeneration (Met Een Onderzoekvan de Zinknkstofwisseling). Master’s Thesis, University of Amsterdam, Amsterdam, The Netherlands, 1961. [Google Scholar]
- Hoogenraad, T.; Van den Hamer, C.; Koevoet, R.; Korver, E.D.R. Oral zinc in Wilson’s disease. Lancet 1978, 312, 1262. [Google Scholar] [CrossRef]
- Dooley, J.S.; Purchase, R. History of Wilson Disease. In Wilson Disease; Weiss, K.H., Schilsky, M., Eds.; Academic Press: London, UK, 2019; Chapter 1; pp. 3–11. [Google Scholar]
- Hou, H.; Chen, D.; Liu, J.; Feng, L.; Zhang, J.; Liang, X.; Xu, Y.; Li, X. Zinc monotherapy for young patients with oligosymptomatic Wilson disease: A single center, retrospective study. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101623. [Google Scholar] [CrossRef]
- Litwin, T.; Dusek, P.; Skowrońska, M.; Członkowska, A. Treatment of Wilson’s disease–an update. Expert Opin. Orphan Drugs 2019, 7, 287–294. [Google Scholar] [CrossRef]
- Appenzeller-Herzog, C.; Mathes, T.; Heeres, M.L.; Weiss, K.H.; Houwen, R.H.; Ewald, H. Comparative effectiveness of common therapies for Wilson disease: A systematic review and meta-analysis of controlled studies. Liver Int. 2019, 39, 2136–2152. [Google Scholar] [CrossRef] [PubMed]
- Wiggelinkhuizen, M.; Tilanus, M.; Bollen, C.; Houwen, R. Systematic review: Clinical efficacy of chelator agents and zinc in the initial treatment of Wilson disease. Aliment. Pharmacol. Ther. 2009, 29, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Wiernicka, A.; Jańczyk, W.; Dądalski, M.; Avsar, Y.; Schmidt, H.; Socha, P. Gastrointestinal side effects in children with Wilson’s disease treated with zinc sulphate. World J. Gastroenterol. 2013, 19, 4356. [Google Scholar] [CrossRef]
- Lang, C.J.; Rabas-Kolominsky, P.; Engelhardt, A.; Kobras, G.; Konig, H.J. Fatal deterioration of Wilson’s disease after institution of oral zinc therapy. Arch. Neurol. 1993, 50, 1007–1008. [Google Scholar] [CrossRef]
- Walshe, J.; Munro, N. Zinc-induced deterioration in Wilson’s disease aborted by treatment with penicillamine, dimercaprol, and a novel zero copper diet. Arch. Neurol. 1995, 52, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Gong, J.-Y.; Yang, J.; Wang, J.-S. Anemia following zinc treatment for Wilson’s disease: A case report and literature review. BMC Gastroenterol. 2019, 19, 1–6. [Google Scholar] [CrossRef]
- European Association For The Study Of The Liver. EASL clinical practice guidelines: Wilson’s disease. J. Hepatol. 2012, 56, 671–685. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.A.; Schilsky, M.L. Diagnosis and treatment of Wilson disease: An update. Hepatology 2008, 47, 2089–2111. [Google Scholar] [CrossRef]
- Socha, P.; Janczyk, W.; Dhawan, A.; Baumann, U.; D’Antiga, L.; Tanner, S.; Iorio, R.; Vajro, P.; Houwen, R.; Fischler, B. Wilson’s disease in children: A position paper by the Hepatology Committee of the European Society for Paediatric Gastroenterology, Hepatology and Nutrition. J. Pediatric Gastroenterol. Nutr. 2018, 66, 334–344. [Google Scholar] [CrossRef]
- Zaino, D.; Chiarotti, I.; Battisti, C.; Salvatore, S.; Federico, A.; Cerase, A. Six-year clinical and MRI quantitative susceptibility mapping (QSM) follow-up in neurological Wilson’s disease under zinc therapy: A case report. Neurol. Sci. 2019, 40, 199–201. [Google Scholar] [CrossRef]
- Nobelpharma Co. Ltd. Nobelzin (Zinc Acetate). Japanese Package Insert; Nobelpharma Co. Ltd.: Tokyo, Japan, 2019. [Google Scholar]
- Sternlieb, I. Wilson’s Disease and Pregnancy; Wiley Online Library: Hoboken, NJ, USA, 2000. [Google Scholar]
- Isagawa, S.; Shiohira, H.; Hokama, N.; Nitta, H.; Yoshida, T.; Masamoto, H.; Aoki, Y.; Nakamura, K. Measurement of zinc concentration in blood and breast milk of a Wilson’s disease patient taking zinc acetate. Die Pharm. Int. J. Pharm. Sci. 2020, 75, 177–178. [Google Scholar]
- Hardyman, J.; Tyson, J.; Jackson, K.; Aldridge, C.; Cockell, S.; Wakeling, L.; Valentine, R.; Ford, D. Zinc sensing by metal-responsive transcription factor 1 (MTF1) controls metallothionein and ZnT1 expression to buffer the sensitivity of the transcriptome response to zinc. Metallomics 2016, 8, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Barber, R.; Grenier, Z.A.; Burkhead, J.L. Copper Toxicity Is Not Just Oxidative Damage: Zinc Systems and Insight from Wilson Disease. Biomedicines 2021, 9, 316. [Google Scholar] [CrossRef]
- Day, C. Non-alcoholic steatohepatitis (NASH): Where are we now and where are we going? Gut 2002, 50, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Fathi, M.; Alavinejad, P.; Haidari, Z.; Amani, R. The effects of zinc supplementation on metabolic profile and oxidative stress in overweight/obese patients with non-alcoholic fatty liver disease: A randomized, double-blind, placebo-controlled trial. J. Trace Elem. Med. Biol. 2020, 62, 126635. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, L.; Wang, J.; Xue, L.; Liu, L.; Yan, X.; Huang, S.; Li, Y.; Yan, X.; Zhang, B. Clinical features of patients with COVID-19 with nonalcoholic fatty liver disease. Hepatol. Commun. 2020, 4, 1758–1768. [Google Scholar] [CrossRef]
- PRASAD, A.S. The role of zinc in gastrointestinal and liver disease. Clin. Gastroenterol. 1983, 12, 713–741. [Google Scholar] [CrossRef]
- Himoto, T.; Nomura, T.; Tani, J.; Miyoshi, H.; Morishita, A.; Yoneyama, H.; Haba, R.; Masugata, H.; Masaki, T. Exacerbation of insulin resistance and hepatic steatosis deriving from zinc deficiency in patients with HCV-related chronic liver disease. Biol. Trace Elem. Res. 2015, 163, 81–88. [Google Scholar] [CrossRef]
- Himoto, T.; Tani, J.; Miyoshi, H.; Yoneyama, H.; Mori, H.; Inukai, M.; Masugata, H.; Goda, F.; Senda, S.; Haba, R. The ratio of insulin-like growth factor-I/insulin-like growth factor–binding protein-3 in sera of patients with hepatitis C virus–related chronic liver disease as a predictive marker of insulin resistance. Nutr. Res. 2013, 33, 27–33. [Google Scholar] [CrossRef]
- Dieck, H.T.; Döring, F.; Roth, H.-P.; Daniel, H. Changes in rat hepatic gene expression in response to zinc deficiency as assessed by DNA arrays. J. Nutr. 2003, 133, 1004–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viktorinova, A.; Tošerová, E.; Križko, M.; Ďuračková, Z. Altered metabolism of copper, zinc, and magnesium is associated with increased levels of glycated hemoglobin in patients with diabetes mellitus. Metabolism 2009, 58, 1477–1482. [Google Scholar] [CrossRef]
- Himoto, T.; Masaki, T. Associations between zinc deficiency and metabolic abnormalities in patients with chronic liver disease. Nutrients 2018, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Kosari, F.; Jamali, R.; Ramim, T.; Abad, E.M.J. The correlation between serum zinc level and liver histology in non-alcoholic steatohepatitis. Iran. J. Pathol. 2019, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Ishigami, M.; Ishizu, Y.; Kuzuya, T.; Honda, T.; Ishikawa, T.; Toyoda, H.; Kumada, T.; Fujishiro, M. Correlation of serum zinc levels with pathological and laboratory findings in patients with nonalcoholic fatty liver disease. Eur. J. Gastroenterol. Hepatol. 2020, 32, 748–753. [Google Scholar] [CrossRef]
- Qi, X.-Y.; Li, J.-Y.; Wang, Y.-D.; Zeng, Y.-W.; Liao, Z.-Z.; Ran, L.; Yang, J.; Wen, G.-B.; Liu, J.-H.; Xiao, X.-H. Association of serum zinc-α2-glycoprotein with non-alcoholic fatty liver disease. Chin. Med. J. 2020, 133, 1882. [Google Scholar] [CrossRef]
- Xiao, X.-H.; Wang, Y.-D.; Qi, X.-Y.; Wang, Y.-Y.; Li, J.-Y.; Li, H.; Zhang, P.-Y.; Liao, H.-L.; Li, M.-H.; Liao, Z.-Z. Zinc alpha2 glycoprotein protects against obesity-induced hepatic steatosis. Int. J. Obes. 2018, 42, 1418–1430. [Google Scholar] [CrossRef]
- Vranešić Bender, D.; Nutrizio, M.; Jošić, M.; Ljubas Kelečić, D.; Karas, I.; Premužić, M.; Domislović, V.; Rotim, C.; Krznarić, Ž. Nutritional status and nutrition quality in patients with non-alcoholic fatty liver disease. Acta Clin. Croat. 2017, 56, 625–634. [Google Scholar] [CrossRef]
- Kim, M.-H.; Aydemir, T.B.; Kim, J.; Cousins, R.J. Hepatic ZIP14-mediated zinc transport is required for adaptation to endoplasmic reticulum stress. Proc. Natl. Acad. Sci. USA 2017, 114, E5805–E5814. [Google Scholar] [CrossRef] [Green Version]
- Bjørklund, G.; Dadar, M.; Pivina, L.; Doşa, M.D.; Semenova, Y.; Aaseth, J. The role of zinc and copper in insulin resistance and diabetes mellitus. Curr. Med. Chem. 2020, 27, 6643–6657. [Google Scholar] [CrossRef]
- Himoto, T.; Yoneyama, H.; Deguch, A.; Kurokohchi, K.; Inukai, M.; Masugata, H.; Goda, F.; Senda, S.; Watanabe, S.; Kubota, S. Insulin resistance derived from zinc deficiency in non-diabetic patients with chronic hepatitis C. Exp. Ther. Med. 2010, 1, 707–711. [Google Scholar] [CrossRef] [Green Version]
- Koruk, M.; Taysi, S.; Savas, M.C.; Yilmaz, O.; Akcay, F.; Karakok, M. Oxidative stress and enzymatic antioxidant status in patients with nonalcoholic steatohepatitis. Ann. Clin. Lab. Sci. 2004, 34, 57–62. [Google Scholar] [PubMed]
- Su, T.-H.; Kao, J.-H. The clinical manifestations and management of COVID-19-related liver injury. J. Formos. Med. Assoc. 2020, 119, 1016. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kubota, Y.; Chernov, M.; Kasuya, H. Potential role of zinc supplementation in prophylaxis and treatment of COVID-19. Med. Hypotheses 2020, 144, 109848. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.P.; Mishra, S.; Jha, D.K.; Shukla, J.; Choudhury, A.; Mohindra, R.; Mandavdhare, H.S.; Dutta, U.; Sharma, V. Coronavirus disease (COVID-19) and the liver: A comprehensive systematic review and meta-analysis. Hepatol. Int. 2020, 14, 711–722. [Google Scholar] [CrossRef]
- Gholizadeh, P.; Safari, R.; Marofi, P.; Zeinalzadeh, E.; Pagliano, P.; Ganbarov, K.; Esposito, S.; Khodadadi, E.; Yousefi, M.; Kafil, H.S. Alteration of liver biomarkers in patients with SARS-CoV-2 (COVID-19). J. Inflamm. Res. 2020, 13, 285. [Google Scholar] [CrossRef]
- Chang, D.; Lin, M.; Wei, L.; Xie, L.; Zhu, G.; Cruz, C.S.D.; Sharma, L. Epidemiologic and clinical characteristics of novel coronavirus infections involving 13 patients outside Wuhan, China. JAMA 2020, 323, 1092–1093. [Google Scholar] [CrossRef]
- Li, L.; Li, S.; Xu, M.; Zheng, S.; Duan, Z.; Chen, Y.; Li, J. The level of plasma C-reactive protein is closely related to the liver injury in patients with COVID-19. MedRxiv 2020. [Google Scholar] [CrossRef]
- Fan, Z.; Chen, L.; Li, J.; Cheng, X.; Yang, J.; Tian, C.; Zhang, Y.; Huang, S.; Liu, Z.; Cheng, J. Clinical features of COVID-19-related liver functional abnormality. Clin. Gastroenterol. Hepatol. 2020, 18, 1561–1566. [Google Scholar] [CrossRef]
- Fu, L.; Fei, J.; Xu, S.; Xiang, H.-X.; Xiang, Y.; Tan, Z.-X.; Li, M.-D.; Liu, F.-F.; Li, Y.; Han, M.-F. Acute liver injury and its association with death risk of patients with COVID-19: A hospital-based prospective case-cohort study. MedRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.-C.; Huo, T.-I.; Huang, Y.-H. Gastrointestinal and liver manifestations in patients with COVID-19. J. Chin. Med. Assoc. 2020, 83, 521–523. [Google Scholar] [CrossRef]
- Sivandzadeh, G.R.; Askari, H.; Safarpour, A.R.; Ejtehadi, F.; Raeis-Abdollahi, E.; Lari, A.V.; Abazari, M.F.; Tarkesh, F.; Lankarani, K.B. COVID-19 infection and liver injury: Clinical features, biomarkers, potential mechanisms, treatment, and management challenges. World J. Clin. Cases 2021, 9, 6178. [Google Scholar] [CrossRef]
- Sun, J.; Aghemo, A.; Forner, A.; Valenti, L. COVID-19 and liver disease. Liver 2020, 40, 1278–1281. [Google Scholar]
- Cai, Q.; Huang, D.; Ou, P.; Yu, H.; Zhu, Z.; Xia, Z.; Su, Y.; Ma, Z.; Zhang, Y.; Li, Z. COVID-19 in a designated infectious diseases hospital outside Hubei Province, China. Allergy 2020, 75, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G. Reduction and functional exhaustion of T cells in patients with coronavirus disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Li, J.; Fan, J.-G. Characteristics and mechanism of liver injury in 2019 coronavirus disease. J. Clin. Transl. Hepatol. 2020, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Zhu, B.; Liang, H.; Fang, C.; Gong, Y.; Guo, Q.; Sun, X.; Zhao, D.; Shen, J. Characteristics of pediatric SARS-CoV-2 infection and potential evidence for persistent fecal viral shedding. Nat. Med. 2020, 26, 502–505. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Han, B.; Wang, J. COVID-19: Gastrointestinal manifestations and potential fecal–oral transmission. Gastroenterology 2020, 158, 1518–1519. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, L.; Wang, F.-S. Liver injury in COVID-19: Management and challenges. Lancet Gastroenterol. Hepatol. 2020, 5, 428–430. [Google Scholar] [CrossRef]
- Polak, Y.; Speth, R.C. Metabolism of angiotensin peptides by angiotensin converting enzyme 2 (ACE2) and analysis of the effect of excess zinc on ACE2 enzymatic activity. Peptides 2021, 137, 170477. [Google Scholar] [CrossRef]
- Gruengreiff, K.; Reinhold, D. Zinc: A complementary factor in the treatment of chronic hepatitis C? Mol. Med. Rep. 2010, 3, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuasa, K.; Naganuma, A.; Sato, K.; Ikeda, M.; Kato, N.; Takagi, H.; Mori, M. Zinc is a negative regulator of hepatitis C virus RNA replication. Liver Int. 2006, 26, 1111–1118. [Google Scholar] [CrossRef]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The role of zinc in antiviral immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef] [Green Version]
- Sandstead, H.H.; Prasad, A.S. Zinc intake and resistance to H1N1 influenza. Am. J. Public Health 2010, 100, 970–971. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, H.; Tavakoli, A.; Moradi, A.; Tabarraei, A.; Bokharaei-Salim, F.; Zahmatkeshan, M.; Farahmand, M.; Javanmard, D.; Kiani, S.J.; Esghaei, M. Inhibition of H1N1 influenza virus infection by zinc oxide nanoparticles: Another emerging application of nanomedicine. J. Biomed. Sci. 2019, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Te Velthuis, A.J.; van den Worm, S.H.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn2+ inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Wright-Minogue, J.; Fang, J.W.; Baroudy, B.M.; Lau, J.Y.; Hong, Z. Characterization of soluble hepatitis C virus RNA-dependent RNA polymerase expressed in Escherichia coli. J. Virol. 1999, 73, 1649–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterworth, B.E.; Korant, B.D. Characterization of the large picornaviral polypeptides produced in the presence of zinc ion. J. Virol. 1974, 14, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Pasternak, C. A novel form of host defence: Membrane protection by Ca2+ and Zn2+. Biosci. Rep. 1987, 7, 81–91. [Google Scholar] [CrossRef]
- Ischia, J.; Bolton, D.M.; Patel, O. Why is it worth testing the ability of zinc to protect against ischaemia reperfusion injury for human application. Metallomics 2019, 11, 1330–1343. [Google Scholar] [CrossRef] [PubMed]
- O’Kane, D.; Gibson, L.; May, C.N.; du Plessis, J.; Shulkes, A.; Baldwin, G.S.; Bolton, D.; Ischia, J.; Patel, O. Zinc preconditioning protects against renal ischaemia reperfusion injury in a preclinical sheep large animal model. BioMetals 2018, 31, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.; Sethi, K.; Ischia, J.; Gibson, L.; Galea, L.; Xiao, L.; Yim, M.; Chang, M.; Papa, N.; Bolton, D. Protective effect of zinc preconditioning against renal ischemia reperfusion injury is dose dependent. PLoS ONE 2017, 12, e0180028. [Google Scholar] [CrossRef] [Green Version]
- Cheung, E.; Nikfarjam, M.; Jackett, L.; Bolton, D.M.; Ischia, J.; Patel, O. The protective effect of zinc against liver ischaemia reperfusion injury in a rat model of global ischaemia. J. Clin. Exp. Hepatol. 2020, 10, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.S. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J. Trace Elem. Med. Biol. 2014, 28, 364–371. [Google Scholar] [CrossRef]
- Basu, S. Non-communicable disease management in vulnerable patients during Covid-19. Indian J. Med. Ethics 2020, 2, 103–105. [Google Scholar] [CrossRef]
- Haase, H.; Rink, L. The immune system and the impact of zinc during aging. Immun. Ageing 2009, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dardenne, M. Zinc and immune function. Eur. J. Clin. Nutr. 2002, 56, S20–S23. [Google Scholar] [CrossRef] [Green Version]
- Faa, G.; Nurchi, V.M.; Ravarino, A.; Fanni, D.; Nemolato, S.; Gerosa, C.; Van Eyken, P.; Geboes, K. Zinc in gastrointestinal and liver disease. Coord. Chem. Rev. 2008, 252, 1257–1269. [Google Scholar] [CrossRef]
- Prasad, A.S. Impact of the discovery of human zinc deficiency on health. J. Am. Coll. Nutr. 2009, 28, 257–265. [Google Scholar] [CrossRef]
- Malavolta, M.; Piacenza, F.; Basso, A.; Giacconi, R.; Costarelli, L.; Mocchegiani, E. Serum copper to zinc ratio: Relationship with aging and health status. Mech. Ageing Dev. 2015, 151, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Oyagbemi, A.A.; Ajibade, T.O.; Aboua, Y.G.; Gbadamosi, I.T.; Adedapo, A.D.A.; Aro, A.O.; Adejumobi, O.A.; Thamahane-Katengua, E.; Omobowale, T.O.; Falayi, O.O. Potential health benefits of zinc supplementation for the management of COVID-19 pandemic. J. Food Biochem. 2021, 45, e13604. [Google Scholar] [CrossRef]
- Cao, J.-w.; Duan, S.-y.; Zhang, H.-x.; Chen, Y.; Guo, M. Zinc deficiency promoted fibrosis via ROS and TIMP/MMPs in the myocardium of mice. Biol. Trace Elem. Res. 2020, 196, 145–152. [Google Scholar] [CrossRef]
- Frontera, J.A.; Rahimian, J.O.; Yaghi, S.; Liu, M.; Lewis, A.; de Havenon, A.; Mainali, S.; Huang, J.; Scher, E.; Wisniewski, T. Treatment with Zinc is associated with reduced in-hospital mortality among COVID-19 patients: A multi-center cohort study. Res. Sq. 2020, rs. 3, rs-94509. [Google Scholar]
- Demurtas, P.; Orrù, G.; Coni, P.; Minerba, L.; Corrias, M.; Sirigu, P.; Zucca, I.; Demurtas, E.; Maxia, C.; Piras, F. Association between the ACE insertion/deletion polymorphism and pterygium in Sardinian patients: A population based case–control study. BMJ Open 2014, 4, e005627. [Google Scholar] [CrossRef] [Green Version]
- Beattie, J.H.; Kwun, I.-S. Is zinc deficiency a risk factor for atherosclerosis? Br. J. Nutr. 2004, 91, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Beattie, J.H.; Gordon, M.J.; Duthie, S.J.; McNeil, C.J.; Horgan, G.W.; Nixon, G.F.; Feldmann, J.; Kwun, I.S. Suboptimal dietary zinc intake promotes vascular inflammation and atherogenesis in a mouse model of atherosclerosis. Mol. Nutr. Food Res. 2012, 56, 1097–1105. [Google Scholar] [CrossRef]
- Saba, L.; Gerosa, C.; Fanni, D.; Marongiu, F.; La Nasa, G.; Caocci, G.; Barcellona, D.; Balestrieri, A.; Coghe, F.; Orru, G. Molecular pathways triggered by COVID-19 in different organs: ACE2 receptor-expressing cells under attack? A review. Eur. Rev. Med. Pharm. Sci. 2020, 24, 12609–12622. [Google Scholar]


{kind=link}
| NCT Number | Title | Status | Conditions | Interventions | Characteristics | Population | Locations | |
|---|---|---|---|---|---|---|---|---|
| 1 | NCT04542993 | Can SARS-CoV-2 Viral Load and COVID-19 Disease Severity be Reduced by Resveratrol-assisted Zinc Therapy | Active, not recruiting | •Covid19 •SARS-CoV Infection | •Dietary Supplement: Zinc Picolinate •Dietary Supplement: Resveratro •Dietary Supplement: Zinc Picolinate Placebo •Dietary Supplement: Resveratrol Placebo | Study TypeInterventional | Age: 18 Years to 75 Years (Adult, Older Adult) | •Swedish Medical Center, Seattle, Washington, United States |
| 2 | NCT04468139 | The Study of Quadruple Therapy Zinc, Quercetin, Bromelain and Vitamin C on the Clinical Outcomes of Patients Infected With COVID-19 | Recruiting | •Covid-19 | •Drug: Quercetin •Dietary Supplement: bromelain•Drug: Zinc •Drug: Vitamin C | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •Ministry of health. First health cluster, Riaydh, Riyadh, Saudi Arabia |
| 3 | NCT05003492 | Utilizing the Crosstalk Among Aerosolized Phenformin, Methylene Blue, Photodynamic Therapy, Zinc and Potassium for Treating Severe COVID-19 Infection and Its Inflammatory Complication | Not yet recruiting | •COVID-19 | •Combination Product: Combination therapy plus Standard therapy•Radiation: Photodynamic therapy •Drug: Standard therapy | Study Type: Interventional | Age: 18 Years to 70 Years (Adult, Older Adult) | •Ministry of health. First health cluster, Riaydh, Riyadh, Saudi Arabia |
| 4 | NCT04395768 | International ALLIANCE Study of Therapies to Prevent Progression of COVID-19 | Recruiting | •COVID19 | •Dietary Supplement: Vitamin C •Drug: Hydroxychloroquine •Drug: Azithromycin. •Dietary Supplement: Zinc Citrate •Dietary Supplement: VitaminD3 •Dietary Supplement: Vitamin B12 | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •National Institute of Integrative Medicine, Melbourne, Victoria, Australia |
| 5 | NCT04447534 | Zinc With Chloroquine/ Hydroxychloroquine in Treatment of COVID-19 | Recruiting | •COVID | •Drug: Chloroquine •Drug: zinc | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •Tanta university hospital, Assuit University, Ainshams University, Tanta, Egypt |
| 6 | NCT04828538 | Vitamin D, Omega-3, and Combination Vitamins B, C and Zinc Supplementation for the Treatment and Prevention of COVID-19 | Active, not recruiting | •Covid19 | •Dietary Supplement: VitaminD •Dietary Supplement: Omega DHA/EPA •Dietary Supplement: Vitamin C, Vitamin B complex and Zinc Acetate | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •Hospital de Soledad, San Luis Potosí, SLP, Mexico |
| 7 | NCT04370782 | Hydroxychloroquine and Zinc With Either Azithromycin or Doxycycline for Treatment of COVID-19 in Outpatient Setting | Completed | •COVID-19 | •Drug: Hydroxychloroquine •Drug: Azithromycin •Drug: Zinc Sulfate •Drug: Doxycycline | Study Type: Interventional | Age: 30 Years and older (Adult, Older Adult) | •St Francis Hospital, Roslyn, New York, United States |
| 8 | NCT04641195 | Vitamin D and Zinc Supplementation for Improving Treatment Outcomes Among COVID-19 Patients in India | Recruiting | •COVID-19 | •Dietary Supplement: Vitamin D3 (cholecalciferol) •Dietary Supplement: Zinc (zinc gluconate) •Dietary Supplement: Zinc (zinc gluconate) & Vitamin D(cholecalciferol) •Other: Placebo | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •Saifee Hospital, Mumbai, Maharashtra, India •King Edward Memorial (KEM) Hospital, Pune, Maharashtra, India |
| 9 | NCT04621461 | Placebo Controlled Trial to Evaluate Zinc for the Treatment of COVID-19 in the Outpatient Setting | Completed | •Corona Virus Infection | •Dietary Supplement: Zinc Sulfate 220 MG •Drug: Placebo | Study Type: Interventional | Age: 30 Years and older (Adult, Older Adult) | •St. Francis Hospital—The Heart Center, Roslyn, New York, United States |
| 10 | NCT00212368 | Efficacy and Safety Study of Zinc Acetate to Treat Wilson’s Disease in Japan. | Completed | •Wilson’s Disease | •Drug: Zinc acetate | Study Type: Interventional | Age: 1 Year and older (Child, Adult, Older Adult) | |
| 11 | NCT00212355 | Efficacy and Safety, Long-term Study of Zinc Acetate to Treat Wilson’s Disease in Japan. | Completed | •Wilson’s Disease | •Drug: NPC-02 | Study Type: Interventional | Age: 1 Year and older (Child, Adult, Older Adult) | |
| 12 | NCT03659331 | A Controlled Study of Potential Therapeutic Effect of Oral Zinc in Manifesting Carriers of Wilson’s Disease | Unknown status | •Wilson’s Disease | •Dietary Supplement: Zinc | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | |
| 13 | NCT02475928 | Zinc Supplementation in Cirrhotic Patients | Unknown status | •Dysgeusia •Liver Cirrhosis | •Dietary Supplement: zinc gluconate •Dietary Supplement: Placebo •Behavioral: Nutritional education | Study Type: Interventional | Age: 18 Years to 70 Years (Adult, Older Adult) | •Medica Sur Clinic & Foundation, Mexico City, Mexico |
| 14 | NCT01899521 | Examination of Zinc, S- adenosylmethionine, and Combination Therapy Versus Placebo in Alcoholics | Completed | •Alcoholism | •Procedure: Bronchoscopy •Dietary Supplement: Zinc sulfate 220 mg once daily •Dietary Supplement: Sadenosylmethionine 400 mg twice daily | Study Type: Interventional | Age: 18 Years to 60 Years (Adult) | •Atlanta VA Medical and Rehab Center, Decatur, GA, Decatur, Georgia, United States |
| 15 | NCT00149552 | Zinc Therapy in HIV Infected Individuals Who Abuse Drugs | Completed | •HIV Infections •Substance-Related Disorders | •Dietary Supplement: zinc | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •Camillus House, Miami, Florida, United States |
| 16 | NCT00449592 | Oral Zinc Therapy for the Prevention of Mucositis | Completed | •Mucositis | •Drug: Zinc •Drug: Placebo | Study Type: Interventional | Age: 18 Years to 70 Years (Adult, Older Adult) | •Division of Hematology and Bone Marrow Transplantation, Sheba Medical Center, Ramat-Gan, Israel |
| 17 | NCT00325247 | Efficacy of Zinc Therapy in Acute Diarrhoea in Young Children | Completed | •Acute Watery Diarrhea | •Drug: ZINC | Study Type: Interventional | Age: 1 Month to 59 Months (Child) | •ICDDR,B, Dhaka, Bangladesh |
| 18 | NCT01440608 | Effectiveness of High-dose Zinc Therapy and Albendazole in the Treatment of Environmental Enteropathy | Completed | •Enteropathy | •Drug: Albendazole •Drug: Placebo •Dietary Supplement: High dose zinc | Study Type: Interventional | Age: 1 Year to 3 Years (Child) | •Saint Louis Nutrition Project, Blantyre, Malawi |
| 19 | NCT01162109 | Zinc Therapy in Critical Illness | Active, not recruiting | •Severe Sepsis | •Dietary Supplement: Zinc sulfate | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •University of Vermont College of Medicine, Burlington, Vermont, United States |
| 20 | NCT01259050 | Safety Study of High Doses of Zinc in ALS Patients | Completed | •Amyotrophic Lateral Sclerosis | •Drug: Zinc and Copper | Study Type: Interventional | Age: 18 Years to 85 Years (Adult, Older Adult) | •Phoenix Neurological Associates, Phoenix, Arizona, United States |
| 21 | NCT00252304 | Therapeutic Zinc in Childhood Pneumonia | Completed | •Pneumonia | •Drug: Zinc (zinc sulphate) •Drug: Placebo | Study Type: Interventional | Age: 2 Months to 35 Months (Child) | •Kanti Children Hospital, Kathmandu, Nepal |
| 22 | NCT00693680 | Zinc Supplementation of Imipramine Therapy | Completed | •Major Depression | •Dietary Supplement: Zincas Forte •Dietary Supplement: Placebo | Study Type: Interventional | Age: 18 Years and older (Adult, Older Adult) | •Department of Psychiatry, Collegium Medicum, Jagiellonian University, Krakow, Poland |
| 23 | NCT02601742 | Effectiveness of Oral Rehydration Therapy Supplemented With Zinc in the Management of Diarrhea Acute | Unknown status | •Diarrhea •Children | •Other: Zinc group •Other: Placebo group | Study Type: Interventional | Age: 6 Months to 5 Years (Child) | |
| 24 | NCT03923829 | The Effect of Zinc on the Gingival Crevicular Fluid Level of Total Oxidant Capacity in Type 2 Diabetic Patients | Unknown status | •Chronic Periodontitis | •Drug: Zinc Sulfate •Procedure: scaling and root planing | Study Type: Interventional | Age: Child, Adult, Older Adult |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coni, P.; Pichiri, G.; Lachowicz, J.I.; Ravarino, A.; Ledda, F.; Fanni, D.; Gerosa, C.; Piras, M.; Coghe, F.; Gibo, Y.; et al. Zinc as a Drug for Wilson’s Disease, Non-Alcoholic Liver Disease and COVID-19-Related Liver Injury. Molecules 2021, 26, 6614. https://doi.org/10.3390/molecules26216614
Coni P, Pichiri G, Lachowicz JI, Ravarino A, Ledda F, Fanni D, Gerosa C, Piras M, Coghe F, Gibo Y, et al. Zinc as a Drug for Wilson’s Disease, Non-Alcoholic Liver Disease and COVID-19-Related Liver Injury. Molecules. 2021; 26(21):6614. https://doi.org/10.3390/molecules26216614
Chicago/Turabian StyleConi, Pierpaolo, Giuseppina Pichiri, Joanna Izabela Lachowicz, Alberto Ravarino, Francesca Ledda, Daniela Fanni, Clara Gerosa, Monica Piras, Ferdinando Coghe, Yukio Gibo, and et al. 2021. "Zinc as a Drug for Wilson’s Disease, Non-Alcoholic Liver Disease and COVID-19-Related Liver Injury" Molecules 26, no. 21: 6614. https://doi.org/10.3390/molecules26216614
APA StyleConi, P., Pichiri, G., Lachowicz, J. I., Ravarino, A., Ledda, F., Fanni, D., Gerosa, C., Piras, M., Coghe, F., Gibo, Y., Cau, F., Castagnola, M., Van Eyken, P., Saba, L., Piludu, M., & Faa, G. (2021). Zinc as a Drug for Wilson’s Disease, Non-Alcoholic Liver Disease and COVID-19-Related Liver Injury. Molecules, 26(21), 6614. https://doi.org/10.3390/molecules26216614