1. Introduction
Obesity is the excessive accumulation of body fat, mainly due to genetics, environmental factors, and the gut microbiota. Increasing evidence shows that long-term obesity may trigger specific metabolic disorders, like cardiovascular diseases, dyslipidemia, asthma, insulin resistance, type 2 diabetes, non-alcoholic fatty liver disease (NAFLD) and some types of cancer [
1,
2,
3,
4]. It has been reported that about 1.1 billion adults are overweight and 312 million are obese [
5]. Moreover, it has been reported that about 60% of dogs and cats are overweight or obese [
6,
7]. Therefore, its prevention and treatment in humans and companion animals is a serious societal concern [
8,
9].
In recent years, probiotics has been used to treat obesity [
10]. Specific bacteria are associated with obesity and related metabolism, so they can be regarded as therapeutic targets. As such, the intestinal mucin-degrading symbiont
Akkermansia muciniphila is generally considered to be important for maintaining gut health and glucose homoeostasis [
11]. However, effective probiotic therapies with anti-obesity applications for cats and dogs are not available [
12], perhaps because of the diverse gut microbiota of these species. Consequently, typical therapies to treat obese pets are changes in exercise and diet [
13].
Recent studies have found that the relative abundance of
A. muciniphila decreases in obese mice, whereas administration of fructooligosaccharides to obese mice increases it approximately 100-fold [
14]. Administration of live
A. muciniphila reverses high-fat diet-induced metabolic disorders, which might be associated with an increase in the intestinal levels of endocannabinoids [
15]. However, its anti-obesity effect on dogs has yet to be revealed, and the exact physiological function it plays during these processes has remained elusive.
To verify its anti-obesity function and provide a rationale for its sustainable use and commercial development in the treatment of dogs, we isolated A. muciniphila strains from mice and investigated anti-obesity in mice and dogs.
3. Materials and Methods
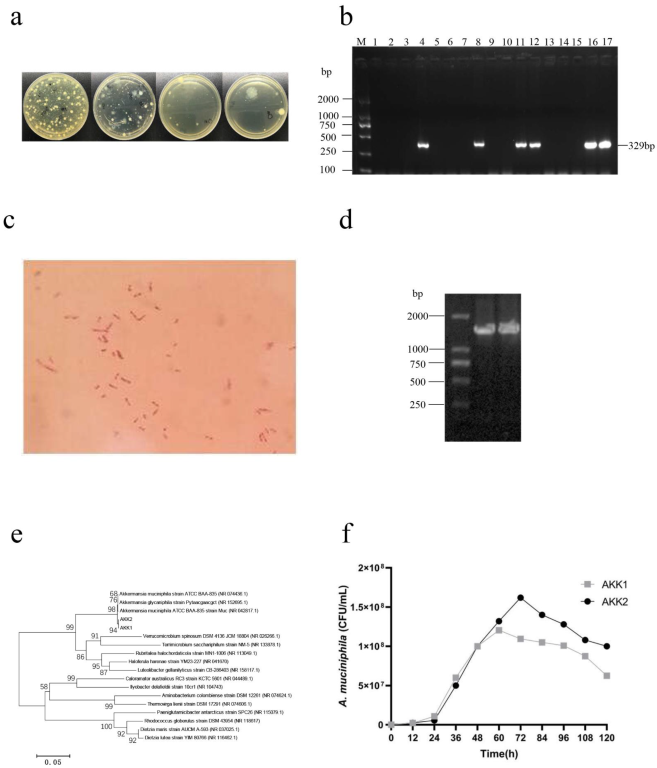
3.1. Isolation, Identification and Selection of A. muciniphila
The fresh fecal samples of interferon-inducible protein 204 (IFI204)-deficient male mice were collected and suspended in a sterile synthetic liquid medium (3.8% w/v Brain Heart Infusion (BHI) powder, 1.6% w/v soya peptone, 0.4% w/v threonine, 25 mM glucose, 25 mM N-acetylglucosamine, pH 7.2–7.4) and homogenized for 3 min. One hundred microliter aliquots were taken and serially diluted 10-fold with sterile physiological saline. Then, 0.1 mL of appropriate dilutions was spread onto BHI agar supplemented with 4 g/L mucin and 5 μg/mL vancomycin (Vancomycin agar), which were then incubated at 37 °C under anaerobic conditions (85% N2, 10% CO2 and 5% H2) for 5–7 d. Colonies with different morphologies were picked out and subcultured onto fresh vancomycin agar plates until the cultures were deemed to be pure. Isolated colonies were characterized preliminarily on the basis of morphology and Gram staining. Furthermore, each isolate was PCR-amplified using the A. muciniphila specific primers (forward, 5-GCATATCAATAAGCGGAGGAAAAG; reverse, 5-GGTCCGTGTTTCAAGACGG). Positive clones were further identified by amplification and sequencing of 16S rRNA genes with universal primers 27F and 1492R (GenBank: OM956292, OM956294).
A. muciniphila strains AKK1 and AKK2 were inoculated in a synthetic liquid medium and incubated for 72 h at 37 °C under anaerobic conditions. At the stationary phase, the bacteria suspension was transferred to 20 mL of fresh synthetic liquid medium at a ratio of 1% for the physiological studies presented here. The inoculated broth was incubated as described above, and 1 mL samples were taken at 12 h intervals for 120 h. The samples were serially diluted 10-fold in PBS and 100 µL from each dilution was plated on LB agar plates in triplicate followed by incubation at 37 °C for 72 h. The bacterial colonies were counted and expressed as colony-forming units (CFUs)/mL.
The two strains were inoculated in synthetic the liquid medium at pH 2.0, 3.0, 4.0, 5.0, and 7.0 under anaerobic condition (85% N
2, 10% CO
2 and 5% H
2) for 2 h [
28]. Then the suspensions were serially diluted, plated onto BHI agar medium, and incubated at 37 °C for 72 h to count CFUs. The bile salt tolerance test was performed as described previously, but the synthetic liquid medium contained 0.1, 0.2, 0.3 and 0.4% bile salt [
29].
On the other hand, two strains were inoculated in a synthetic liquid medium at 37 °C under anaerobic conditions for 48 h. The cultures were centrifuged at 4500× g for 20 min; the pellets were partly pasteurized at 70 °C for 30 min; and the bacteria were stored at −80 °C in 25% glycerol until needed.
3.2. Animal Experiments
All animal studies complied with the guidelines for the care and use of laboratory animals as described by the U.S. National Institutes of Health and were approved by the Animal Welfare and Research Ethics Committee at Jilin University (No. 20150601).
3.3. Mouse Trial
IFI204-deficient male mice were purchased from Nanjing Biomedical Research Institute of Nanjing University (Nanjing, China) and were subsequently backcrossed onto the C57BL/6J background at least eight generations. Six-week-old male C57BL/6J wild-type (WT) mice were purchased from Huafukang Biotechnology Co., Ltd. (Beijing, China) and housed in a 12 h light/dark cycle under controlled temperature and humidity. The normal diet (ND) contained 19% protein, 10% fat and 71% carbohydrate: total calories, 3.6 kcal/g. Liaoning Changsheng Biotechnology Co., Ltd., Benxi, China). The high-fat diet (HFD) contained 19.4% protein, 20.6% carbohydrate and 60% fat: total calories, 5.0 kcal/g (TP23300, Trophic Animal Feed High-Tech Co., Ltd., Nantong, China).
Twenty-four C57BL/6J mice were divided randomly into normal (ND, n = 8), high-fat diet groups (HFD, n = 8), and A. muciniphila AKK2-treated high-fat diet groups (HFD + AKK2, n = 8). The HFD + AKK2 group was challenged with 2 × 108 CFU of the pasteurized A. muciniphila AKK2 strain orally in 150 µL of sterile PBS, and the ND and HFD groups were administered an equivalent volume of sterile PBS orally. The treatments with the above-mentioned regimens were performed daily. The body weights and fasting blood glucose of the mice were measured weekly following a 12 h fast. Blood samples were collected and placed in tubes at 4 °C for 2 h. The serum samples were then separated by centrifugation for 15 min at 2000× g, and stored at −20 °C until analysis.
3.4. Dog Trial
Male beagles about 1.5 years old were purchased from Kangping Experimental Animal Research Institute (Shenyang, China). All dogs were housed in individual cages and fed twice a day. All diets were prepared by our laboratory, and the content of protein and fat was tested to ensure essential daily nutrients. The HFD consisted of 63.16% cornmeal, 31.58% lard, 5.255% soy flour and 0.005% vitamins (8.78% protein, 19.56% fat, 71.66% carbohydrate). The normal diet (ND) consisted of 66.67% cornmeal, 22.22% lard, 11.105% soy flour and 0.005% vitamins (8.06% protein, 14.64% fat, 77.30% carbohydrate).
A total of 24 adult beagles were randomly divided into ND, HFD and HFD + AKK2 groups of eight each. The HFD + AKK2 group was challenged with 2 × 109 CFU of pasteurized A. muciniphila AKK2 strain orally in 100 µL of sterile PBS, while the ND and HFD groups were orally administered an equivalent volume of sterile PBS. The body weight of the beagles was measured weekly following a 12 h fast. The intervention lasted for 9 weeks. At weeks 0, 2, 4, 6, 8 and 10, fresh fecal samples were collected from each dog for further microbiological analysis. Blood samples were collected from the forelimb veins prior to the commencement of experiments and at week 9. Serum was then separated from clotted blood by centrifugation for 15 min at 2000× g, and stored at −20 °C until needed for analysis.
3.5. Analyses of the Serum Biochemical Parameters
Serum lipids (total cholesterol, triglycerides, LDL cholesterol, HDL cholesterol) were measured by enzymatic photometric methods (Roche Diagnostics, Mannheim, Germany). The serum concentrations of alanine aminotransferase (ALT), aspartate aminotransferase (AST) and glutamine transpeptidase (GGT) were tested by commercial assay kits (Beckman Coulter, Brea, CA, USA).
3.6. Body Condition Scoring (BCS)
The visual method for the body condition score (BCS) was confirmed to be a semi-quantitative tool for assessing body fat composition in multiple animal health conditions. In this study, the body condition of all the beagles was determined using the nine-scale BCS system. The score was evaluated once a week for 9 weeks and performed according to the descriptions and illustrations provided by WSAVA [
30]. Each dog was assessed by at least two persons who learned the BCS evaluation technique from a veterinarian.
3.7. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
At week 9 of the experimental schedule, all beagles underwent an OGTT following an overnight (18 h) fast as described previously [
31]. The ITT was performed by commercial assay kits (Changjin Bioscience Co., Changsha, China).
3.8. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
The abundance of
A. muciniphila was analyzed by RT-qPCR on a Light Cycler 96 platform (Roche, Mannheim, Germany). The DNA of fecal bacteria was extracted by TIANamp Stool DNA Kit (Tiangen, Beijing, China) according to the manufacturer’s instructions. The primer sequences used are listed in
Table 1.
3.9. High-Throughput Sequencing of Intestinal Flora
The fecal samples of beagles were analyzed to determine the gut microbial composition. DNA was extracted and purified from each sample as previously described, and the V4–V5 region of the 16S rRNA gene was amplified using modified universal bacterial primers. Purified PCR products were sequenced using the HiSeq Illumina platform. The sequences were analyzed using the QIIME software [
32]. The UPARSE pipeline was used to cluster the sequences with 97% similarity into operational taxonomic units (OTUs). An operating taxon with domain values of 0.99 was considered to be a genus. A representative sequence was selected for each OUT and was compared using an RDP classifier (v 2.2) [
33].
3.10. Microcapsule Fabrication of A. muciniphila
The microcapsule of
A. muciniphila AKK2 was prepared by a modified chitosan linked alginate method after being resuspended in a sterile alginate solution containing 5% trehalose and 5% sucrose and transferred into sterile mixed colloidal solution (
v:
v = 1:1). The mixed colloidal solution consisted of 2% low methoxy pectin and 1.5% sodium alginate and was autoclaved at 121 °C for 20 min. The mixture was then squeezed into a 0.3 M calcium chloride solution using a 1 mL injector to crosslink with calcium and form droplets. The crosslinked droplets were generated after shaking at 200 rpm for 30 min on a magnetic stirrer and then filtered and washed with sterile water, as previous research reported [
34,
35,
36]. To improve formation efficiency, the microcapsules were then placed in a 10 mL microfiltered chitosan solution (0.4%
w/
v in 0.1 M acetic acid, adjusted to pH 6.0 with 1 M NaOH) to allow for coating the alginate droplet. Droplets were collected by filtration before washing and freeze-drying. The microcapsule formation efficiency was measured by the ratio of droplet bacteria to original bacteria.
3.11. Simulated Gastrointestinal Fluid Assay
Simulated gastric fluid consisted of 1% w/v pepsin and adjusted to pH 2.0 with 1 M HCl. Simulated intestinal fluid was made by mixing 5.3 mL monosodium phosphate (NaH2PO4) solution (0.2 mol/L) with 94.7 mL disodium phosphate (Na2HPO4) solution (0.2 mol/L) adjusted to pH 7.2 with 1 M HCl. The release of cells from the microcapsules was determined by placing 0.1 g microcapsules in 10 mL simulated gastric juice (pH 2.0) for 1 h, followed by 10 mL simulated intestinal juice (pH 7.2) for 2 h under 37 °C anaerobic conditions. During this process, aliquots (0.1 mL) were taken from the mixture, serially diluted, and plated onto BHI agar plates for colony enumeration. The lyophilized powder of nonencapsulated AKK2 was used as a control, and the same treatment as the previous microcapsules was performed. Three replicates of each experiment were performed independently.
3.12. Stability of Microcapsules
The stability of microcapsule was studied after 0, 7, 14 and 28 d of storage at 4 °C or 25 °C under anaerobic (85% N2, 10% CO2, 5% H2) or aerobic conditions. The bacteria released from the microcapsule were enumerated after treatment with a 3% sodium citrate solution. The lyophilized powder of nonencapsulated bacteria was used as a control, and the same treatment as the previous microcapsules was performed.
3.13. Statistical Analyses
Data were presented as mean ± SEM. All statistical analyses were performed using GraphPad Prism 7.0 (GraphPad, San Diego, CA, USA). Statistical analysis was performed by one-way ANOVA with the Tukey–Kramer post hoc test and Student’s t-test. A p value of < 0.05 was considered significant, and p < 0.01 was considered highly statistically significant (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
4. Discussion
The prevalence of overweight/obesity has reached epidemic proportions. Compared with the classic mice or pig models, recent studies have reported the dog microbiome to be closer to that of a human [
37]. Thus, we first verified the anti-obesity function of
A. muciniphila AKK2, isolated from HFD-fed mice and observed its role in obese beagles. Results demonstrated the
A. muciniphila AKK2 treatment, tended to reduce HFD-induced obesity in both, which was manifested in significantly lower levels of weight gain, fasting blood glucose, serum TC, and LDL-C. The above results indicated that
A. muciniphila AKK2 is important for protection against HFD-induced obesity.
A great number of studies have revealed that
A. muciniphila and
Verrucomicrobia are very rare or not detected in the digestive tract of dogs [
38]. Some reasons might be inadequate sequence depth, the primers, 16S rRNA probes, or the reference sequence database [
39,
40]. This was also demonstrated by the very low relative abundance of
A. muciniphila in fecal samples from healthy and obese beagles in our study.
The gold measurement standards for diagnosing of type II diabetes mellitus are fasting blood glucose and the oral glucose tolerance test (OGTT). In this study, the concentration of blood glucose was significantly higher in the HFD-induced obese beagles than for those in the normal group but still within normal range. Therefore, we speculated that obesity and type II diabetes in beagles may not necessarily be connected. Nevertheless, these parameters decreased significantly after the AKK2 treatment, which further confirmed that it is important for regulating blood glucose.
Many studies have found that HFD intake induced obesity and varying degrees of liver damage. In this study, along with the increase in body weight of beagles, the biomarkers of liver function (GGT, ALT, and AST) were also higher in the HFD group than in the normal group. However, AKK2 reduced these parameters in the HFD group, indicating the hepatoprotective activity of the A. muciniphila AKK2 in obese beagles.
Numerous studies have demonstrated that obesity is closely associated with the gut microbiota, which directly affects food digestion, absorption, and metabolism. Dietary patterns and habits play an important role in the regulation of the intestinal flora. Accumulated evidence reveals that probiotics significantly alleviate obesity-related dysbacteriosis [
41]. In this study, fecal bacterial community sequencing showed that a high-fat diet alters gut microbiota composition and
A. muciniphila supplementation increased the
Firmicutes/Bacteroidetes (F/B) ratio in beagles. Furthermore, we found that obesity phenotypes such as weight gain and blood glucose concentration were inversely associated with the F/B ratio. At the phylum level,
A. muciniphila reversed the decreased abundance of F
irmicutes and
Fusobacteria and the increased abundance of
Actinobacteria, Proteobacteria, and
Bacteroidetes in obese beagles. At the genus level,
A. muciniphila increased the abundance of
Bifidobacterium and
Lactobacillus although the difference was not significant. Lu et al. observed that HFD feeding markedly decreased the abundance of
Lactobacillus reuteri,
Lactobacillus mucosa,
Lactobacillus johnsonii,
Prevotella copri,
and Clostridium perfringens [
42]. Therefore, the weight loss function of
A. muciniphila may be related to regulating the abundance of these bacteria.
A. muciniphila supplemented orally is confronted with multiple impediments owing to the complex digestive environment, such as variable pH and high bile salt content in the small intestine. Besides, A. muciniphila is highly sensitive to oxygen, which limits its further application. Thus, it is essential that A. muciniphila be properly protected in the development of any functional probiotic products.
The diverse preparation of microcapsules can protect probiotics against harsh environments and improve their fluidity to provide convenient conditions for later product development. Alginate is a biocompatible polymer, commonly used in the formation of hydrogels, microspheres, fibres, and microcapsules. Chitosan-linked alginate hydrogels could form a polyelectrolyte complex (PEC) to reduce particle porosity [
43,
44,
45]. Moreover, chitosan buffers excess protons in the gastric environment. In this study, we found that the abundance of
A. muciniphila in the intestine of beagles was extremely low. The bile acids produced by the duodenum had a degrading effect, which significantly reduced the number of viable bacteria, making it impossible to maintain continuous colonization. However, after AKK2 was encapsulated by chitosan and alginate its survival improved in simulated gastric fluid and the intestinal environment, and it became more stable under different storage conditions. Thus, we hypothesize that the administration of
A. muciniphila microcapsules was more conducive to the colonization of live bacteria in the gastrointestinal tract of animals.
In conclusion, this study revealed the beneficial role of anti-obesity treatments in beagles. A. muciniphila AKK2 promoted weight loss and ameliorated obesity-related metabolism in HFD-fed beagles. Moreover, A. muciniphila AKK2 restored HFD-induced changes in canine gut microbiota and promoted the abundance of probiotics, especially Bifidobacterium and Lactobacillus. These observations revealed that the anti-obesity effect of A. muciniphila in obese beagles was related to the regulation of intestinal flora. A better understanding of A. muciniphila may provide a new strategy for the effective treatment of overweight dogs.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}