
Antimicrobial Polyketide Metabolites from Penicillium bissettii and P. glabrum

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Evaluation of Extract Bioactivity
2.2. Derivatisation of Penicillic Acid
2.3. 13C NMR Study of Penicillic Acid (1) Biosynthesis
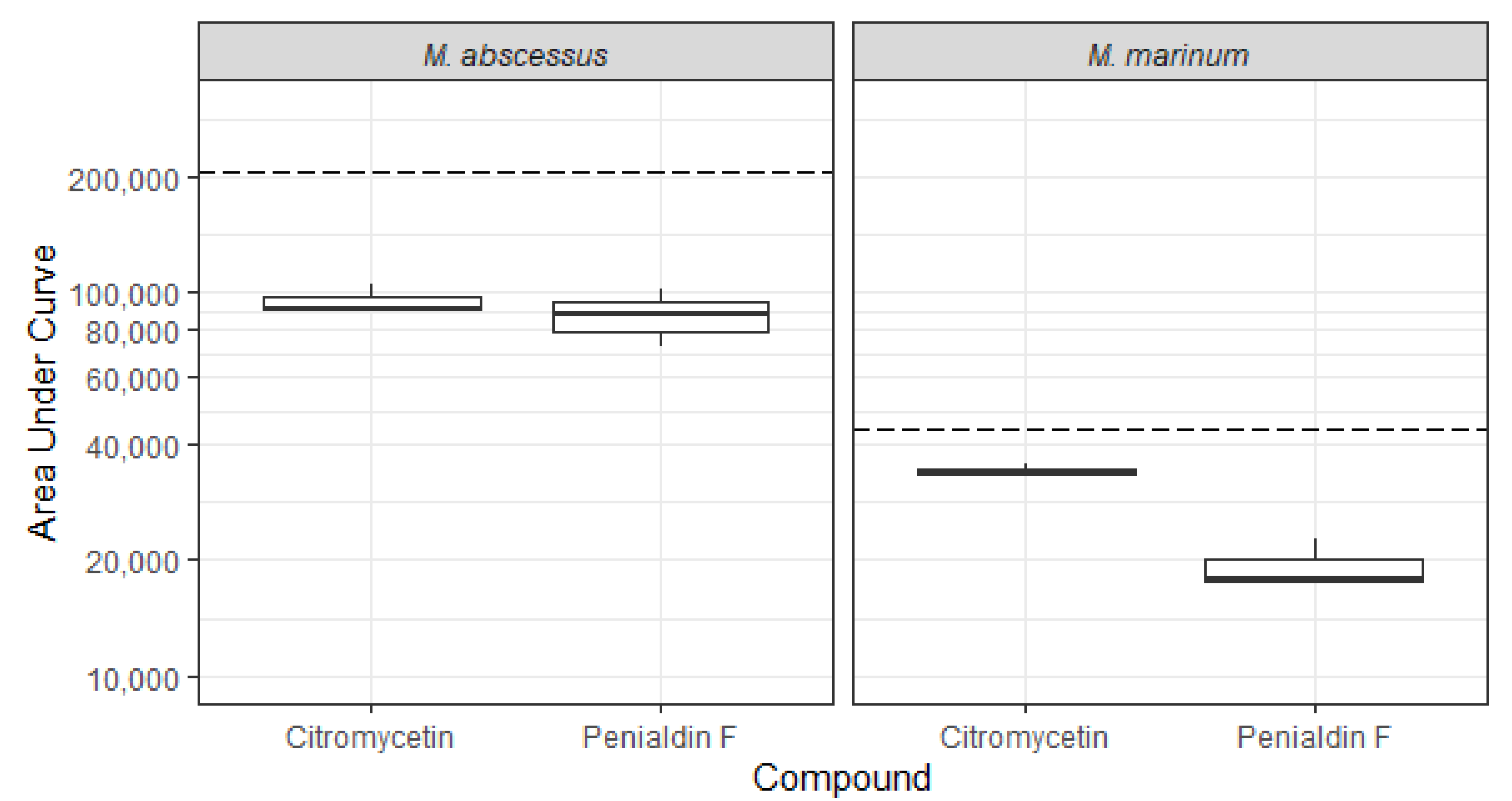
2.4. Bioactivity
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fungal Material
3.3. Antimicrobial Testing of Fungal Cultures
3.4. Extraction and Isolation
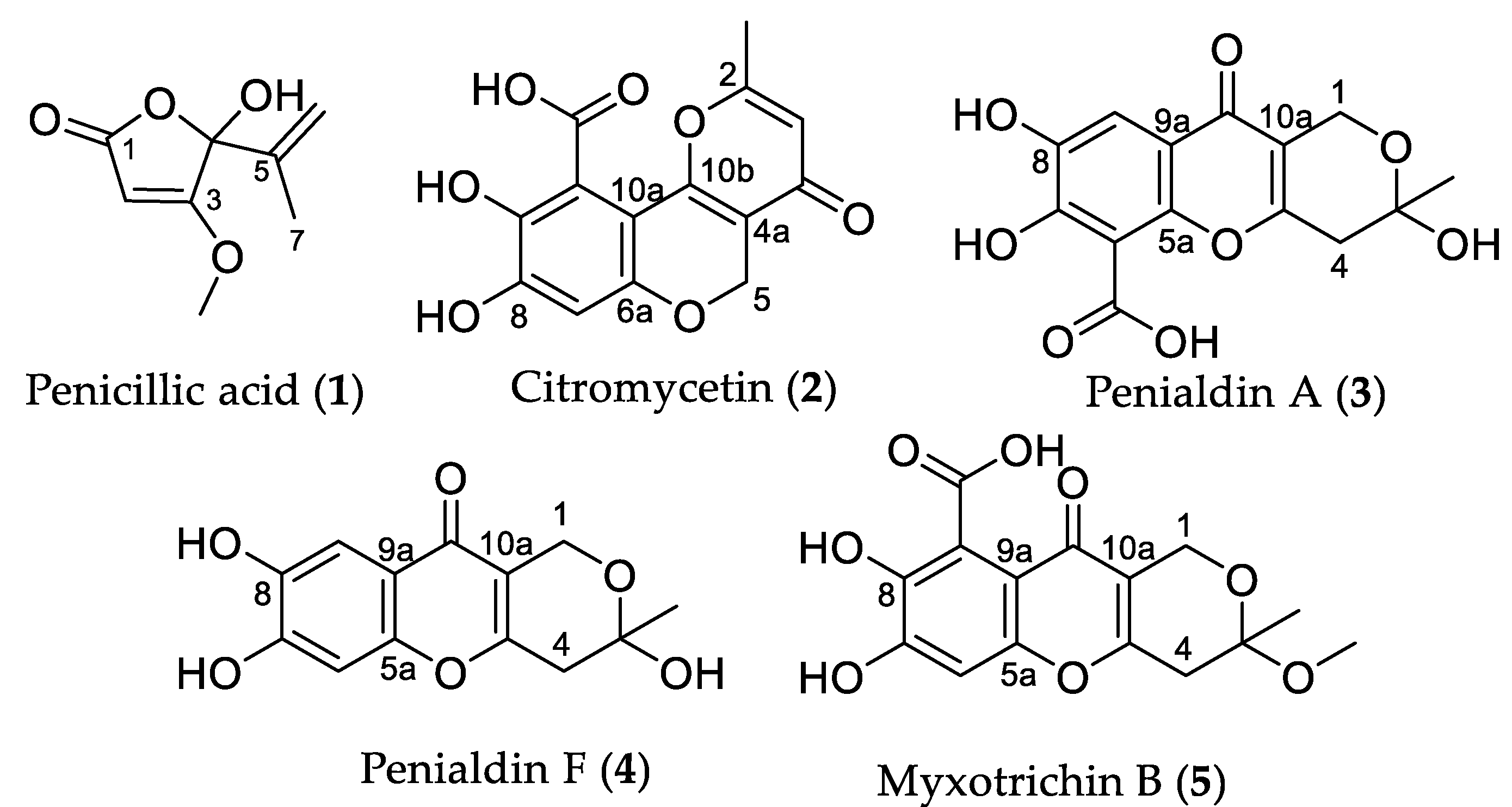
3.4.1. Penicillic acid (1)
3.4.2. Citromycetin (2)
3.4.3. Penialdin A (3)
3.4.4. Penialdin F (4)
3.4.5. Myxotrichin B (5)
3.4.6. (E)-3-Methoxy-5-methyl-4-oxohex-2-enoic acid (1a)
3.4.7. 5-Hydroxy-5-isopropyl-4-methoxyfuran-2(5H)-one (1b)
3.4.8. 3-Methoxy-5-oxo-2-(prop-1-en-2-yl)-2,5-dihydrofuran-2-yl acetate (1c)
3.5. Sodium [1-13C] Acetate Incorporated Acetate Fermentation of P. bissettii
Extraction and Isolation
3.6. Antimicrobial Assays of Pure Compounds
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Johnston, P.R.; Weir, B.S.; Cooper, J.A. Open Data on Fungi and Bacterial Plant Pathogens in New Zealand. Mycology 2017, 8, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Lobanovska, M.; Pilla, G. Penicillin’s Discovery and Antibiotic Resistance: Lessons for the Future? Yale J. Biol. Med. 2017, 90, 135–145. [Google Scholar] [PubMed]
- Houbraken, J. Fleming’s Penicillin Producing Strain Is Not Penicillium Chrysogenum but P. Rubens. Ima Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.-B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and Nomenclature of the Genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Houbraken, J.; Visagie, C.M.; Meijer, M.; Frisvad, J.C.; Busby, P.E.; Pitt, J.I.; Seifert, K.A.; Louis-Seize, G.; Demirel, R.; Yilmaz, N.; et al. A Taxonomic and Phylogenetic Revision of Penicillium Section Aspergilloides. Stud. Mycol. 2014, 78, 373–451. [Google Scholar] [CrossRef] [Green Version]
- Visagie, C.M.; Renaud, J.B.; Burgess, K.M.N.; Malloch, D.W.; Clark, D.; Ketch, L.; Urb, M.; Louis-Seize, G.; Assabgui, R.; Sumarah, M.W.; et al. Fifteen New Species of Penicillium. Pers. Int. Mycol. J. 2016, 36, 247–280. [Google Scholar] [CrossRef] [Green Version]
- Pohland, A.E.; Schuller, P.L.; Steyn, P.S.; Van Egmond, H.P. Physicochemical data for some selected mycotoxins. Pure Appl. Chem. 1982, 54, 2219–2284. [Google Scholar] [CrossRef]
- Li, H.-J.; Cai, Y.-T.; Chen, Y.-Y.; Lam, C.-K.; Lan, W.-J. Metabolites of Marine Fungus Aspergillus sp. Collected from Soft Coral Sarcophyton tortuosum. Chem. Res. Chin. U. 2010, 26, 415–419. [Google Scholar]
- Capon, R.J.; Stewart, M.; Ratnayake, R.; Lacey, E.; Gill, J.H. Citromycetins and Bilains A–C: New Aromatic Polyketides and Diketopiperazines from Australian Marine-Derived and Terrestrial Penicillium Spp. J. Nat. Prod. 2007, 70, 1746–1752. [Google Scholar] [CrossRef]
- Jouda, J.-B.; Kusari, S.; Lamshöft, M.; Mouafo Talontsi, F.; Douala Meli, C.; Wandji, J.; Spiteller, M. Penialidins A–C with Strong Antibacterial Activities from Penicillium Sp., an Endophytic Fungus Harboring Leaves of Garcinia nobilis. Fitoterapia 2014, 98, 209–214. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, L.; Wang, Q.; Ding, W.; Chen, Z.; Ma, Z. New Brefeldins and Penialidins from Marine Fungus Penicillium Janthinellum DT-F29. Nat. Prod. Res. 2018, 32, 282–286. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, H.-Y.; Wu, C.-S.; Jiao, Y.; Li, M.; Wang, Y.-Y.; Wang, S.-Q.; Zhao, Z.-T.; Lou, H.-X. Austdiol, Fulvic Acid and Citromycetin Derivatives from an Endolichenic Fungus, Myxotrichum Sp. Phytochem. Lett. 2013, 6, 662–666. [Google Scholar] [CrossRef]
- Kumla, D.; Pereira, J.A.; Dethoup, T.; Gales, L.; Freitas-Silva, J.; Costa, P.M.; Lee, M.; Silva, A.M.S.; Sekeroglu, N.; Pinto, M.M.M.; et al. Chromone Derivatives and Other Constituents from Cultures of the Marine Sponge-Associated Fungus Penicillium Erubescens KUFA0220 and Their Antibacterial Activity. Mar. Drugs 2018, 16, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ruan, Q.; Jiang, S.; Qu, Y.; Chen, J.; Zhao, M.; Yang, B.; Liu, Y.; Zhao, Z.; Cui, H. Cytochalasins and Polyketides from the Fungus Diaporthe Sp. GZU-1021 and Their Anti-Inflammatory Activity. Fitoterapia 2019, 137, 104187. [Google Scholar] [CrossRef]
- Fu, J.; Hu, L.; Shi, Z.; Sun, W.; Yue, D.; Wang, Y.; Ma, X.; Ren, Z.; Zuo, Z.; Peng, G.; et al. Two Metabolites Isolated from Endophytic Fungus Coniochaeta Sp. F-8 in Ageratina Adenophora Exhibit Antioxidative Activity and Cytotoxicity. Nat. Prod. Res. 2021, 35, 2840–2848. [Google Scholar] [CrossRef]
- Gou, X.; Tian, D.; Wei, J.; Ma, Y.; Zhang, Y.; Chen, M.; Ding, W.; Wu, B.; Tang, J. New Drimane Sesquiterpenes and Polyketides from Marine-Derived Fungus Penicillium Sp. TW58-16 and Their Anti-Inflammatory and α-Glucosidase Inhibitory Effects. Mar. Drugs 2021, 19, 416. [Google Scholar] [CrossRef] [PubMed]
- Alsberg, C.L.; Black, O.F. Contributions to the Study of Maize Deterioration: Biochemical and Toxicological Investigations of Penicillium Puberulum and Penicillium Stoloniferum; G.P.O.: Washington, DC, USA, 1913; Volume 270. [Google Scholar]
- Birkinshaw, J.H.; Oxford, A.E.; Raistrick, H. Studies in the Biochemistry of Micro-Organisms. Biochem. J. 1936, 30, 394–411. [Google Scholar] [CrossRef] [Green Version]
- Frisvad, J.C. A Critical Review of Producers of Small Lactone Mycotoxins: Patulin, Penicillic Acid and Moniliformin. World Mycotoxin, J. 2018, 11, 73–100. [Google Scholar] [CrossRef]
- Raphael, R.A. Compounds Related to Penicillic Acid; Synthesis of Dihydropenicillic Acid. J. Chem. Soc. 1947, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Shaw, E. A Synthesis of Protoanemonin. The Tautomerism of Acetylacrylic Acid and of Penicillic Acid. J. Am. Chem. Soc. 1946, 68, 2510–2513. [Google Scholar] [CrossRef]
- Shan, R.; Stadler, M.; Anke, H.; Sterner, O. The Reactivity of the Fungal Toxin Papyracillic Acid. Tetrahedron 1997, 53, 6209–6214. [Google Scholar] [CrossRef]
- Birch, A.J.; Blance, G.E.; Smith, H. Studies in Relation to Biosynthesis. Part XVIII. Penicillic Acid. J. Chem. Soc. 1958, 4582–4583. [Google Scholar] [CrossRef]
- Bentley, R.; Keil, J.G. Tetronic Acid Biosynthesis in Molds. II. Formation of Penicillic Acid in Penicillium Cyclopium. J. Biol. Chem. 1962, 237, 867–873. [Google Scholar] [CrossRef]
- Olivigni, F.J.; Bullerman, L.B. A Microbiological Assay for Penicillic Acid. J. Food. Prot. 1978, 41, 432–434. [Google Scholar] [CrossRef]
- Geiger, W.B.; Conn, J.E. The Mechanism of the Antibiotic Action of Clavacin and Penicillic Acid. J. Am. Chem. Soc. 1945, 67, 112–116. [Google Scholar] [CrossRef]
- EUCAST: Clinical Breakpoints and Dosing of Antibiotics. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 23 December 2021).
- Dalton, J.P.; Uy, B.; Okuda, K.S.; Hall, C.J.; Denny, W.A.; Crosier, P.S.; Swift, S.; Wiles, S. Screening of Anti-Mycobacterial Compounds in a Naturally Infected Zebrafish Larvae Model. J. Antimicrob. Chemother. 2017, 72, 421–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, J.P.; Uy, B.; Phummarin, N.; Copp, B.R.; Denny, W.A.; Swift, S.; Wiles, S. Effect of Common and Experimental Anti-Tuberculosis Treatments on Mycobacterium tuberculosis Growing as Biofilms. Peer J. 2016, 4, e2717. [Google Scholar] [CrossRef] [Green Version]
- Grey, A.B.J.; Cadelis, M.M.; Diao, Y.; Park, D.; Lumley, T.; Weir, B.S.; Copp, B.R.; Wiles, S. Screening of Fungi for Antimycobacterial Activity Using a Medium-Throughput Bioluminescence-Based Assay. Front. Microbiol. 2021, 12, 739995. [Google Scholar] [CrossRef]
- Jouda, J.B.; Mawabo, I.K.; Notedji, A.; Mbazoa, C.D.; Nkenfou, J.; Wandji, J.; Nkenfou, C.N. Anti-mycobacterial activity of polyketides from Penicillium sp. endophyte isolated from Garcinia nobilis against Mycobacteriumsmegmatis. Int. J. Mycobacteriol. 2016, 5, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.-Y.; Park, M.S.; Lim, Y.W. The Influence of Microfungi on the Mycelial Growth of Ectomycorrhizal Fungus Tricholoma Matsutake. Microorganisms 2019, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- Blaskovich, M.A.T.; Zuegg, J.; Elliott, A.G.; Cooper, M.A. Helping Chemists Discover New Antibiotics. ACS Infect. Dis. 2015, 1, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Andreu, N.; Zelmer, A.; Fletcher, T.; Elkington, P.T.; Ward, T.H.; Ripoll, J.; Parish, T.; Bancroft, G.J.; Schaible, U.; Robertson, B.D.; et al. Optimisation of Bioluminescent Reporters for Use with Mycobacteria. PLoS ONE 2010, 5, e10777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiles, S.; Grey, A. Bioluminescence-Based Minimum Inhibitory Concentration (MIC) Testing of Pure Compounds Isolated from Fungi against Mycobacterium abscessus. 2021. Available online: Dx.doi.org/10.17504/protocols.io.bumcnu2w (accessed on 30 December 2021).
- Wiles, S.; Grey, A. Bioluminescence-Based Minimum Inhibitory Concentration (MIC) Testing of Pure Compounds Isolated from Fungi against Mycobacterium marinum. 2021. Available online: Dx.doi.org/10.17504/protocols.io.3x7gprn (accessed on 30 December 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1a | 1b | ||
|---|---|---|---|---|
| δH | δC | δH | δC | |
| 1 | 174.6 | 173.5 | ||
| 2 | 5.19 | 97.2 | 5.20 | 89.2 |
| 3 | 171.2 | 182.1 | ||
| 4 | 217.1 | 107.2 | ||
| 5 | Not observed | 39.3 | 2.12 | 34.6 |
| 6 | 1.07 | 17.5 | 1.04 | 16.1 |
| 6′ | 1.07 | 17.5 | 0.88 | 16.4 |
| 7 | 3.72 | 57.8 | 3.93 | 59.9 |
| Position | Δc | Relative Integral (Labelled/Unlabelled) |
|---|---|---|
| C-1 | 181.2 | 12.3 |
| C-2 | 90.1 | 1.0 |
| C-3 | 173.2 | 10.1 |
| C-4 | Not observed | |
| C-5 | 141.7 | 8.5 |
| C-6 | 116.7 | 1.1 |
| C-7 | 17.5 | 1.0 |
| C-8 | 60.4 | 0.9 |
| Compound | Percentage Inhibition at 32 µg/mL | ||||||
|---|---|---|---|---|---|---|---|
| S. aa | E. cb | K. pc | P. ad | A. be | C. af | C. ng | |
| 1 | 53.07 **** | 26.11 *** | 18.06 * | −1.15 | 6.12 | 6.50 | 2.15 |
| 1b | −1.05 | 0.34 | 4.32 | 2.75 | −26.55 | 2.49 | 18.33 ** |
| 1c | 17.41 * | 5.15 | 9.34 | −10.06 | −11.88 | 2.56 | −7.04 |
| 2 | 14.22 | 11.16 | 13.49 | 4.70 | −4.31 | −1.86 | 1.11 |
| 3 | −7.23 | 13.60 | −7.65 | −4.08 | −24.24 | −2.20 | 20.72 ** |
| 4 | 71.52 **** | 9.29 | 15.62 * | 0.52 | 12.55 | 8.84 | 6.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cadelis, M.M.; Nipper, N.S.L.; Grey, A.; Geese, S.; van de Pas, S.J.; Weir, B.S.; Copp, B.R.; Wiles, S. Antimicrobial Polyketide Metabolites from Penicillium bissettii and P. glabrum. Molecules 2022, 27, 240. https://doi.org/10.3390/molecules27010240
Cadelis MM, Nipper NSL, Grey A, Geese S, van de Pas SJ, Weir BS, Copp BR, Wiles S. Antimicrobial Polyketide Metabolites from Penicillium bissettii and P. glabrum. Molecules. 2022; 27(1):240. https://doi.org/10.3390/molecules27010240
Chicago/Turabian StyleCadelis, Melissa M., Natasha S. L. Nipper, Alex Grey, Soeren Geese, Shara J. van de Pas, Bevan S. Weir, Brent R. Copp, and Siouxsie Wiles. 2022. "Antimicrobial Polyketide Metabolites from Penicillium bissettii and P. glabrum" Molecules 27, no. 1: 240. https://doi.org/10.3390/molecules27010240
APA StyleCadelis, M. M., Nipper, N. S. L., Grey, A., Geese, S., van de Pas, S. J., Weir, B. S., Copp, B. R., & Wiles, S. (2022). Antimicrobial Polyketide Metabolites from Penicillium bissettii and P. glabrum. Molecules, 27(1), 240. https://doi.org/10.3390/molecules27010240