
Isoflavone Changes in Immature and Mature Soybeans by Thermal Processing


Abstract
:1. Introduction
2. Results and Discussion
2.1. Physiological Characteristics of Immature and Mature Soybeans
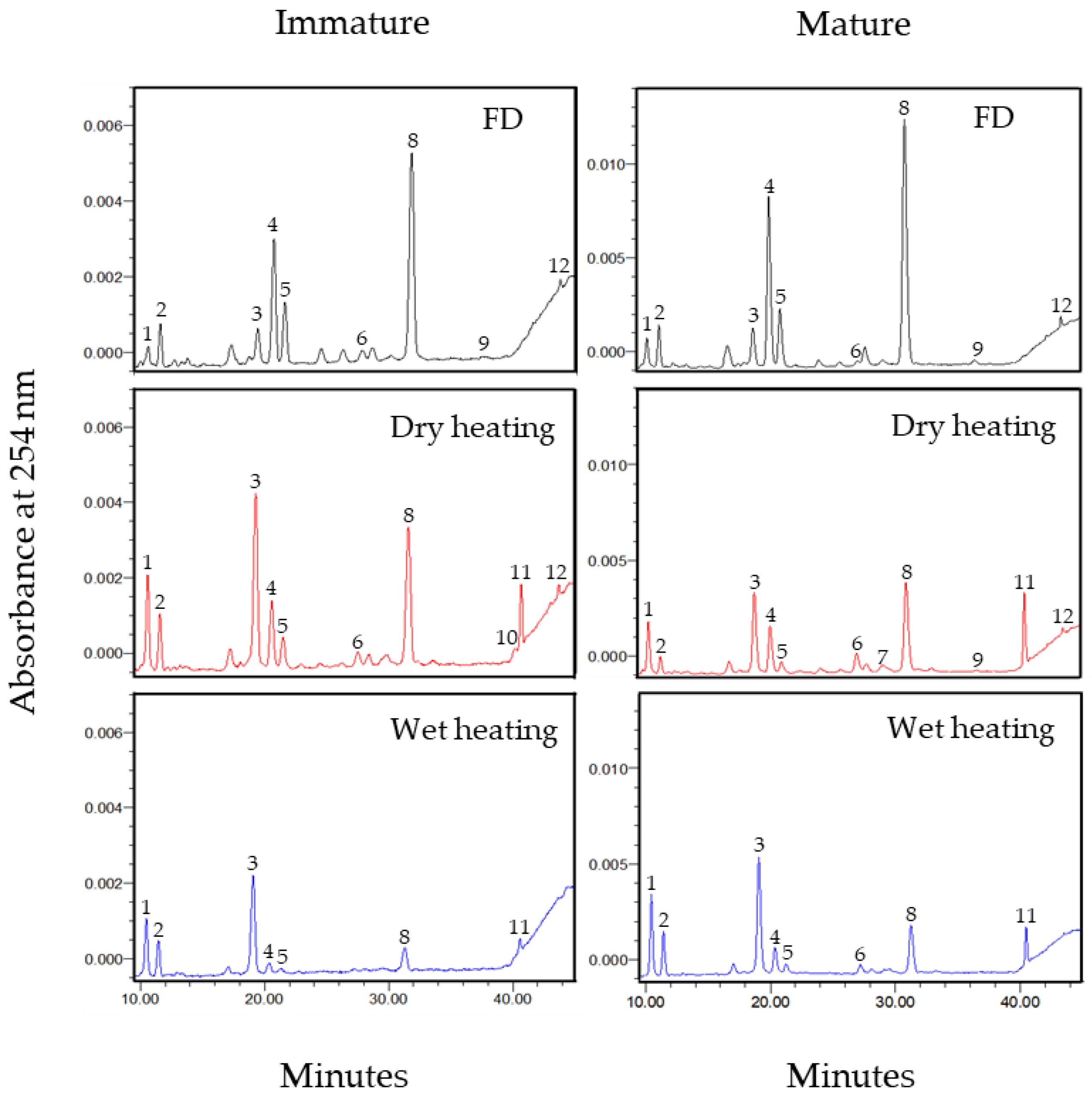
2.2. The Variation in the Total Isoflavone Content and Four Isoflavone Forms of Soybeans during Thermal Processing
2.3. Correlation Analysis between Isoflavone Form and Corresponding Individual Isoflavones in Immature and Mature Soybeans
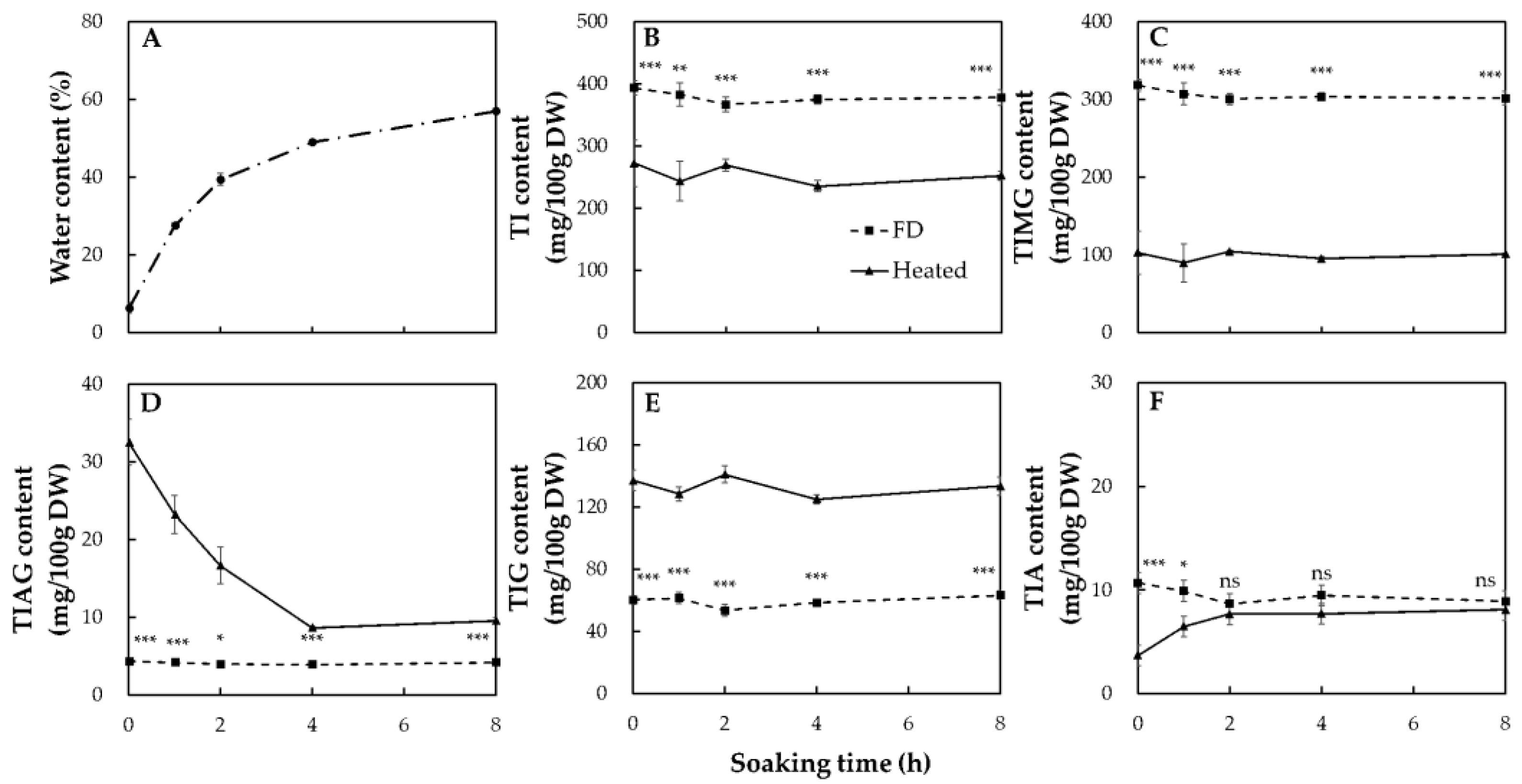
2.4. Verification of the Relationship between Soybean Water Content and Changes in Patterns of Isoflavone Contents
3. Materials and Methods
3.1. Chemical Reagents
3.2. Soybean Cultivation
3.3. Physical Characteristics of Immature and Mature Seeds
3.4. Thermal Treatment
3.5. Extraction of Isoflavones
3.6. Determination of Isoflavones
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- O’Keefe, S.; Bianchi, L.; Sharman, J. Soybean nutrition. J. Nutr. Metab. 2015, 1, 1006. [Google Scholar]
- Kim, H.K.; Kang, S.T.; Cho, J.H.; Choung, M.G.; Suh, D.Y. Quantitative trait loci associated with oligosaccharide and sucrose contents in soybean (Glycine max L.). J. Plant Biol. 2005, 48, 106–112. [Google Scholar] [CrossRef]
- Ollberding, N.J.; Lim, U.; Wilkens, L.R.; Setiawan, V.W.; Shvetsov, Y.B.; Henderson, B.E.; Kolonel, L.N.; Goodman, M.T. Legume, soy, tofu, and isoflavone intake and endometrial cancer risk in postmenopausal women in the multiethnic cohort study. J. Natl. Cancer Inst. 2012, 104, 67–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.S.; Rha, C.S.; Baik, M.Y.; Baek, N.I.; Kim, D.O. A brief history and spectroscopic analysis of soy isoflavones. Food Sci. Biotechnol. 2020, 29, 1605–1617. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.J.; Lyu, J.I.; Kwon, S.J.; Eom, S.H. Effects of UV-A radiation on organ-specific accumulation and gene expression of isoflavones and flavonols in soybean sprout. Food Chem. 2021, 339, 128080. [Google Scholar] [CrossRef]
- Hoeck, J.A.; Fehr, W.R.; Murphy, P.A.; Welke, J.A. Influence of genotype and environment on isoflavone contents of soybean. Crop Sci. 2000, 40, 48–51. [Google Scholar] [CrossRef]
- Barnes, S.; Peterson, T.G. Biochemical targets of the isoflavone genistein in tumor cell lines. Proc. Soc. Exp. Biol. Med. 1995, 208, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Honjo, H.; Higashi, A.; Fotsis, T.; Hämäläinen, E.; Hasegawa, T.; Okada, H. Urinary excretion of lignans and isoflavonoid phytoestrogens in Japanese men and women consuming a traditional Japanese diet. Am. J. Clin. Nutr. 1991, 54, 1093–1100. [Google Scholar] [CrossRef]
- Knight, D.C.; Eden, J.A. A review of the clinical effects of phytoestrogens. Obstet. Gynecol. 1996, 87, 897–904. [Google Scholar]
- Xu, B.; Chang, S.K.C. Total phenolics, phenolic acids, isoflavones, and anthocyanins and antioxidant properties of yellow and black soybeans as affected by thermal processing. J. Agric. Food Chem. 2008, 56, 7165–7175. [Google Scholar] [CrossRef]
- Severson, R.K.; Nomura, A.M.Y.; Grove, J.S.; Stemmermann, G.N. A prospective study of demographics, diet, and prostate cancer among men of Japanese ancestry in Hawaii. Cancer Res. 1989, 49, 1857–1860. [Google Scholar] [PubMed]
- Lee, H.P.; Lee, J.; Gourley, L.; Duffy, S.W.; Day, N.E.; Estève, J. Dietary effects on breast-cancer risk in Singapore. Lancet 1991, 337, 1197–1200. [Google Scholar] [CrossRef]
- Yerramsetty, V.; Gallaher, D.D.; Ismail, B. Malonylglucoside conjugates of isoflavones are much less bioavailable compared with unconjugated β-glucosidic forms in rats. J. Nutr. 2014, 144, 631–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.H.; Yang, L.; Xu, J.Z.; Yeung, S.Y.V.; Huang, Y.; Chen, Z.Y. Relative antioxidant activity of soybean isoflavones and their glycosides. Food Chem. 2005, 90, 735–741. [Google Scholar] [CrossRef]
- Izumi, T.; Pikula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy isoflavone aglycones are absorbed faster and in higher amount than their glucosides in humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, W.J. Lipoxygenase and flavor of soybean protein products. J. Agric. Food Chem. 1975, 23, 136–141. [Google Scholar] [CrossRef]
- MacLeod, G.; Ames, J.; Betz, N.L. Soy flavor and its improvement. Crit. Rev. Food Sci. Nutr. 1989, 27, 219–400. [Google Scholar] [CrossRef]
- Cai, J.S.; Zh, Y.Y.; Ma, R.H.; Thakur, K.; Zhang, J.G.; Wei, Z.J. Effects of roasting level on physicochemical, sensory, and volatile profiles of soybeans using electronic nose and HS-SPME-GC–MS. Food Chem. 2021, 340, 127880. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, S.; Liu, Z.; Chang, S.K. Off-flavor related volatiles in soymilk as affected by soybean variety, grinding, and heat-processing methods. J. Agric. Food Chem. 2012, 60, 7457–7462. [Google Scholar] [CrossRef]
- Endo, H.; Ohno, M.; Tanji, K.; Shimada, S.; Kaneko, K. Effect of heat treatment on the lipid peroxide content and aokusami (beany flavor) of soymilk. Food Sci. Technol. Res. 2007, 10, 328–333. [Google Scholar] [CrossRef] [Green Version]
- Villares, A.; Rostagno, M.A.; García-Lafuente, A.; Guillamón, E.; Martínez, J.A. Content and profile of isoflavones in soy-based foods as a function of the production process. Food Bioprocess Technol. 2011, 4, 27–38. [Google Scholar] [CrossRef]
- Muliterno, M.M.; Rodrigues, D.; Lima, F.S.; Ida, E.L.; Kurozawa, L.E. Conversion/degradation of isoflavones and color alterations during the drying of okara. LWT 2017, 75, 512–519. [Google Scholar] [CrossRef]
- Chan, S.G.; Murphy, P.A.; Ho, S.C.; Kreiger, N.; Darlington, G.; So, E.K.F.; Chong, P.Y.Y. Isoflavonoid content of Hong Kong soy foods. J. Agric. Food Chem. 2009, 57, 5386–5390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chang, S.K.; Liu, Z. Isoflavone profile in soymilk as affected by soybean variety, grinding, and heat-processing methods. J. Food Sci. 2015, 80, C983–C988. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, B.W.; Kim, B.; Kim, H.T.; Ko, J.M.; Baek, I.Y.; Seo, W.T.; Kang, Y.M.; Cho, K.M. Changes in phenolic compounds (Isoflavones and Phenolic acids) and antioxidant properties in high-protein soybean (Glycine max L., cv. Saedanbaek) for different roasting conditions. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 605–612. [Google Scholar] [CrossRef]
- Yuan, J.P.; Liu, Y.B.; Peng, J.; Wang, J.H.; Liu, X. Changes of isoflavone profile in the hypocotyls and cotyledons of soybeans during dry heating and germination. J. Agric. Food Chem. 2009, 57, 9002–9010. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Lee, J.H.; Vodovotz, Y.; Schwartz, S.J. Changes in distribution of isoflavones and β-glucosidase activity during soy bread proofing and baking. Cereal Chem. 2004, 81, 741–745. [Google Scholar] [CrossRef]
- Wang, H.J.; Murphy, P.A. Mass balance study of isoflavones during soybean processing. J. Agric. Food Chem. 1996, 44, 2377–2383. [Google Scholar] [CrossRef]
- Jackson, C.J.C.; Dini, J.P.; Lavandier, C.; Rupasinghe, H.P.V.; Faulkner, H.; Poysa, V.; Buzzell, D.; DeGrandis, S. Effects of processing on the content and composition of isoflavones during manufacturing of soy beverage and tofu. Process Biochem. 2002, 37, 1117–1123. [Google Scholar] [CrossRef]
- Yu, X.; Meenu, M.; Xu, B.; Yu, H. Impact of processing technologies on isoflavones, phenolic acids, and antioxidant capacities of soymilk prepared from 15 soybean varieties. Food Chem. 2021, 345, 128612. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Murphy, P.A. Isoflavone content in commercial soybean foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar] [CrossRef]
- Andrade, J.C.; Mandarino, J.M.G.; Kurozawa, L.E.; Ida, E.I. The effect of thermal treatment of whole soybean flour on the conversion of isoflavones and inactivation of trypsin inhibitor. Food Chem. 2016, 194, 1095–1101. [Google Scholar] [CrossRef]
- Murphy, P.A.; Barua, K.; Hauck, C.C. Solvent extraction selection in the determination of isoflavones in soy foods. J. Chromatogr. B Biomed. Appl. 2002, 777, 129–138. [Google Scholar] [CrossRef]
- Chien, J.T.; Hsieh, H.C.; Kao, T.H.; Chen, B.H. Kinetic model for studying the conversion and degradation of isoflavones during heating. Food Chem. 2005, 91, 425–434. [Google Scholar] [CrossRef]
- Huang, R.Y.; Chou, C.C. Heating affects the content and distribution profile of isoflavones in steamed black soybeans and black soybean koji. J. Agric. Food Chem. 2008, 56, 8484–8489. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Rani, A.; Dixit, A.K.; Bhatnagar, D.; Chauhan, G.S. Relative changes in tocopherols, isoflavones, total phenolic content, and antioxidative activity in soybean seeds at different reproductive stages. J. Agric. Food Chem. 2009, 57, 2705–2710. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Sasanuma, T.; Abe, T. Accumulation of gamma-aminobutyrate (GABA) caused by heat-drying and expression of related genes in immature vegetable soybean (edamame). Breed. Sci. 2013, 63, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonne, A.; Smith, M.; Weaver, D.B.; Vail, T.; Barnes, S.; Wei, C.I. Retention and changes of soy isoflavones and carotenoids in immature soybean seeds (Edamame) during processing. J. Agric. Food Chem. 2000, 48, 6061–6069. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.L.; Berhow, M.A.; Kim, J.T.; Chi, H.Y.; Lee, S.J.; Chung, I.M. Evaluation of soyasaponin, isoflavone, protein, lipid, and free sugar accumulation in developing soybean seeds. J. Agric. Food Chem. 2006, 54, 10003–10010. [Google Scholar] [CrossRef]
- Simonne, A.H.; Weaver, D.B.; Wei, C.I. Immature soybean seeds as a vegetable or snack food: Acceptability by American consumers. Innov. Food Sci. Emerg. Technol. 2000, 1, 289–296. [Google Scholar] [CrossRef]
- Rubel, A.; Rinne, R.W.; Canvin, D.T. Protein, oil, and fatty acid in developing soybean seeds. Crop Sci. 1972, 12, 739–741. [Google Scholar] [CrossRef]
- Gogoi, N.; Farooq, M.; Barthakur, S.; Baroowa, B.; Paul, S.; Bharadwaj, N.; Ramanjulu, S. Thermal stress impacts on reproductive development and grain yield in grain legumes. J. Plant Biol. 2018, 61, 265–291. [Google Scholar] [CrossRef]
- Sale, P.W.G.; Campbell, L.C. Patterns of mineral nutrient accumulation in soybean seed. Field Crop Res. 1980, 3, 157–163. [Google Scholar] [CrossRef]
- Im, M.H.; Choi, J.D.; Choi, K.S. The oxidation stability and flavor acceptability of oil from roasted soybean. J. Agric. Food Chem. Biotechnol. 1995, 38, 425–430. [Google Scholar]
- Duan, S.; Kwon, S.J.; Eom, S.H. Effect of thermal processing on color, phenolic compounds, and antioxidant activity of Faba bean (Vicia faba L.) leaves and seeds. Antioxidants 2021, 10, 1207. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Lee, J.H. Effects of oven-drying, roasting, and explosive puffing process on isoflavone distributions in soybeans. Food Chem. 2009, 112, 316–320. [Google Scholar] [CrossRef]
- Coward, L.; Smith, M.; Kirk, M.; Barnes, S. Chemical modification of isoflavones in soyfoods during cooking and processing. Am. J. Clin. Nutr. 1998, 68, 1486S–1491S. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Sakamota, A.; Takayagi, T.; Yokotsuka, K. Changes in isoflavone compositions of soybean foods during cooking process. Food Sci. Technol. 2000, 6, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Niamnuy, C.; Nachaisin, M.; Laohavanich, J.; Devahastin, S. Evaluation of bioactive compounds and bioactivities of soybean dried by different methods and conditions. Food Chem. 2011, 129, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, C.L.; Haddad, R.; Eberlin, M.N.; Carrão-Panizzi, M.C.; Tsai, S.M.; Park, Y.K. Thermal behavior of malonylglucoside isoflavones in soybean flour analyzed by RPHPLC/DAD and eletrospray ionization mass spectrometry. LWT 2012, 48, 114–119. [Google Scholar] [CrossRef]
- Kao, T.H.; Lu, Y.F.; Hsieh, H.C.; Chen, B.H. Stability of isoflavone glucosides during processing of soymilk and tofu. Food Res. Int. 2004, 37, 891–900. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, Q.; Godber, J.S. Stabilities of daidzin, glycitin, genistin, and generation of derivatives during heating. J. Agric. Food Chem. 2002, 50, 7402–7406. [Google Scholar] [CrossRef] [PubMed]
- Sakthivelu, G.; Akitha Devi, M.K.; Giridhar, P.; Rajasekaran, T.; Ravishankar, G.A.; Nikolova, M.T.; Angelov, G.B.; Todorova, R.M.; Kosturkova, G.P. Isoflavone composition, phenol content, and antioxidant activity of soybean seeds from India and Bulgaria. J. Agric. Food Chem. 2008, 56, 2090–2095. [Google Scholar] [CrossRef]
- Kim, J.J.; Kim, S.H.; Hahn, S.J.; Chung, I.M. Changing soybean isoflavone composition and concentrations under two different storage conditions over three years. Food Res. Int. 2005, 38, 435–444. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Hoffmann, M.; Carle, R. Thermal degradation kinetics of isoflavone aglycones from soy and red clover. Mol. Nutr. Food Res. 2006, 50, 373–377. [Google Scholar] [CrossRef]
- Mathias, K.; Ismail, B.; Corvalan, C.M.; Hayes, K.D. Heat and pH effects on the conjugated forms of genistin and daidzin isoflavones. J. Agric. Food Chem. 2006, 54, 7495–7502. [Google Scholar] [CrossRef] [PubMed]
- Lima, F.S.; Kurozawa, L.E.; Lda, E.L. The effects of soybean soaking on grain properties and isoflavones loss. LWT 2014, 59, 1274–1282. [Google Scholar] [CrossRef]
- Oh, Y.; Kim, K.; Yun, H.; Park, K.; Suh, S.; Moon, J.; Cho, S.; Kim, Y.; Kim, S.; Park, H.; et al. A new soybean cultivar, ”Pungwon” for sprout with disease resistance, lodging resistance and high yielding. Korean J. Breed. Sci. 2007, 39, 502–503. [Google Scholar]
- Lim, Y.J.; Lim, B.; Kim, H.Y.; Kwon, S.J.; Eom, S.H. Deglycosylation patterns of isoflavones in soybean extracts inoculated with two enzymatically different strains of lactobacillus species. Enzym. Microb. Technol. 2020, 132, 109394. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seed Maturity | Water Content (%) | Dry Weight (g/10 ea) | SSC (°Brix) |
|---|---|---|---|
| Immature | 66.87 ± 0.32 a | 0.66 ± 0.02 b | 2.75 ± 0.18 b |
| Mature | 13.04 ± 0.22 b | 0.88 ± 0.03 a | 15.76 ± 0.31 a |
| FD (0 Min) | Dry-Heating | Wet-Heating | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 Min | 60 Min | 90 Min | 120 Min | 30 Min | 60 Min | 90 Min | 120 Min | |||
| Malonyl glycosides | MDZI | 62.32 ± 1.54 a | 63.38 ± 9.37 a | 34.81 ± 3.21 b | 29.62 ± 3.55 b | 23.01 ± 2.31 b,c | 23.16 ± 2.64 b,c | 9.39 ± 1.27 c,d | 4.01 ± 0.08 d | 2.03 ± 0.34 d |
| MGLI | 29.26 ± 2.47 a | 26.09 ± 7.54 a | 18.51 ± 3.35 a,b | 10.51 ± 1.80 b,c | 7.70 ± 0.63 b,c | 5.73 ± 0.42 c | 1.57 ± 0.44 c | 0.62 ± 0.21 c | N.D. | |
| MGNI | 78.18 ± 0.21 a | 72.87 ± 9.58 a | 57.24 ± 4.00 a,b | 40.79 ± 6.04 b,c | 37.21 ± 4.76 b,c | 38.68 ± 2.06 b,c | 20.07 ± 1.80 c,d | 7.13 ± 0.83 d | 4.23 ± 0.48 d | |
| Acetyl glycosides | ADZI | 5.20 ± 0.29 a | 2.80 ± 0.62 b,c | 2.64 ± 0.40 b,c | 4.39 ± 0.70 a,b | 5.66 ± 0.70 a | 1.06 ± 0.20 c | 0.68 ± 0.05 c | N.D. | N.D. |
| AGLI | N.D. | Tr. | 0.36 ± 0.19 a | 2.56 ± 0.77 a | 3.65 ± 1.23 a | N.D. | N.D. | N.D. | N.D. | |
| AGNI | Tr. | 0.12 ± 0.12 c,d | 2.20 ± 0.26 c,d | 4.02 ± 1.83 b | 9.94 ± 1.27 a | 2.25 ± 0.25 c,d | 2.86 ± 0.13 c | 2.25 ± 0.17 c,d | 2.04 ± 0.16 c,d | |
| β-glycosides | DZI | 3.71 ± 0.53 c | 5.16 ± 1.17 b,c | 6.23 ± 0.59 b,c | 10.11 ± 2.81 a,b,c | 15.45 ± 1.80 a | 8.91 ± 1.47 a,b,c | 11.72 ± 1.51 a,b | 10.55 ± 1.02 a,b,c | 9.11 ± 0.70 a,b,c |
| GLI | 11.62 ± 1.16 a | 11.30 ± 3.98 a | 10.82 ± 1.37 a | 10.84 ± 2.27 a | 11.70 ± 1.50 a | 8.62 ± 1.08 a | 7.42 ± 1.12 a | 6.24 ± 0.80 a | 7.31 ± 0.48 a | |
| GNI | 8.08 ± 0.96 e | 10.26 ± 1.90 d,e | 14.30 ± 0.57 c,d,e | 22.52 ± 2.20 a,b,c | 30.16 ± 2.90 a | 19.16 ± 1.80 b,c,d | 26.12 ± 2.22 a,b | 25.47 ± 2.30 a,b | 19.9 ± 1.41 b,c | |
| Aglycones | DZE | 0.77 ± 0.12 a | 0.33 ± 0.03 a | N.D. | 0.61 ± 0.24 a | 0.75 ± 0.32 a | N.D. | N.D. | N.D. | N.D. |
| GLE | Tr. | 0.28 ± 0.14 a | 0.74 ± 0.42 a | 1.13 ± 0.25 a | 1.03 ± 0.42 a | N.D. | N.D. | N.D. | N.D. | |
| GNE | 0.22 ± 0.11 a,b | 0.70 ± 0.12 a,b | 0.74 ± 0.17 a,b | 0.87 ± 0.21 a | 0.89 ± 0.14 a | 0.07 ± 0.07 b | 0.22 ± 0.14 a,b | N.D. | N.D. | |
| FD (0 Min) | Dry-Heating | Wet-Heating | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 Min | 60 Min | 90 Min | 120 Min | 30 Min | 60 Min | 90 Min | 120 Min | |||
| Malonyl glycosides | MDZI | 119.20 ± 11.24 a | 68.96 ± 4.43 b | 57.43 ± 3.54 b,c | 47.09 ± 2.40 b,c,d | 31.92 ± 1.20 d,e | 48.00 ± 2.54 b,c,d | 35.03 ± 1.36 c,d,e | 20.91 ± 0.64 d,e | 18.34 ± 1.20 e |
| MGLI | 24.18 ± 5.96 a | 21.53 ± 1.52 a | 22.27 ± 2.22 a,b | 15.58 ± 2.61 b,c | 14.15 ± 1.38 b,c | 16.22 ± 2.80 b,c | 12.87 ± 2.84 c | 4.65 ± 0.28 c | 4.09 ± 0.22 c | |
| MGNI | 141.25 ± 7.92 a | 89.11 ± 3.38 b | 80.17 ± 3.10 b | 68.91 ± 2.22 b,c,d | 52.48 ± 3.44 d,e | 63.58 ± 0.59 b,c | 52.53 ± 0.48 c,d | 32.76 ± 1.20 e | 26.17 ± 0.76 e | |
| Acetyl glycosides | ADZI | 3.09 ± 0.35 d | 5.74 ± 0.56 d | 10.46 ± 0.61 b,c | 15.83 ± 1.97 a | 12.23 ± 0.98 a,b | 5.96 ± 0.29 d | 5.65 ± 0.30 d | 6.32 ± 0.11 d | 6.81 ± 0.35 c,d |
| AGLI | N.D. | 2.13 ± 0.83 c | 6.12 ± 1.48 b,c | 12.67 ± 1.04 a | 8.67 ± 2.17 a,b | 3.77 ± 0.66 b,c | 3.22 ± 0.65 b,c | 2.85 ± 0.25 c | 3.57 ± 0.66 b,c | |
| AGNI | Tr. | 5.62 ± 0.34 e | 14.51 ± 0.78 b | 21.10 ± 0.53 a | 20.58 ± 1.61 a | 6.57 ± 0.06 d,e | 9.09 ± 0.19 c,d | 9.42 ± 0.22 c,d | 11.84 ± 0.35 b,c | |
| β-glycosides | DZI | 7.56 ± 0.80 e | 11.22 ± 0.66 d,e | 19.53 ± 1.11 b | 19.92 ± 0.61 b | 19.07 ± 0.88 b | 13.62 ± 1.00 c,d | 17.97 ± 0.58 b,c | 20.03 ± 0.58 b | 27.45 ± 0.38 a |
| GLI | 7.53 ± 1.96 a,b | 7.71 ± 0.86 b | 15.05 ± 0.43 a,b | 14.47 ± 1.09 a,b | 10.37 ± 0.98 a,b | 14.82 ± 1.82 a,b | 17.30 ± 2.20 a | 13.97 ± 0.97 a,b | 16.75 ± 0.57 a | |
| GNI | 8.64 ± 1.99 e | 16.83 ± 0.58 d | 29.26 ± 1.04 c | 33.99 ± 0.87 b,c | 31.11 ± 2.08 b,c | 21.07 ± 0.18 d | 30.52 ± 0.71 b,c | 36.74 ± 0.91 b | 47.44 ± 1.62 a | |
| Aglycones | DZE | 1.21 ± 0.35 a | 0.50 ± 0.10 a,b | 0.78 ± 0.11 a,b | 0.63 ± 0.05 a,b | 0.59 ± 0.09 a,b | 0.38 ± 0.03 b | 0.29 ± 0.15 b | 0.31 ± 0.16 b | 0.57 ± 0.13 a,b |
| GLE | Tr. | N.D. | N.D. | N.D. | N.D. | 0.38 ± 0.02 a | N.D. | N.D. | N.D. | |
| GNE | 0.94 ± 0.14 a | 0.61 ± 0.08 a,b,c | 0.70 ± 0.07 a,b,c | 0.90 ± 0.12 a | 0.72 ± 0.04 a,b | 0.27 ± 0.06 c | 0.30 ± 0.02 b,c | 0.40 ± 0.10 b,c | 0.45 ± 0.10 b,c | |
| Seed Maturation | Heating Methods | MDZI | MGLI | MGNI | |
|---|---|---|---|---|---|
| Immature | D z | TIMG | 0.983 *** | 0.931 *** | 0.970 *** |
| W y | TIMG | 0.999 *** | 0.976 *** | 0.944 *** | |
| Mature | D | TIMG | 0.996 *** | 0.712 *** | 0.987 *** |
| W | TIMG | 0.996 *** | 0.889 *** | 0.997 *** | |
| ADZI | AGLI | AGNI | |||
| Immature | D | TIAG | 0.704 ** | 0.963 *** | 0.967 *** |
| W | TIAG | 0.902 *** | - | −0.537 * | |
| Mature | D | TIAG | 0.961 *** | 0.955 *** | 0.980 *** |
| W | TIAG | 0.901 ** | 0.844 *** | 0.983 *** | |
| DZI | GLI | GNI | |||
| Immature | D | TIG | 0.939 *** | 0.410 ns | 0.946 *** |
| W | TIG | 0.972 *** | −0.314 ns | 0.953 *** | |
| Mature | D | TIG | 0.943 *** | 0.769 *** | 0.956 *** |
| W | TIG | 0.983 *** | 0.687 ** | 0.976 *** | |
| DZE | GLE | GNE | |||
| Immature | D | TIA | −0.053 ns | 0.892 *** | 0.420 ns |
| W | TIA | 1.000 *** | - | - | |
| Mature | D | TIA | 0.601 * | - | 0.202 ns |
| W | TIA | 0.727 ** | - | 0.658 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, S.; Kwon, S.J.; Duan, S.; Lim, Y.J.; Eom, S.H. Isoflavone Changes in Immature and Mature Soybeans by Thermal Processing. Molecules 2021, 26, 7471. https://doi.org/10.3390/molecules26247471
Qu S, Kwon SJ, Duan S, Lim YJ, Eom SH. Isoflavone Changes in Immature and Mature Soybeans by Thermal Processing. Molecules. 2021; 26(24):7471. https://doi.org/10.3390/molecules26247471
Chicago/Turabian StyleQu, Shanshan, Soon Jae Kwon, Shucheng Duan, You Jin Lim, and Seok Hyun Eom. 2021. "Isoflavone Changes in Immature and Mature Soybeans by Thermal Processing" Molecules 26, no. 24: 7471. https://doi.org/10.3390/molecules26247471