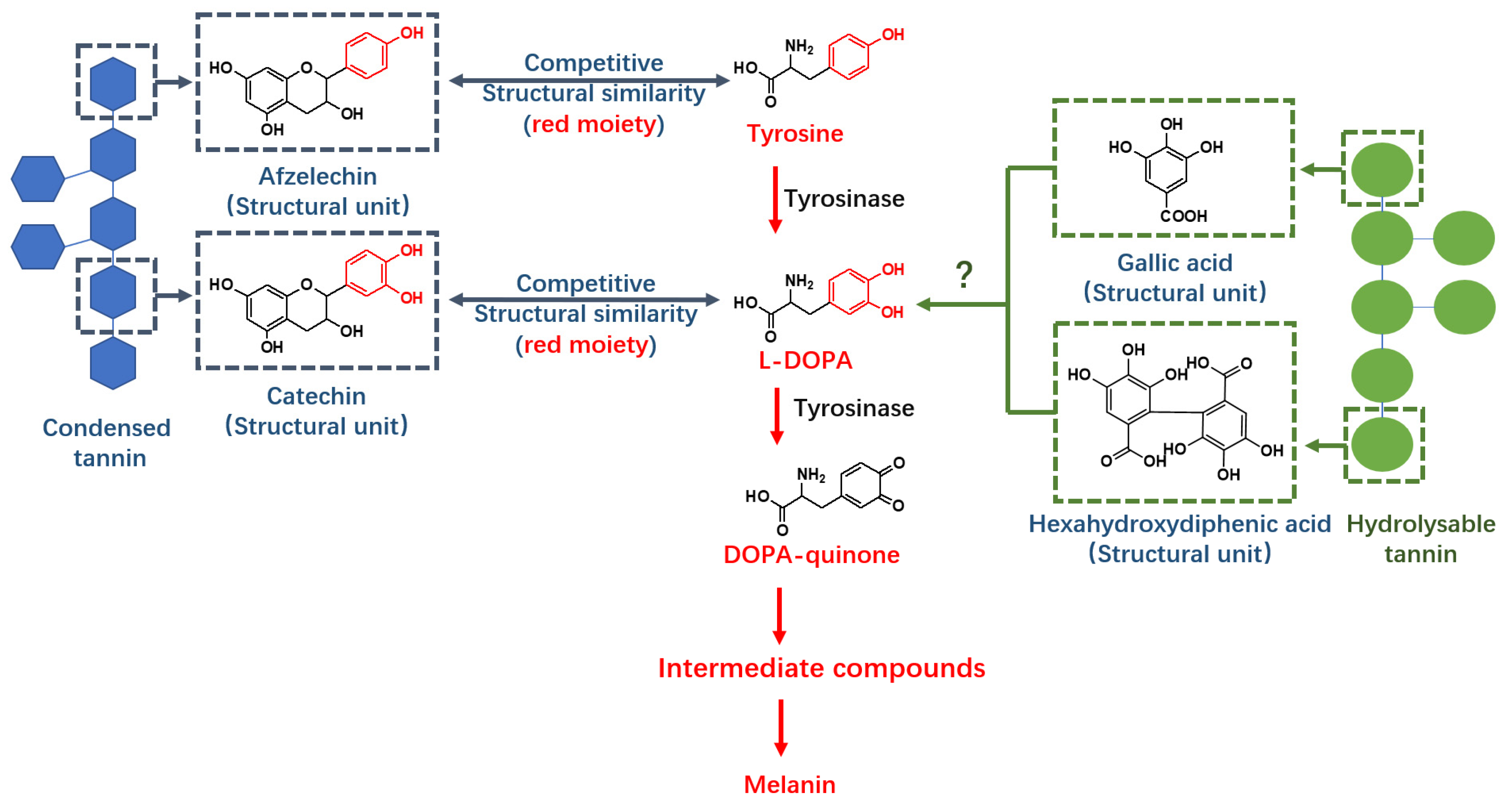
2.1. Structure Elucidation of Valonea Tannins
Tannin is a mixture of oligomers and polymers with similar structures and physical properties [
1]. Composition of structural moieties and molecular weights were considered as crucial structural characteristics because these characters have a substantial impact on the functions of tannins [
18]. The valonea tannin was purified with Sephadex LH-20 because it was proven previously that this method can eliminate the simple phenolics, proteins and polysaccharides efficiently, therefore it commonly used for tannin purification [
19]. Then, valonea tannin structure was elucidated using
13C NMR (
Figure S1) to obtain the composition of the structural moieties. Chemical shifts were compared with those of standard tannic and gallic acids as well as those of similar moieties from previous reports [
20] to give the proposed structure of valonea tannin based on the previous 1D and 2D-NMR research of ellagitannins and isolated flavonoid oligomers; resonances from 170 to 55 ppm were assigned to the carbons on carbonyl, phenyl and alkyl moieties, respectively, and shown on
Table 1.
Resonances from 100 to 170 ppm were used to identify the composition of hydrolysable tannins [
20]. The chemical shifts appeared at 164.41, 158.50, 114.89 and 113.63 ppm were attributed to the carbonyl C=O and phenolic C-C bridges on hexahydroxydiphenoyl (HHDP) moieties [
20]. These shifts can be considered as direct evidence to prove the existence of HHDP in valonea tannins. It also implied that valonea tannin belonged to the typical ellagitannin in the hydrolysable tannin family. This finding is in agreement with the report from Özgünay et al., who also found the HHDP moieties based on precursor and fragments ions in MALDI TOF MS spectrum of the valonea tannin [
21].
Resonances at 163.98, 144.67 and 109.61 ppm were attributed to the carbonyl C=O, phenolic –OH and phenolic carbon on galloyl moieties [
20]. Similarly, these shifts can be also found in gallic acid (169.1, 144.9 and 109.0 ppm) and tannic acid (169.7, 144.9 and 109.1 ppm).
The resonances from 70 to 54 ppm, were characteristic resonances from the open glucose forms [
22], while the closed glucose form (95 to 80 ppm) was not detected in the current study. Other resonances were attributed to the phenolic carbon linked with or without hydroxyl groups and were attributed to both HHDP and galloyl moieties [
20].
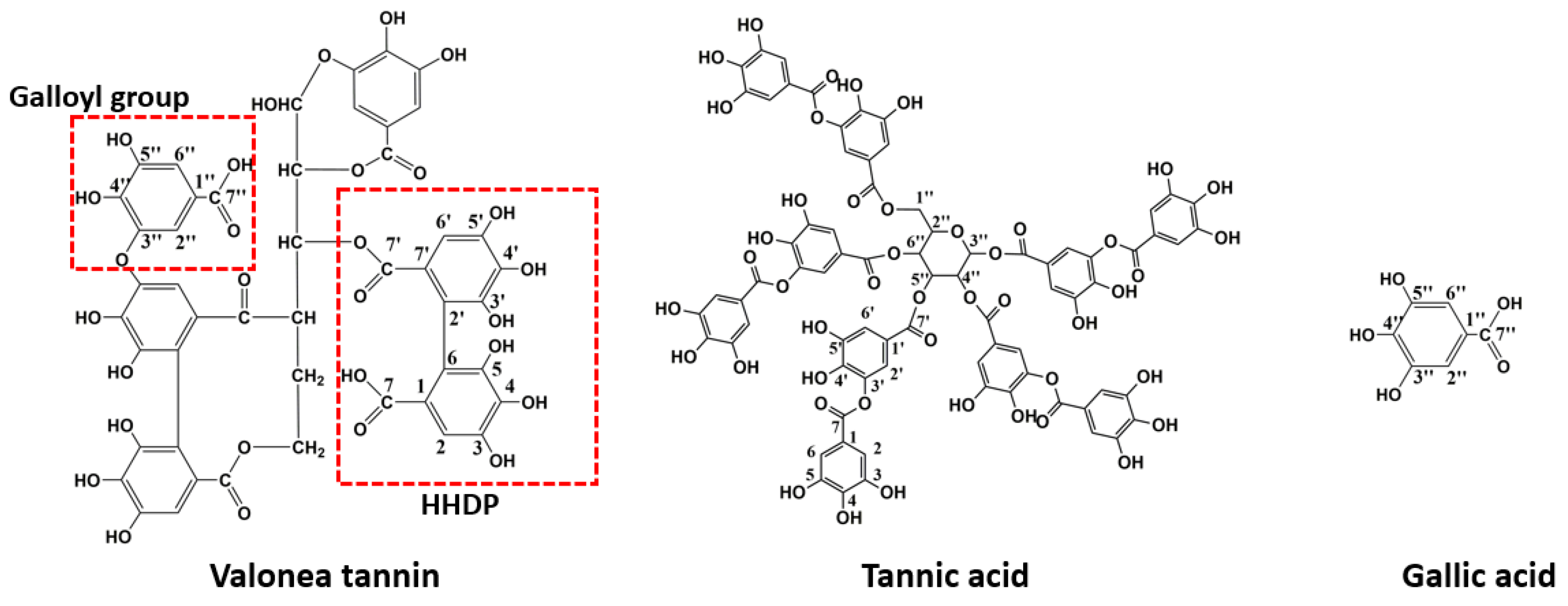
These chemical shifts not only confirmed the existence of galloyl and HHDP moieties in valonea tannins, but also indicated the structural characteristics of the valonea tannins were significantly different to condensed tannins (composed of flavanol subunits) [
23]. Based on the information obtained from
13C NMR analysis, the typical structure of the valonea tannin was deduced and shown on
Figure 2, accompanied with the structure of tannic acid and gallic acid.
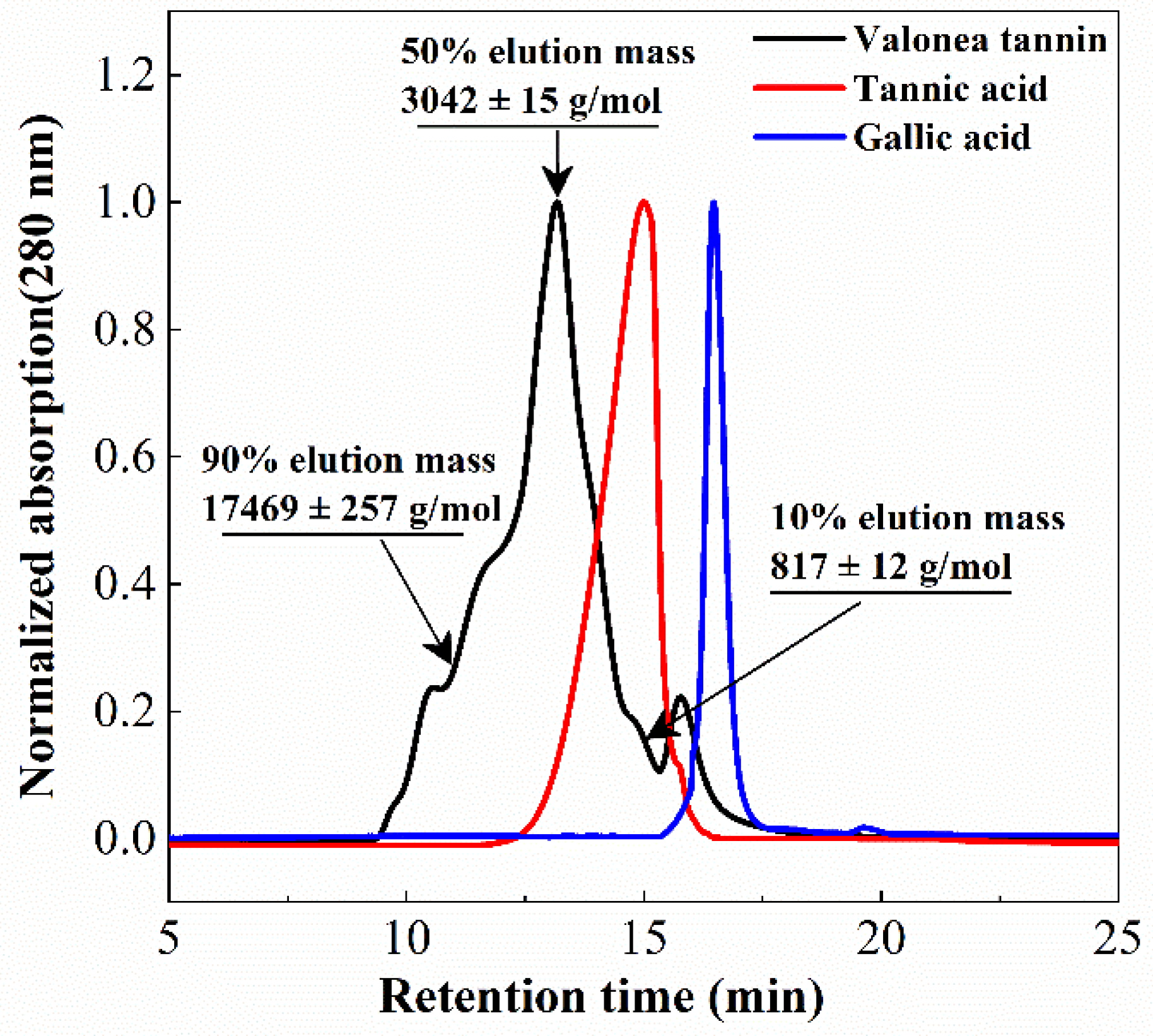
Tannin molecular weight is directly related to the biochemical properties and is a crucial parameter for protein interaction and enzyme inhibition [
15]. Determining the average molecular weight of a tannin can also assist in elucidating the average size of the tannins [
15,
18]. The molecular weight distribution of the isolated valonea tannin was analysed using gel permeation chromatography (GPC) (
Figure 3).
Based on the GPC results, 10%, 50% and 90% elution mass of the valonea tannins were 817 ± 12, 3042 ± 15 and 17,469 ± 257 g/mol, respectively. This indicated that 80% of the tannin molecules had a mass ranging from 817 to 17,469 g/mol. Furthermore, 50% valonea tannin molecules eluted earlier than tannic acid and 90% of the valonea tannin showed higher molecular weight than gallic acid. This indicated the average molecular weight of valonea tannin should be higher than the other two compounds. Based on the results from Kennedy’s report, 50% elution mass of tannin is in good agreement with the average molecular weight determined by other method [
19]. Therefore, the 50% elution mass (3042 ± 15 g/mol) of the valonea tannin was taken for the following analysis.
2.2. Tyrosinase Inhibition Activity of the Valonea Tannin
Valonea tannin was assessed for tyrosinase inhibition to determine if this natural product may have value for commercial application as a whitening agent in cosmetic products or dermatologic medicines. Tannic acid and gallic acid were also assessed for tyrosinase inhibition for comparison as these compounds were: (1) all composed of pyrogallol moieties (
Figure 2) that played critical roles in the enzyme inhibition processes [
9]; (2) have molecular weights that differ to each other and are lower than valonea tannins (
Figure 3); and (3) accepted as tyrosinase inhibitors with better performances than the commercial tyrosinase inhibitors [
9].
The tyrosinase inhibition activities were assessed across a range of concentrations for valonea tannins as well as tannic and gallic acids. Inhibition was measured as an increase in absorbance over time (
Figure S2). The IC
50 values were calculated as the inhibitor concentration inducing a 50% reduction in maximum absorbance and are presented in
Table 2.
The IC
50 values of the involved inhibitors followed a trend: valonea tannin < tannic acid < gallic acid. This suggested that tyrosinase inhibition capability was, positively related to molecular weight, which is also in agreement with a conclusion obtained by condensed tannins with different molecular weights [
15].
Gallic acid has previously been shown to have excellent tyrosinase inhibition ability with an IC
50 value 100-fold lower than kojic acid [
24]. In the current study, the IC
50 value of valonea tannin was 339-fold lower than gallic acid (
Table 2), which indicated that the valonea tannin may has better tyrosinase inhibition ability than the commercial inhibitors. To explore why the valonea tannin showed such an outstanding tyrosinase inhibition ability, the inhibition mechanisms were studied and presented in the following sections.
2.3. The Kinetic Study of Tyrosinase Inhibition Mechanism
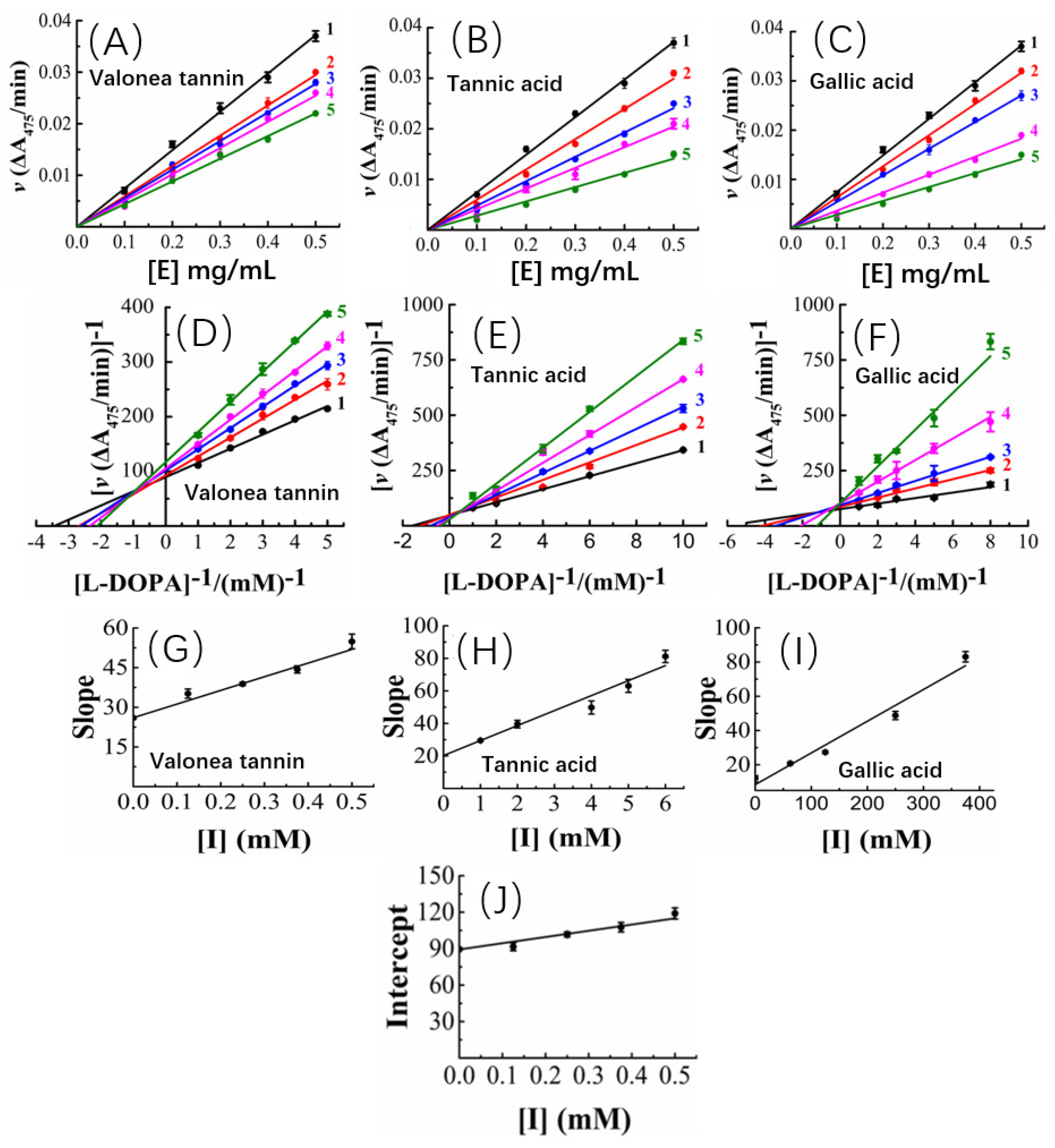
To answer why valonea tannin has inhibition ability, kinetic characteristics of the inhibition reactions were studied to better understand the mechanism of action of valonea tannin’s tyrosinase inhibition. The kinetic studies were applied and the results are provided in
Figure 4.
The tyrosinase concentration-reaction rate plots showed that, with a constant valonea tannin concentration, the initial velocities linearly fitted with tyrosinase concentrations and the fitted lines all passed through origin of the axis (
Figure 4A). Therefore, a deduction can be made as: despite varying valonea tannin concentrations, the reactions will continue proceeding until tyrosinase is eliminated completely from the reaction systems ([E] = 0). Based on the characteristics mentioned above, referencing the previous reports [
16,
25], the valonea tannin inhibition reaction was classified as a reversible type. As for tannic acid or gallic acid, their tyrosinase concentration-reaction rate plots also showed significant characteristics of the reversible type as well (
Figure 4B,C), in agreement with the results obtained by previous research [
26].
Based on the adjusted Michaelis–Menten equation [
27], the Lineweaver–Burk plots of the reactions were obtained by analysing velocity under different valonea tannin concentrations (
Figure 4D). It was evident that the inverse of the velocity (1/v) were all linearly fitted with the inverse of the substrate concentration (R
2 and
p-values are shown in
Table S4) while meanwhile it intersected in the second quadrant of the plot. Based on Waldrop’s finding, these graphical characters indicated the maximum reaction speed (Vmax = 1/intersect on y-axis) and Michaelis’s constant (K
M) of the reactions were all affected with varying inhibitor concentrations [
27]. Thus, the inhibition provided by valonea tannins belonged to a competitive−noncompetitive mixed type. In other words, the valonea tannin presents inhibition through either entering the active centre or binding on the surface (non-active centre) of tyrosinase. This is similar to the inhibition mechanism provided by condensed tannins [
16]. In contrast to valonea tannins, the Lineweaver–Burk plots of the tannic acid and gallic acid showed different characteristics (
Figure 4E,F). The fitted lines were intersected on y-axis, which indicated these compounds inhibit tyrosinase competitively, which is in agreement with a previous report [
28].
Slopes and y-axis intercepts of the fitted Lineweaver–Burk lines were taken for secondary fittings, where they all were linearly fitted with inhibitor concentrations (
Figure 4G–I). The slopes and y-axis intercepts of the new lines were calculated to obtain the inhibition constants of inhibitor-tyrosinase (K
I) and inhibitor–tyrosinase–substrate complexes (K
IS) (
Figure 4J), respectively [
27]. K
I is the dissociation constant between inhibitor and enzyme, and K
IS is the dissociation constant between inhibitor and the enzyme–substrate complex. The smaller the K
I or K
IS values provided by inhibitor, the better competition abilities to substrate were indicated [
27].
In the current research, the K
I and K
IS values of the involved inhibitors followed a trend as: valonea tannin (K
I) < valonea tannin (K
IS) < tannic acid (K
I) < gallic acid (K
I) (
Table 3). Indicated in comparison with tannic acid and gallic acid, the valonea tannin showed better competition ability than substrate. Furthermore, the higher the molecular weight of tested inhibitors, the better the competition ability that was shown. This trend is not only in agreement with the results observed from IC
50, but also similar to the conclusions that were obtained from condensed tannins [
29]. The results obtained from the kinetic study are summarized on
Table 3.
To find more information to explain why valonea tannin showed a better competition ability to substrate, the mechanism of valonea tannin–tyrosinase interactions were investigated using fluorescence and isothermal titration calorimetry (ITC) as described below.
2.4. Fluorescence and Thermodynamic Characters of the Valonea Tannin-Tyrosinase Interaction
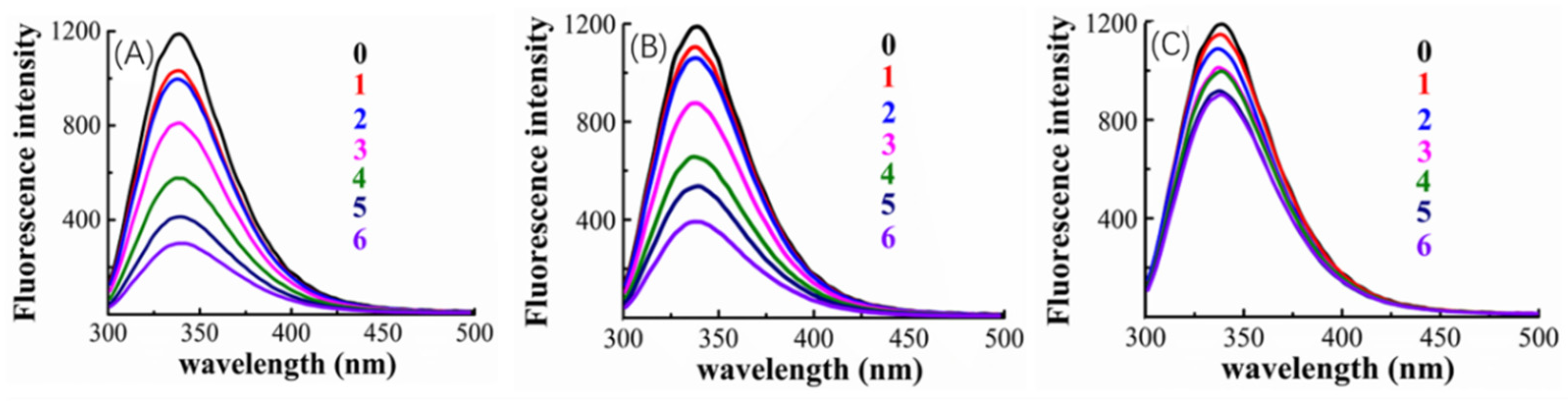
The cause of the observed improved competition ability of valonea tannin was explored using fluorescence quenching (
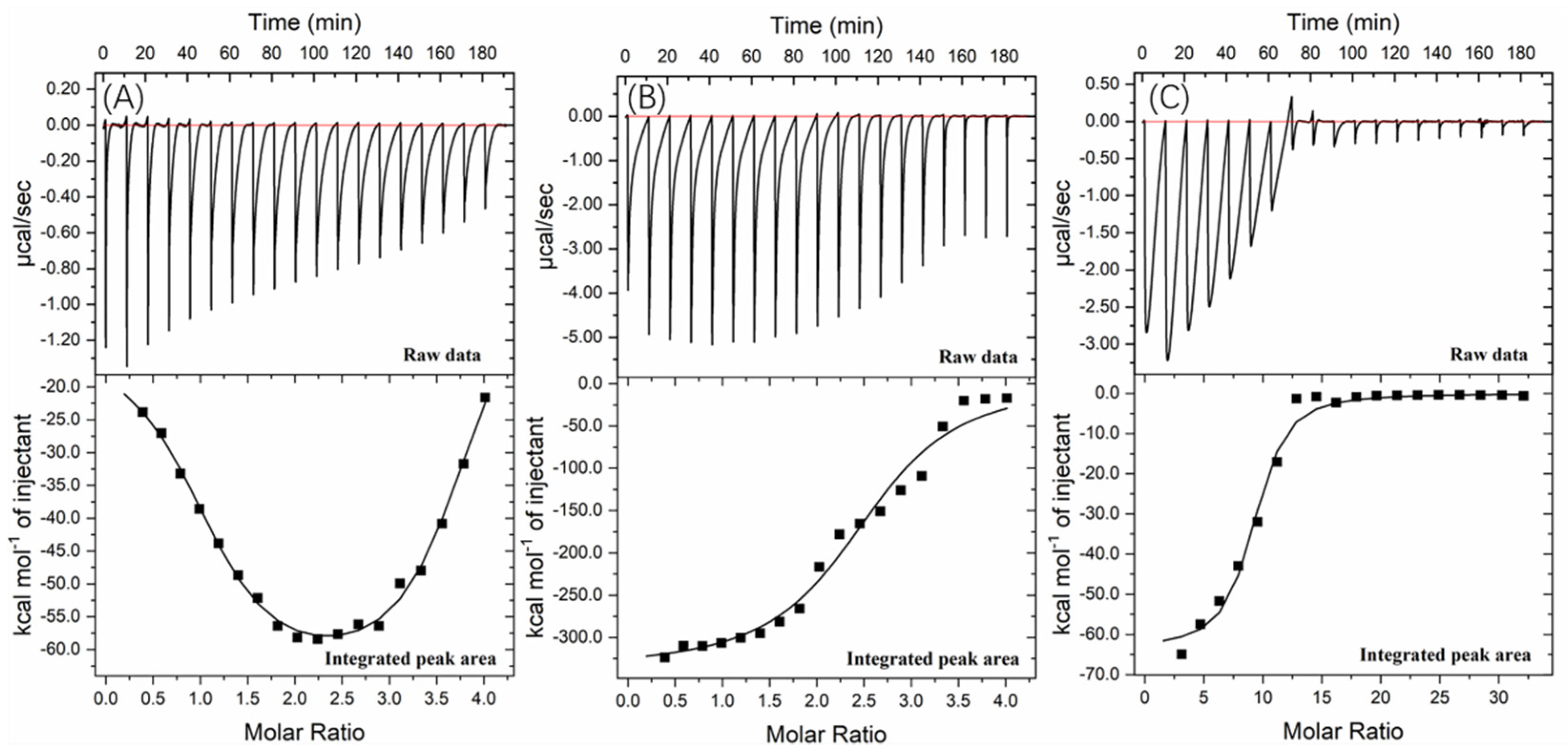
Figure 5) and isothermal titration calorimetry (ITC) (
Figure 6).
The tryptophan inside tyrosinase is the fluorophore that has florescence emitting with a 280 nm excitation wavelength [
30]. The tyrosinase–inhibitor binding process may lead to a fluorescence quenching induced by the structural changes of the tyrosinase around tryptophan, consequently, fluorescence quenching was used to provide primary information about inhibitor–tyrosinase interaction [
30].
Figure 5 indicated that, the tyrosinase showed a florescence emission at 340 nm, and fluorescence quenching appeared after adding inhibitors (valonea tannin, tannic acid or gallic acid, respectively). Greater concentrations of inhibitors also increased the observed quenching, which implies that the quenching may be induced by the interaction between inhibitors and tyrosinase.
The quenching phenomenon can be classified into dynamic and static types, while the former one does not involve chemical interaction between fluorophore and quencher, only the static quenching refers to binding interactions [
29]. Based on the study provided by Van De Weert and Stella [
29], the Stern–Volmer quenching rate constant (K
q) and apparent binding constant (K
a) were consequently obtained by linear regressions of the Stem–Volmer equation and double log Stem–Volmer equation (
Figure S3) and shown in
Table 4.
The fluorescence analysis results showed that K
q presented by valonea tannin–tyrosinase complex was significantly higher than the maximum scatter collision quenching rate constant for dynamic quenching (2 × 10
10 L/mol/s) [
29], meanwhile their maximum emission wavelengths were observed as constant at 340 nm (
Table 4). This phenomenon implied a non-covalent bond was formed between valonea tannin and tyrosinase [
29], which is in agreement with the results obtained through hydrolysable tannin–protein interactions [
31]. As for the tannic acid and gallic acid, the tyrosinase showed similar quenching characters. This further suggests that these interactions were non-covalent as previously reported for interactions between tyrosinase and tannic acid [
32]. The quenching efficiency as well as the K
a of the involved inhibitors followed: valonea tannin > tannic acid > gallic acid. These results preliminary indicated the valonea tannin maybe more strongly bound to tyrosinase than the other two inhibitors [
29].
Further information about the valonea tannin–tyrosinase binding process was further investigated using ITC analysis. The tyrosinase is a typical globular protein which has complicated stereochemical structures, and tannin is a natural product with polydispersity, therefore tannin–protein interactions are believed to be complex [
33]. ITC enables measurement of the binding strength of tannin–protein interactions by quantifying the thermodynamic changes of complex reactions [
34]. The ITC curves of valonea tannin, tannic acid and gallic acid titrated into tyrosinase are shown in
Figure 6. These negative peaks, shown on the raw data, were attributed to the negative enthalpy from tannin–tyrosinase interaction. In other words, the interaction was exothermic [
33]. These titration signals were typical of protein–ligand interactions, resulting from the combination of hydrogen bonding and hydrophobic interactions as has been previously seen in the initial section of ITC curves from earlier reports [
34].
The rapid reduction of exothermic peaks (ΔH < 0) upon titration indicated the valonea tannins showed a strong affinity to tyrosinase. With increasing valonea tannin addition, the number of available binding sites decreased, which resulted in the associated enthalpy changes decreasing. The titration curve was found to better fitted with the “two sets of binding sites” model, which implied the two kinds of binding sites were occupied by valonea tannins. On the contrary, the titration curves obtained from tannic acid and gallic acid showed better fitted with the “one set of binding sites” model. The inhibition mechanism results classified valonea tannin as a competitive–noncompetitive mixed type, while classified gallic acid and tannic acid were competitive types (
Table 3). This also indicated the valonea tannin bound on both active and non-active sites of the tyrosinase, while gallic acid and tannic acid bound on the active sites further confirming the proposed inhibition mechanism.
Enthalpy change (ΔH) was calculated from the area under each ITC curve, and entropy changes (TΔS), stoichiometry (molar ratio of inhibitor to site, expressed as “n” [
34]) and equilibrium binding constants (K) were calculated from the graphical characters of the curve using the Origin 7.0 software package. Results are shown in
Table 5. The negative ΔH values of valonea tannin–tyrosinase interaction indicating that interactions on these two sites were exothermic. The negative ΔH and TΔS, observed from inhibitor–tyrosinase complexes, demonstrate both hydrophobic interactions and hydrogen bonding were essential for the binding [
34].
The valonea tannin showed K values 70.6 ± 3.0 (×10
5) at binding site 1 and 4.3 ± 0.8 (×10
5) at binding site 2, which were significantly higher than the tannic or gallic acids, suggesting stronger binding. This is likely to relate to the greater molecular weight of the valonea tannin that enabled quicker and stronger inhibition to tyrosinase. As expected, the n values of valonea tannins were smaller than (or similar to) tannic acid and gallic acid. This phenomenon could be attributed to the stronger steric hinderance, larger hydrodynamic volumes and higher binding strength presented by valonea tannin, which were obtained from GPC, inhibition kinetic and fluorescence study. This result supports the trend observed in apparent binding constant (K
a, from Stern–Volmer quenching) and inhibition constant in enzyme−substrate complex (K
IS, from Michaelis–Menten kinetic) in this study, as well as a previous hypotheses that considers molecular weight as a key influence of tannin–protein interaction [
35].
2.5. Molecular Docking Analysis
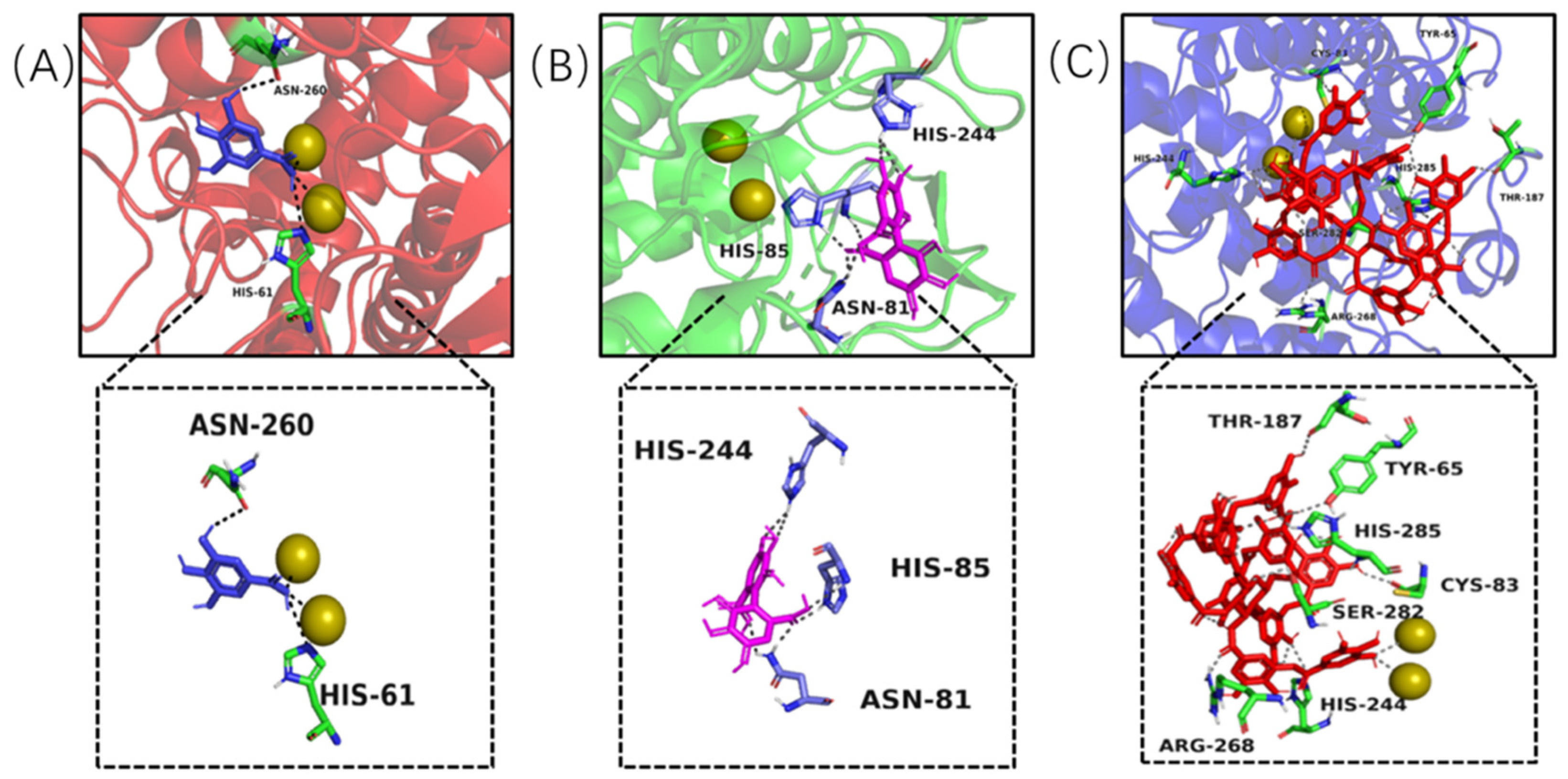
Molecular docking modelling was performed to provide insight to the specific tyrosinase inhibition activity of valonea tannin. As revealed by
13C NMR, the valonea tannins were composed of galloyl and HHDP as structural units. Thereby the gallic acid and HHDP were chosen as ligand models and applied for molecular docking analysis. For the docking results from each ligand-tyrosinase interaction, 20 complexes with different docking poses (also with lowest docking energies) were collected, among which the complexes with lowest energies are shown on
Figure 7.
Molecular docking results indicated the model molecules (including gallic acid and HHDP) bind with tyrosinase at the active centre via hydrogen bonding [
36]. Two hydrogen bonds were observed (Asn260 and His61) in the gallic acid–tyrosinase complex. As for HHDP, six hydrogen bonds were formed via His85, Asn81 and His244 inside the active centre. As for the tannic acid–tyrosinase complex, more hydrogen bonds were observed (
Figure 7C). Thr187, His285, Cys83, His244, Ser282, Tyr65 and Arg268 were shown participating in the interaction; meanwhile eight hydrogen bonds were formed.
The results from molecular docking not only visualized the active site bindings which were observed from ITC and fluorescence analyses, but also discovered the interaction between copper ions and gallic or tannic acid. The valence states of these copper ions not only determine the type of activity showed by tyrosinase (diphenol or monophenol) but also believed to influence the activity of tyrosinase in catalytic cycles [
37]. Therefore, the copper ion chelating and antioxidant ability of the valonea tannin were assessed and presented in the following section.
2.6. Copper Ion Chelating and Antioxidant Abilities
To further explore the inhibition mechanism proposed by the molecular docking studies, copper chelation and antioxidant analyses were also conducted (
Table 6).
The adjacent hydroxyl groups on galloyl moieties, enabled the hydrolysable tannin to chelate with metal ions and form aggregates and eventually precipitate [
35]. Copper ion chelating ability was analysed using inductively coupled plasma (ICP) spectroscopy and shown in
Table 6. The results indicated that, after reacting with valonea tannin, large quantities of copper ions were precipitated and only 56.61 ± 0.66% were remaining in the aqueous phase. This implied chelating with copper ion was another pathway of tyrosinase inhibition, in agreement with the phenomenon observed through molecular docking.
Chemicals with antioxidant activities are known to also have tyrosinase inhibition activities, not only because the tyrosinase catalysed L-DOPA to DOPA quinone reaction is basically an oxidation reaction, but also because the antioxidants may have an impact on the oxidation state of copper ions thereby becoming a hindrance in catalytic cycles of tyrosinase [
37]. The DPPH
· and ABTS
·+ scavenging abilities of valonea were compared to that of a known antioxidant, ascorbic acid (
Table 6). Valonea tannin provided significantly lower IC
50 values than ascorbic acid, which indicated substantial antioxidant capability that may also contribute to the tyrosinase inhibition ability, as previously noted for ascorbic acid [
38].
In the current study, the tyrosinase inhibition activity of the inhibitors followed: valonea tannin > tannic acid > gallic acid. This trend repeatedly appeared in the results observed from tyrosinase binding analysis, as for the antioxidant and copper chelating abilities, the valonea tannin still showed better performance than the gallic acid and ascorbic acid but was similar to tannic acid (
Table 6). Therefore, the outstanding tyrosinase inhibition provided by valonea tannins is basically attribute to its tyrosinase binding ability.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}