
Chemical Composition Assessment of Structural Parts (Seeds, Peel, Pulp) of Physalis alkekengi L. Fruits

 , , ,
, , ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Basic Evaluation of Fresh Berries and Their Structural Parts
2.3. Determination of Fatty Acids in Seed and Peel Oils
2.4. Determination of Tocopherols in Seed and Peel Oils
2.5. Determination of Amino Acids in Seed Cakes
2.6. Determination of Mineral Elements in Seed Cakes
2.7. Determination of Volatiles in Fruit Concretes
2.8. Statistics
3. Results and Discussion
3.1. Basic Evaluation of Fruit Structural Parts
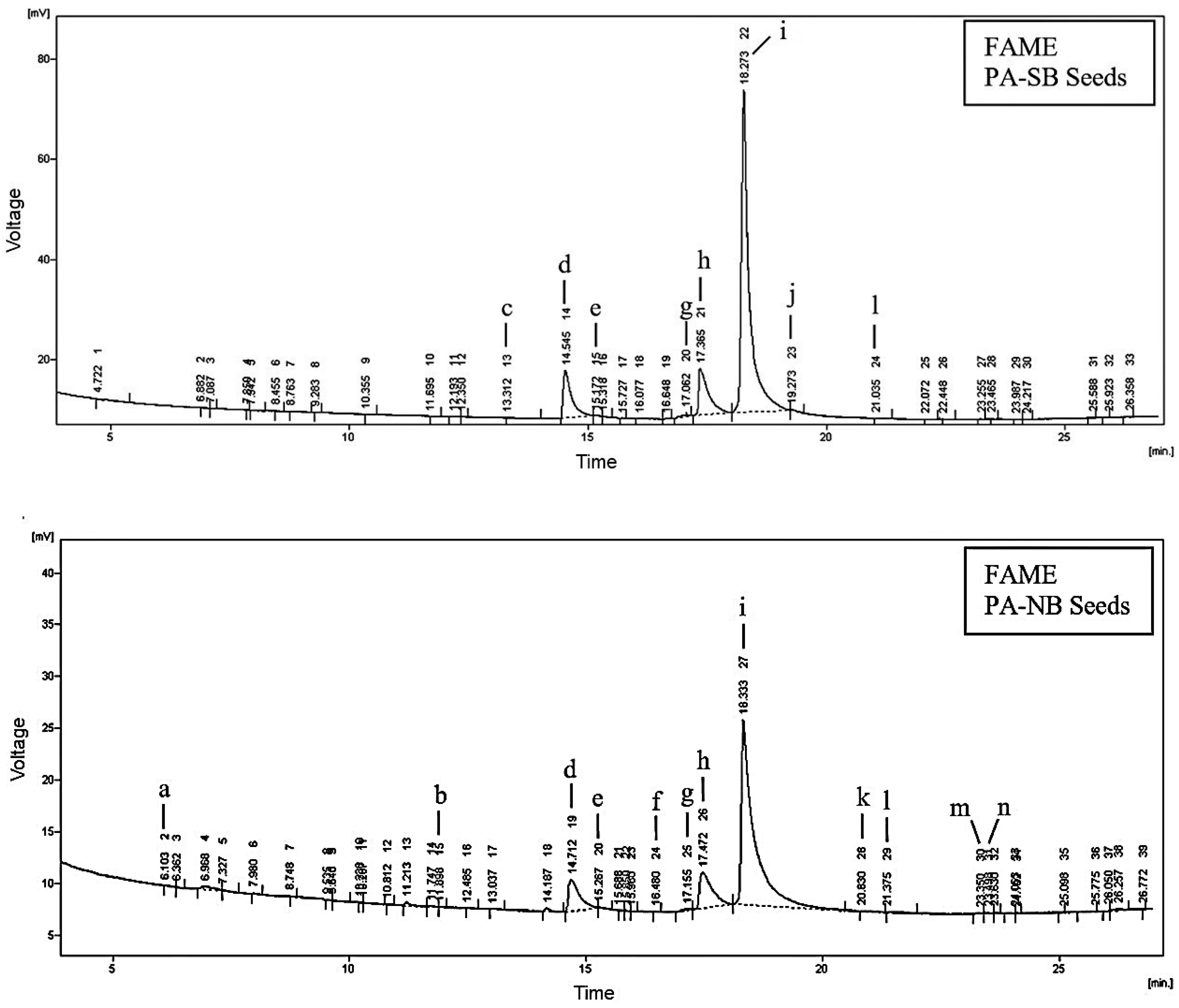
3.2. Determination of Fatty Acids in Seed and Peel Oils
3.3. Determination of Tocopherols in Seed and Peel Oils
3.4. Determination of Amino Acids in Seed Cakes
3.5. Determination of Mineral Elements in Seed Cakes
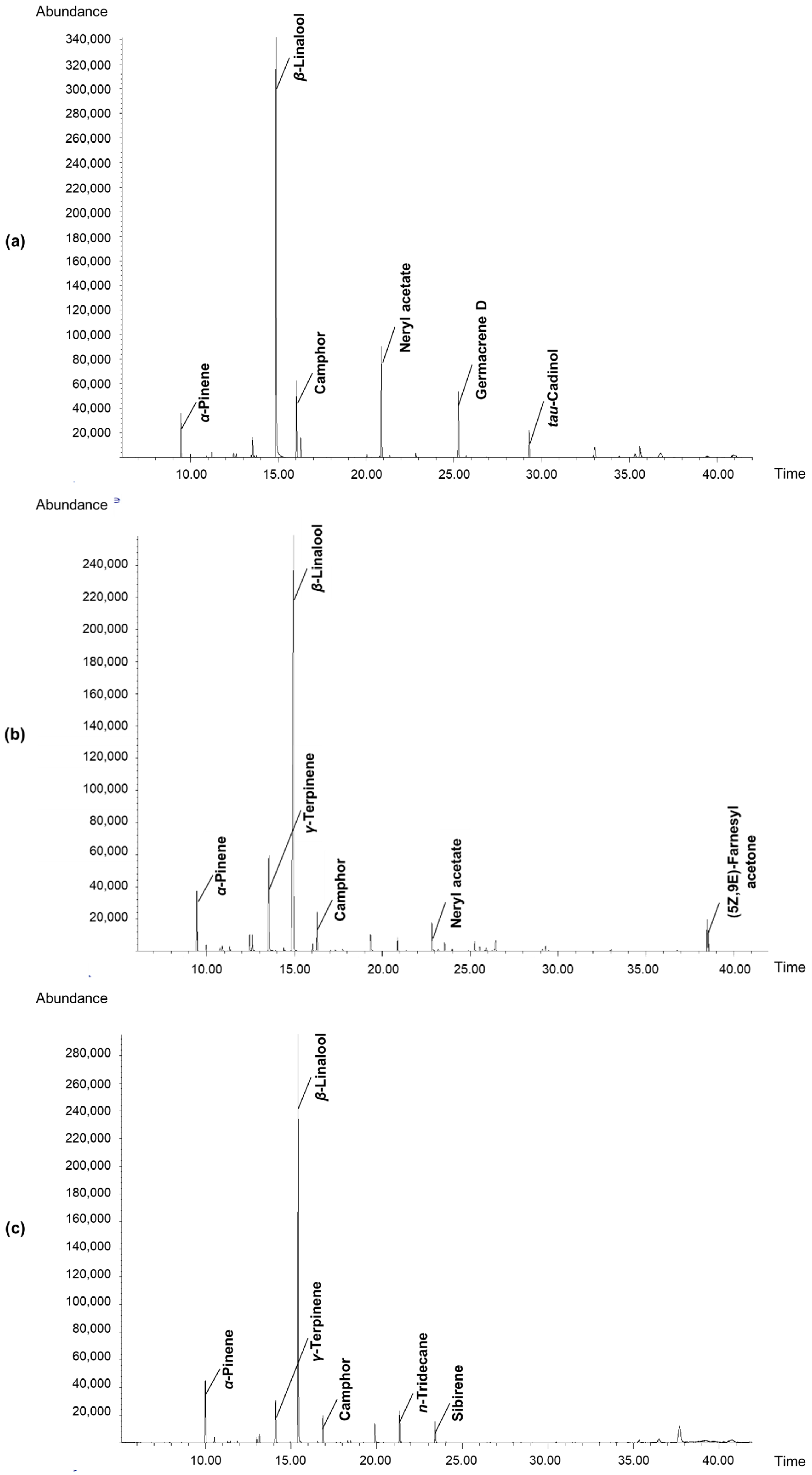
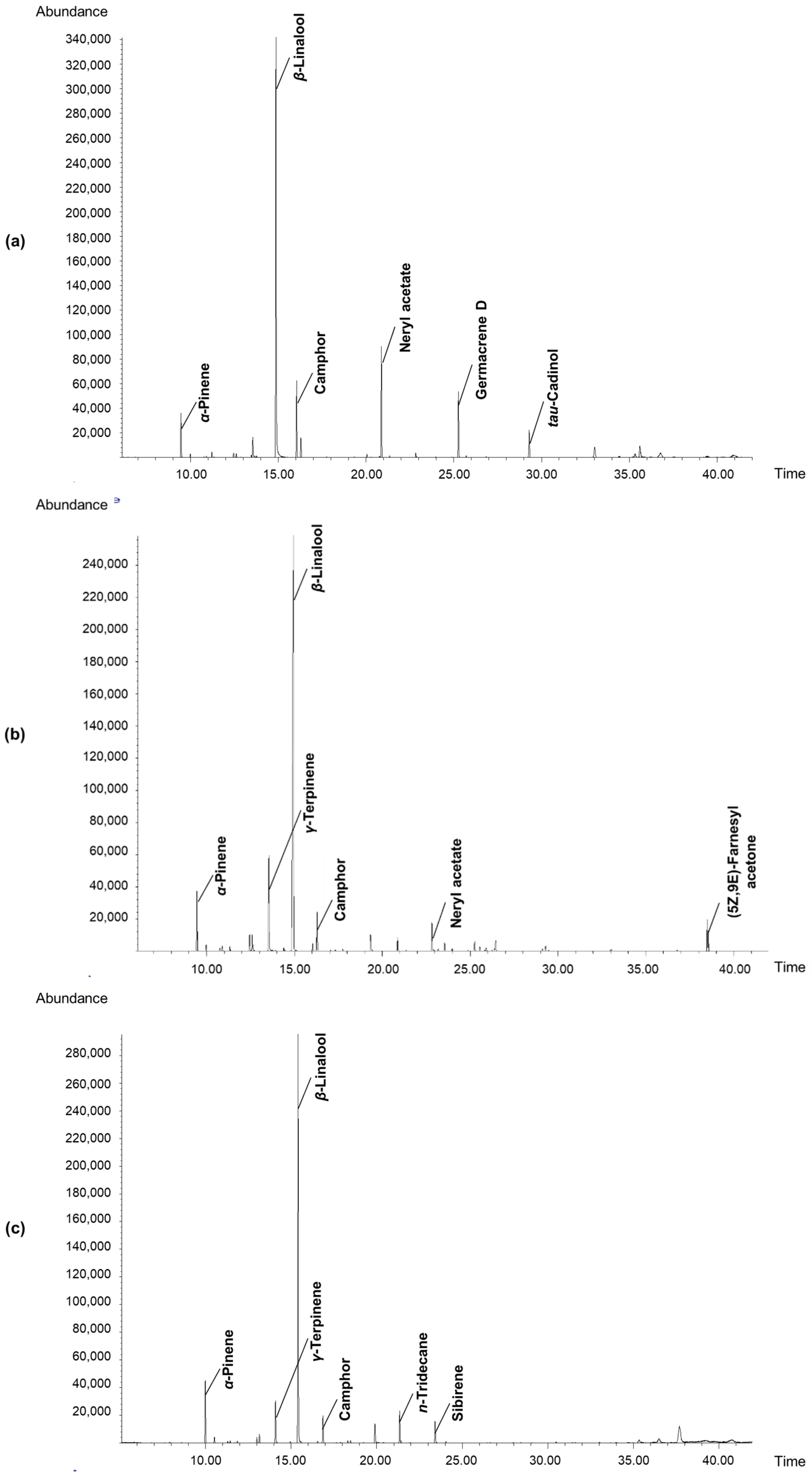
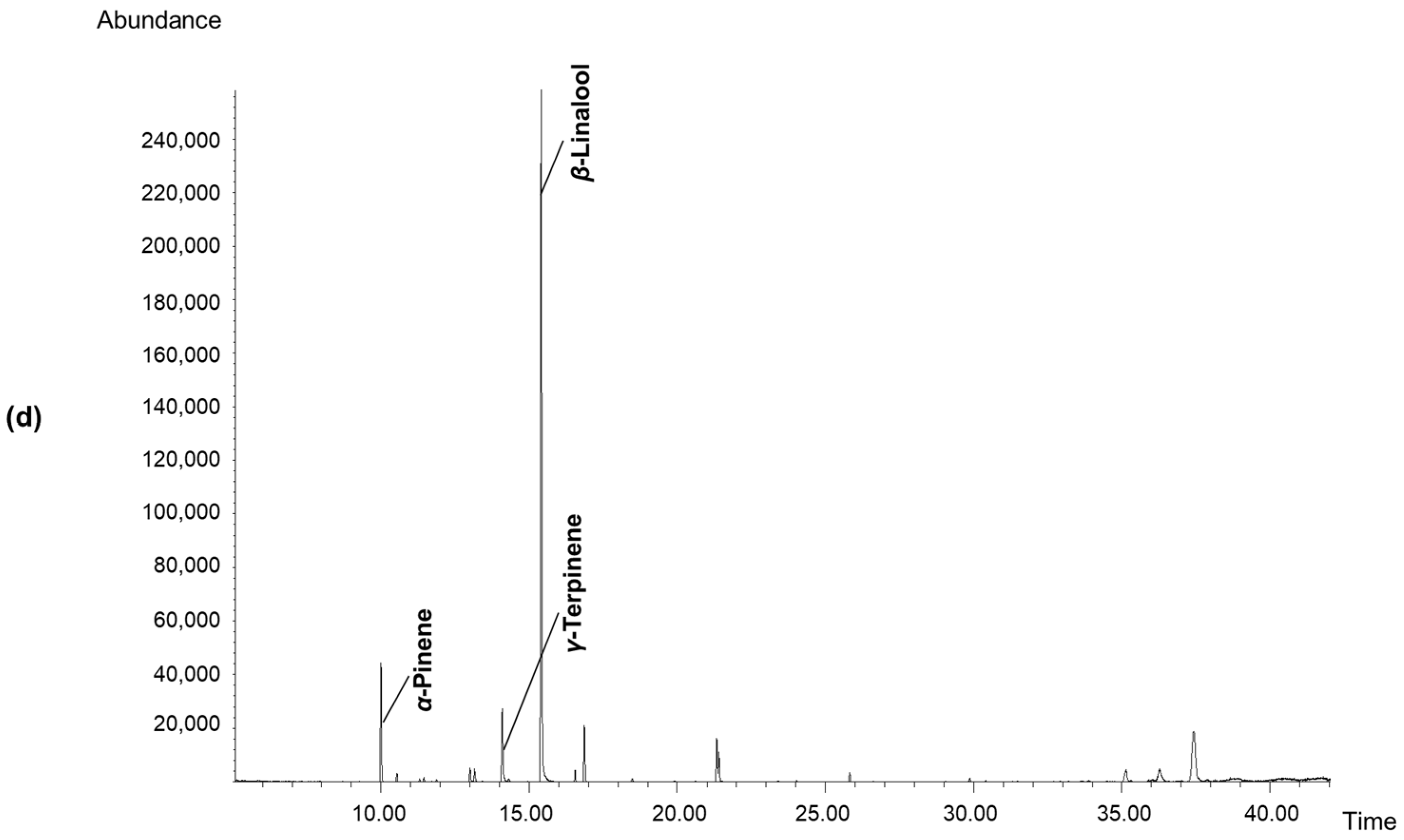
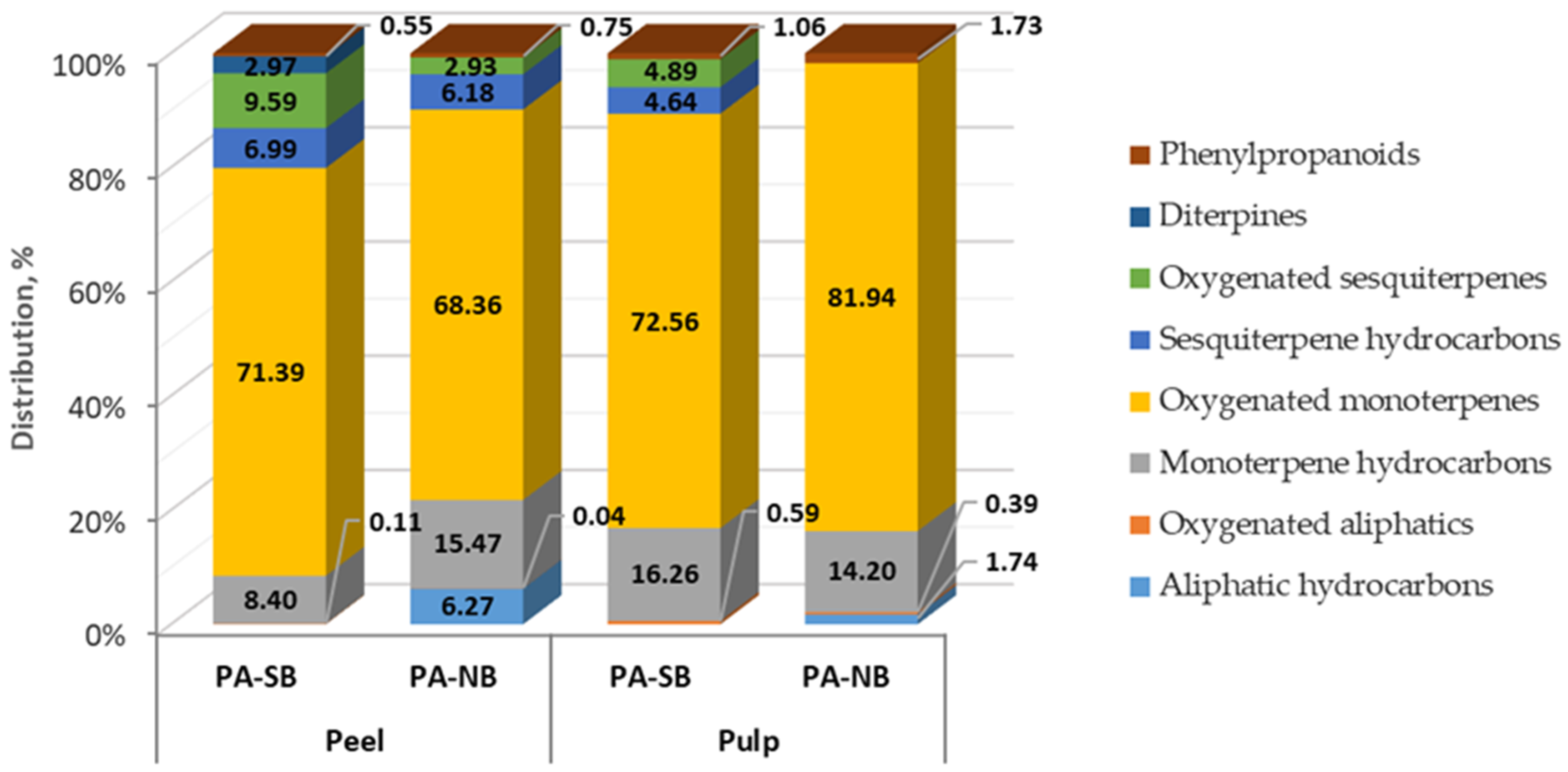
3.6. Determination of Volatile Components in Fruit Concretes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Namjoyan, F.; Jahangiri, A.; Azemi, M.E.; Arkian, E.; Mousavi, H. Inhibitory effects of Physalis alkekengi L., Alcea rosea L., Bunium persicum B. Fedtsch. and Marrubium vulgare L. on mushroom tyrosinase. Jundishapur J. Nat. Pharm. Prod. 2015, 10, e23356. [Google Scholar] [CrossRef]
- Li, A.; Chen, B.; Li, G.; Zhou, M.; Li, Y.; Ren, D.; Lou, H.; Wang, X.; Shen, T. Physalis alkekengi L. var. Franchetii (Mast.) Makino: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2018, 210, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Petkov, V. Contemporary Phytotherapy; Medicina i Fizkultura: Sofia, Bulgaria, 1982. (In Bulgarian) [Google Scholar]
- Delipavlov, D.; Cheshmedzhiev, I.; Popova, M.; Terzijski, D.; Kovatchev, I. Key to the Plants in Bulgaria; Agricultural University Academic Press: Plovdiv, Bulgaria, 2003. (In Bulgarian) [Google Scholar]
- Parliament of the Republic of Bulgaria. Medicinal Plants Act of the 38th Parliament of the Republic of Bulgaria of 23 March 2000. State Gaz. 2000, 29, 8. Available online: https://dv.parliament.bg/DVWeb/index.faces (accessed on 13 April 2022). (In Bulgarian).
- Evstatieva, L.; Hardalov, R.; Stoyanova, K. Medicinal plants in Bulgaria: Diversity, legislation, conservation and trade. Phytol. Balc. 2007, 13, 415–427. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/13_3/13_3_19_Evstatieva_&_al.pdf (accessed on 13 April 2022).
- Ivanov, I.; Landzhev, I.; Neshev, G. Herbs in Bulgaria and Their Use; Zemizdat: Sofia, Bulgaria, 1977. (In Bulgarian) [Google Scholar]
- Pintea, A.; Varga, A.; Stepnowski, P.; Socaciu, C.; Culea, M.; Diehl, H. Chromatographic analysis of carotenol fatty acid esters in Physalis alkekengi and Hippophae rhamnoides. Phytochem. Anal. 2005, 16, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Peng, W.-H.; Tsai, K.-D.; Hsu, S.-L. Luteolin suppresses inflammation-associated gene expression by blocking NF-κB and AP-1 activation pathway in mouse alveolar macrophages. Life Sci. 2007, 81, 1602–1614. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Sun, Y.; Cao, F.; Yang, B.; Kuang, H. Natural products from Physalis alkekengi L. var. franchetii (Mast.) Makino: A review on their structural analysis, quality control, pharmacology, and pharmacokinetics. Molecules 2022, 27, 695. [Google Scholar] [CrossRef]
- Tong, H.; Liang, Z.; Wang, G. Structural characterization and hypoglycemic activity of a polysaccharide isolated from the fruit of Physalis alkekengi L. Carbohydr. Polym. 2008, 71, 316–323. [Google Scholar] [CrossRef]
- Zarei, A.; Ashtiyani, S.; Mohamadi, A.; Gabari, A. The effects of Physalis alkekengi extract on lipids concentrations in rats. J. Arak Uni. Med. Sci. 2011, 14, 36–42. [Google Scholar]
- Chen, L.-X.; Xia, G.-Y.; Liu, Q.-Y.; Xie, Y.-Y.; Qiu, F. Chemical constituents from the calyces of Physalis alkekengi var. franchetii. Biochem. Syst. Ecol. 2014, 54, 31–35. [Google Scholar] [CrossRef]
- Shu, Z.; Xing, N.; Wang, Q.; Li, X.; Xu, B.; Li, Z.; Kuang, H. Antibacterial and anti-inflammatory activities of Physalis alkekengi var. franchetii and its main constituents. Evid. Based Complementary Altern. Med. 2016, 4359394. [Google Scholar] [CrossRef]
- Zhang, C.; Khan, W.; Bakht, J.; Nair, M. New antiinflammatory sucrose esters in the natural sticky coating of tomatillo (Physalis philadelphica), an important culinary fruit. Food Chem. 2016, 196, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bian, J.; Li, D.; Liu, C.; Xu, S.; Zhang, G.; Zhang, L.; Gao, P. Structural features, antioxidant and acetylcholinesterase inhibitory activities of polysaccharides from stem of Physalis alkekengi L. Ind. Crops Prod. 2019, 129, 654–661. [Google Scholar] [CrossRef]
- Vicas, L.G.; Jurca, T.; Baldea, I.; Filip, G.A.; Olteanu, D.; Clichici, S.V.; Pallag, A.; Marian, E.; Micle, O.; Crivii, C.B.; et al. Physalis alkekengi L. extract reduces the oxidative stress, inflammation and apoptosis in endothelial vascular cells exposed to hyperglycemia. Molecules 2020, 25, 3747. [Google Scholar] [CrossRef]
- Sharma, N.; Bano, A.; Dhaliwal, H.; Sharma, V. Perspectives and possibilities of Indian species of genus Physalis (L.)—A comprehensive review. Eur. J. Pharm. Med. Res. 2015, 2, 326–353. [Google Scholar]
- Bahmani, M.; Rafieian-Kopaei, M.; Naghdi, N.; Nejad, A.; Afsordeh, O. Physalis alkekengi: A review of its therapeutic effects. J. Chem. Pharm. Sci. 2016, 9, 1472–1475. Available online: http://eprints.umsha.ac.ir/id/eprint/2360 (accessed on 13 April 2022).
- Lu, Z.; Li, W.; Wang, P.J.; Liu, X.Y. Extraction of the oil from ground cherry seeds by Soxhlet method with different solvents and composition analysis. Chem. Adhesion 2011, 1, 18. [Google Scholar]
- Hu, X.-F.; Zhang, Q.; Zhang, P.-P.; Sun, L.-J.; Liang, J.-C.; Morris-Natschke, S.; Chen, Y.; Lee, K.-H. Evaluation of in vitro/in vivo anti-diabetic effects and identification, of compounds from Physalis alkekengi. Fitoterapia 2018, 127, 129–137. [Google Scholar] [CrossRef]
- Wen, X.; Erşan, S.; Li, M.; Wang, K.; Steingass, C.; Schweiggert, R.; Ni, Y.; Carle, R. Physicochemical characteristics and phytochemical profiles of yellow and red physalis (Physalis alkekengi L. and P. pubescens L.) fruits cultivated in China. Food Res. Int. 2019, 120, 389–398. [Google Scholar] [CrossRef]
- Liu, X.-G.; Jiang, F.-Y.; Gao, P.-Y.; Jin, M.; Yang, D.; Nian, Z.-F.; Zhang, Z.-X. Optimization of extraction conditions for flavonoids of Physalis alkekengi var. franchetii stems by response surface methodology and inhibition of acetylcholinesterase activity. J. Mex. Chem. Soc. 2015, 59, 59–66. [Google Scholar] [CrossRef]
- Cosmetic Ingredient Database (CosIng) of the European Commission. Available online: https://ec.europa.eu/growth/sectors/cosmetics_en (accessed on 18 March 2022).
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 20th ed.; AOAC International: Geithersburg, MD, USA, 2016. [Google Scholar]
- Brendel, O.; Iannetta, P.; Stewart, D. A rapid and simple method to isolate pure alpha-cellulose. Phytochem. Anal. 2000, 11, 7–10. [Google Scholar] [CrossRef]
- Standard ISO 659:2014; Oilseeds. Determination of Oil Content (Reference Method). International Organization for Standardization: Geneva, Switzerland, 2014.
- Standard ISO 12966-1:2014; Animal and Vegetable Fats and Oils. Gas Chromatography of Fatty Acid Methyl Esters—Part 1: Guidelines on Modern Gas Chromatography of Fatty Acid Methyl Esters. International Organization for Standardization: Geneva, Switzerland, 2014.
- Standard ISO 12966-2:2017; Animal and Vegetable Fats and Oils. Gas chromatography of Fatty Acid Methyl Esters—Part 2: Preparation of Methyl Esters of Fatty Acids. International Organization for Standardization: Geneva, Switzerland, 2017.
- Standard ISO 9936:2016; Animal and Vegetable Fats and Oils. Determination of Tocopherol and Tocotrienol Contents by High-performance Liquid Chromatography. International Organization for Standardization: Geneva, Switzerland, 2016.
- Standard ISO 14084:2003; Foodstuffs—Determination of Trace Elements—Determination of Lead, Cadmium, Zinc, Copper and Iron by Atomic Absorption Spectrometry (AAS) after Microwave Digestion. International Organization for Standardization: Geneva, Switzerland, 2003.
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- Shen, V.K.; Siderius, D.W.; Krekelberg, W.P.; Hatch, H.W. (Eds.) NIST Standard Reference Simulation Website, NIST Standard Reference Database 173; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2017. [Google Scholar] [CrossRef]
- Ramadan, M.F. Bioactive phytochemicals, nutritional value, and functional properties of Cape gooseberry (Physalis peruviana): An overview. Food Res. Int. 2011, 44, 1830–1836. [Google Scholar] [CrossRef]
- Popov, A.; Ilinov, P. Chemistry of Lipids; Nauka i Iskustvo: Sofia, Bulgaria, 1986. (In Bulgarian) [Google Scholar]
- Heuzé, V.; Tran, G. Grape Seeds and Grape Seed Oil Meal. Feedipedia, a Programme by INRA, CIRAD, AFZ and FAO. 2017. Available online: https://feedipedia.org/node/692 (accessed on 18 March 2022).
- Asilbekova, D.; Ulchenko, N.; Glushenkova, A. Lipids from Physalis alkekengi. Chem. Nat. Compd 2016, 52, 96–97. [Google Scholar] [CrossRef]
- Petkova, N.; Popova, V.; Ivanova, T.; Mazova, N.; Panayotov, N.; Stoyanova, A. Nutritional composition of different Cape gooseberry genotypes (Physalis peruviana L.)—A comparative study. Food Res. 2021, 5, 191–202. [Google Scholar] [CrossRef]
- Szymańska-Chargot, M.; Chylińska, M.; Gdula, K.; Kozioł, A.; Zdunek, A. Isolation and characterization of cellulose from different fruit and vegetable pomaces. Polymers 2017, 9, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministry of Health of the Republic of Bulgaria. Ordinance No 1 of 22 January 2018 on the Physiological Norms of Nutrition for the Population. State Gaz. 2018, 11, 2. Available online: https://dv.parliament.bg/DVWeb/broeveList.faces (accessed on 13 April 2022). (In Bulgarian).
- Baser, K.H.C.; Buchbauer, G. Handbook of Essential Oils: Science, Technology, and Applications; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruit Part | PA-SB (% fw) | PA-NB (% fw) |
|---|---|---|
| Peel | 4.99 ± 0.05 a | 5.43 ± 0.05 a |
| Seeds | 23.58 ± 0.18 b | 26.15 ± 0.23 b |
| Pulp | 71.43 ± 0.73 c | 68.42 ± 0.64 c |
| Component | PA-SB | PA-NB | ||
|---|---|---|---|---|
| Seeds (% dw) | Peel/Pulp (% dw) | Seeds (% dw) | Peel/Pulp (% dw) | |
| Cellulose | 8.06 ± 0.07 d | 1.44 ± 0.01 a | 6.12 ± 0.05 c | 2.15 ± 0.01 b |
| Protein | 19.14 ± 0.14 d | 2.51 ± 0.01 b | 16.22 ± 0.12 c | 1.94 ± 0.01 a |
| Oil | 14.13 ± 0.12 c | 1.27 ± 0.01 a | 17.57 ± 0.15 d | 1.81 ± 0.01 b |
| Fatty Acid | PA-SB | PA-NB | |||
|---|---|---|---|---|---|
| Seeds (% dw) | Peel (% dw) | Seeds (% dw) | Peel (% dw) | ||
| C10:0 | Capric | nd 1 | nd | 0.10 ± 0.0 a | 0.09 ± 0.0 a |
| C11:0 | Undecylic | nd | 0.33 ± 0.0 b | nd | 0.17 ± 0.0 a |
| C12:0 | Lauric | nd | 1.08 ± 0.0 a | nd | 1.11 ± 0.01 a |
| C13:0 | Tridecylic | nd | 0.09 ± 0.0 a | nd | 0.42 ± 0.0 b |
| C14:0 | Myristic | nd | 3.89 ± 0.01 b | 0.11 ± 0.0 a | 5.01 ± 0.02 c |
| C14:1 | Myristoleic | nd | nd | nd | 0.82 ± 0.0 a |
| C15:0 | Pentadecylic | 0.21 ± 0.0 a | nd | nd | 4.44 ± 0.01 b |
| C16:0 | Palmitic | 11.28 ± 0.10 b | 57.88 ± 0.42 d | 10.49 ± 0.10 a | 36.21 ± 0.29 c |
| C16:1 | Palmitoleic | 0.12 ± 0.0 a | 1.79 ± 0.01 b | 0.18 ± 0.0 a | 8.22 ± 0.02 c |
| C17:0 | Margaric | nd | 0.41 ± 0.0 a | nd | 0.57 ± 0.0 a |
| C17:1 | Heptadecenoic | nd | 0.69 ± 0.0 b | 0.11 ± 0.0 a | 0.49 ± 0.0 b |
| C18:0 | Stearic | 0.51 ± 0.0 a | 4.32 ± 0.02 b | 0.42 ± 0.0 a | 8.37 ± 0.02 c |
| C18:1 | Oleic | 13.88 ± 0.11 a | 24.02 ± 0.31 b | 13.39 ± 0.12 a | 30.08 ± 0.21 c |
| C18:2 (n-6) | Linoleic | 73.67 ± 0.72 b | 1.00 ± 0.01 a | 74.43 ± 0.71 b | 0.83 ±0.0 a |
| C18:3 (n-3) | Linolenic | 0.21 ± 0.0 a | 2.38 ± 0.01 c | nd | 2.09 ± 0.02 b |
| C20:2 (n-6) | Eicosadienoic | nd | 2.12 ± 0.01 c | 0.28 ± 0.0 a | 1.08 ± 0.01 b |
| C20:3 (n-6) | Eicosatrienoic | 0.12 ± 0.0 a | nd | 0.29 ± 0.0 b | nd |
| C20:4 (n-6) | Eicosatetraenoic | nd | nd | 0.10 ± 0.0 | nd |
| C20:5 (n-3) | Eicosapentaenoic | nd | nd | 0.10 ± 0.0 | nd |
| Saturated fatty acids | 12.00 | 68.00 | 11.12 | 56.40 | |
| Unsaturated fatty acids, of which | 88.00 | 32.00 | 88.88 | 43.60 | |
| Monounsaturated fatty acids | 14.00 | 26.50 | 13.68 | 39.60 | |
| Polyunsaturated fatty acids | 74.00 | 5.50 | 75.20 | 4.00 | |
| Tocopherols | PA-SB | PS-NB | ||
|---|---|---|---|---|
| Seeds | Peel | Seeds | Peel | |
| α-Tocopherol (% of the total tocopherols) | 1.01 ± 0.01 a | nd 1 | nd | 100 ± 0.01 b |
| β-Tocopherol (% of the total tocopherols) | 70.63 ± 0.68 b | 30.32 ± 0.20 a | 76.61 ± 0.71 c | nd |
| γ-Tocopherol (% of the total tocopherols) | 28.44 ± 0.22 b | 69.74 ± 0.31 c | 23.42 ± 0.21 a | nd |
| Total tocopherols (mg/kg dw) | 5378 ± 51.00 d | 340.00 ± 17.00 b | 2009 ± 20.00 c | 216.00 ± 11.00 a |
| Amino Acid | PA-SB (mg/g dw) | PA-NB (mg/g dw) |
|---|---|---|
| Aspartic acid | 12.23 ± 0.11 a | 12.16 ± 0.11 a |
| Serine | 8.41 ± 0.07 b | 6.08 ± 0.06 a |
| Glutamic acid | 7.12 ± 0.07 a | 11.97 ± 0.10 b |
| Glycine | 10.94 ± 0.08 a | 14.83 ± 0.11 b |
| Histidine | 7.18 ± 0.07 b | 5.90 ± 0.05 a |
| Arginine | 15.75 ± 0.11 b | 13.17 ± 0.11 a |
| Threonine 1 | 4.17 ± 0.03 b | 2.89 ± 0.03 a |
| Alanine | 9.31 ± 0.08 a | 14.34 ± 0.12 b |
| Proline | 2.54 ± 0.01 a | 4.27 ± 0.02 b |
| Cysteine | 0.24 ± 0.0 b | 0.14 ± 0.0 a |
| Tyrosine | 3.20 ± 0.02 a | 6.19 ± 0.04 b |
| Valine 1 | 4.53 ± 0.02 b | 3.56 ± 0.02 a |
| Methionine 1 | 0.77 ± 0.0 a | 0.70 ± 0.0 a |
| Lysine 1 | 2.94 ± 0.01 a | 3.49 ± 0.02 b |
| Isoleucine 1 | 3.37 ± 0.01 a | 4.41 ± 0.01 b |
| Leucine 1 | 0.56 ± 0.0 a | 0.86 ± 0.0 b |
| Phenylalanine 1 | 3.19 ± 0.02 a | 4.63 ± 0.02 b |
| Mineral Element | PA-SB (mg/kg dw) | PA-NB (mg/kg dw) |
|---|---|---|
| Potassium (K) | 4122.28 ± 19.43 a | 4668.32 ± 21.23 a |
| Sodium (Na) | 182.33 ± 1.33 b | 80.79 ± 0.33 a |
| Calcium (Ca) | 529.15 ± 1.87 a | 1586.23 ± 4.78 b |
| Magnesium (Mg) | 2418.97 ± 11.24 a | 2318.26 ± 11.09 a |
| Iron (Fe) | 61.71 ± 0.21 b | 50.38 ± 0.19 a |
| Manganese (Mn) | 25.40 ± 0.09 a | 23.42 ± 0.08 a |
| Copper (Cu) | 9.98 ± 0.03 a | 10.30 ± 0.04 a |
| Zinc (Zn) | 29.64 ± 0.09 b | 25.24 ± 0.08 a |
| Lead (Pb) | 2.67 ± 0.0 a | 2.93 ± 0.0 a |
| Cadmium (Cd) | nd 1 | nd |
| Chromium (Cr) | nd | 2.32 ± 0.0 |
| Index | PA-SB (% DW; w/w) | PS-NB (% DW; w/w) |
|---|---|---|
| Yield of pulp concrete | 0.02 ± 0.00 a | 0.03 ± 0.00 a |
| Yield of peel concrete | 0.72 ± 0.01 a | 2.16 ± 0.01 b |
| Volatiles | RT 1 | RI 2 | PA-SB (% or TIC 2) | PA-NB (% or TIC 3) | ||
|---|---|---|---|---|---|---|
| Peel | Pulp | Peel | Pulp | |||
| α-Pinene | 9.45 | 933 | 4.63 ± 0.03 a | 5.30 ± 0.04 b | 7.57 ± 0.06 d | 6.85 ± 0.05 c |
| Camphene | 9.98 | 945 | 0.31 ± 0.0 a | 0.53 ± 0.0 b | 0.69 ± 0.0 c | 0.28 ± 0.0 a |
| Sabinene | 10.76 | 969 | 0.05 ± 0.0 a | 0.22 ± 0.0 b | 0.08 ± 0.0 a | 0.15 ± 0.0 b |
| β-Pinene | 10.90 | 975 | 0.09 ± 0.0 a | 0.41 ± 0.0 c | 0.06 ± 0.0 a | 0.12 ± 0.0 b |
| Myrcene | 11.33 | 987 | 0.44 ± 0.0 b | 0.59 ± 0.0 c | 0.23 ± 0.0 a | 0.25 ± 0.0 a |
| p-Cymene | 12.45 | 1020 | 0.54 ± 0.0 a | 1.04 ± 0.09 c | 0.74 ± 0.0 b | 0.56 ± 0.0 a |
| Limonene | 12.60 | 1023 | 0.43 ± 0.0 a | 1.21 ± 0.03 d | 1.07 ± 0.09 c | 0.89 ± 0.0 b |
| γ-Terpinene | 13.54 | 1055 | 2.34 ± 0.01 a | 7.77 ± 0.06 c | 5.52 ± 0.05 b | 5.46 ± 0.05 b |
| Camphenilone | 14.40 | 1077 | 0.35 ± 0.0 b | 0.41 ± 0.0 b | 0.12 ± 0.0 a | 0.09 ± 0.0 a |
| β-Linalool | 14.92 | 1095 | 49.57 ± 0.47 a | 61.91 ± 0.60 c | 57.76 ± 0.50 b | 78.29 ± 0.70 d |
| Nonanal | 15.02 | 1101 | 0.11 ± 0.0 b | 0.58 ± 0.0 d | 0.04 ± 0.0 a | 0.38 ± 0.0 c |
| Camphor | 16.29 | 1140 | 7.53 ± 0.06 d | 3.26 ± 0.03 b | 5.53 ± 0.05 c | 2.31 ± 0.02 a |
| 1-Terpinen-4-ol | 17.31 | 1174 | 0.04 ± 0.0 a | 0.18 ± 0.0 b | nd 4 | nd |
| α-Terpineol | 17.75 | 1185 | 0.09 ± 0.0 a | 0.32 ± 0.0 b | nd | nd |
| Nerol | 19.33 | 1126 | 1.48 ± 0.01 b | 1.73 ± 0.01 c | 2.80 ± 0.02 d | 0.06 ± 0.0 a |
| n-Tridecane | 21.34 | 1300 | nd | nd | 6.18 ± 0.05 b | 1.72 ± 0.01 a |
| Isoamyl benzyl ether | 21.40 | 1310 | nd | nd | nd | 1.15 ± 0.01 |
| Neryl acetate | 22.83 | 1349 | 11.42 ± 0.10 c | 3.71 ± 0.03 b | 1.06 ± 0.01 a | nd |
| Sibirene | 23.55 | 1398 | nd | 0.72 ± 0.0 a | 5.85 ± 0.05 b | nd |
| β-Caryophyllene | 23.98 | 1419 | nd | 0.33 ± 0.0 | nd | nd |
| Germacrene D | 25.24 | 1483 | 6.85 ± 0.06 c | 0.88 ± 0.0 b | 0.23 ± 0.03 a | nd |
| β-Selinene | 25.40 | 1491 | 0.05 ± 0.0 a | 0.43 ± 0.0 b | nd | nd |
| α-Zingiberene | 25.61 | 1493 | nd | 0.27 ± 0.0 | nd | nd |
| Bicyclogermacrene | 25.91 | 1501 | nd | 0.55 ± 0.0 | nd | nd |
| δ-Cadinene | 26.55 | 1522 | nd | 1.07 ± 0.01 | nd | nd |
| α-Cadinene | 26.80 | 1536 | nd | 0.32 ± 0.05 | nd | nd |
| 1-epi-Cubenol | 29.11 | 1627 | nd | 0.41 ± 0.05 | nd | nd |
| tau-Cadinol | 29.30 | 1640 | 3.69 ± 0.03 b | 0.54 ± 0.0 a | nd | nd |
| (2E,6E)-Methyl farnesoate | 35.12 | 1785 | 1.11 ± 0.01 | nd | nd | nd |
| (2Z,6E)-Farnesyl acetate | 36.19 | 1820 | 2.58 ± 0.02 | nd | nd | nd |
| (5Z,9E)-Farnesyl acetone | 38.30 | 1889 | 2.09 ± 0.02 b | 3.87 ± 0.03 c | 2.88 ± 0.02 a | nd |
| Phytol | 40.74 | 1940 | 2.93 ± 0.02 | nd | nd | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popova, V.; Petkova, Z.; Mazova, N.; Ivanova, T.; Petkova, N.; Stoyanova, M.; Stoyanova, A.; Ercisli, S.; Okcu, Z.; Skrovankova, S.; et al. Chemical Composition Assessment of Structural Parts (Seeds, Peel, Pulp) of Physalis alkekengi L. Fruits. Molecules 2022, 27, 5787. https://doi.org/10.3390/molecules27185787
Popova V, Petkova Z, Mazova N, Ivanova T, Petkova N, Stoyanova M, Stoyanova A, Ercisli S, Okcu Z, Skrovankova S, et al. Chemical Composition Assessment of Structural Parts (Seeds, Peel, Pulp) of Physalis alkekengi L. Fruits. Molecules. 2022; 27(18):5787. https://doi.org/10.3390/molecules27185787
Chicago/Turabian StylePopova, Venelina, Zhana Petkova, Nadezhda Mazova, Tanya Ivanova, Nadezhda Petkova, Magdalena Stoyanova, Albena Stoyanova, Sezai Ercisli, Zuhal Okcu, Sona Skrovankova, and et al. 2022. "Chemical Composition Assessment of Structural Parts (Seeds, Peel, Pulp) of Physalis alkekengi L. Fruits" Molecules 27, no. 18: 5787. https://doi.org/10.3390/molecules27185787
APA StylePopova, V., Petkova, Z., Mazova, N., Ivanova, T., Petkova, N., Stoyanova, M., Stoyanova, A., Ercisli, S., Okcu, Z., Skrovankova, S., & Mlcek, J. (2022). Chemical Composition Assessment of Structural Parts (Seeds, Peel, Pulp) of Physalis alkekengi L. Fruits. Molecules, 27(18), 5787. https://doi.org/10.3390/molecules27185787