Abstract
Cancer is a serious disease with high mortality and morbidity worldwide. Natural products have served as a major source for developing new anticancer drugs during recent decades. Magnolol, a representative natural phenolic lignan isolated from Magnolia officinali, has attracted considerable attention for its anticancer properties in recent years. Accumulating preclinical studies have demonstrated the tremendous therapeutic potential of magnolol via a wide range of pharmacological mechanisms against cancer. In this review, we summarized the latest advances in preclinical studies investigating anticancer properties of magnolol and described the important signaling pathways explaining its underlying mechanisms. Magnolol was capable of inhibiting cancer growth and metastasis against various cancer types. Magnolol exerted anticancer effects through inhibiting proliferation, inducing cell cycle arrest, provoking apoptosis, restraining migration and invasion, and suppressing angiogenesis. Multiple signaling pathways were also involved in the pharmacological actions of magnolol against cancer, such as PI3K/Akt/mTOR signaling, MAPK signaling and NF-κB signaling. Based on this existing evidence summarized in the review, we have conclusively confirmed magnolol had a multi-target anticancer effect against heterogeneous cancer disease. It is promising to develop magnolol as a drug candidate for cancer therapy in the future.
1. Introduction
Cancer is one of the serious diseases that threatens human life and health. The numbers of new cancer cases and deaths rose to 19.29 million and 9.96 million worldwide in 2020, in which Asia accounted for about the half of total numbers worldwide, with 9.50 million and 5.81 million, respectively [1,2]. Natural products used as traditional medicine have long been widely applied to treat cancer in Asian countries. In modern decades, natural products and their derivatives have served as the main source for developing new anticancer drugs such as paclitaxel, topotecan, and vinorelbine. According to recent statistics reported by Newman and Cragg, there are 39.2% approved anticancer drugs (102 out of 260 approved small molecule drugs) developed from natural products or their derivatives since 1981 [3]. Magnolia officinalis (Magnolia officinalis Rehd.et Wils) is a plant of the Magnolia family. According to the Chinese Pharmacopoeia (version 2020), the dried stem bark, root bark, and branch bark of Magnolia officinalis plant are named Houpu and have the medicinal uses of drying Dampness, transforming Phlegm and descending Rebellious Qi. Magnolol (5,5′-diallyl-2,2′-dihydroxybiphenyl) and its structural analogue, honokiol (3,5′-diallyl-4,2′-dihydroxybiphenyl), are the main active ingredients of Magnolia officinalis bark extract, which also are the representative compounds of phenolic lignan derived from natural products (Figure 1). Both magnolol and honokiol have been reported to exert various biological properties in recent decades, including anticancer, antioxidative, anti-inflammatory, and many other activities. The unique symmetrical diphenol structure of magnolol plays a crucial role in its bioactivity by facilitating the activation of the interactions with the surface of proteins [4,5]. Since magnolol and honokiol are isomers of biphenyl-type neolignan and share similar chemical structure, and the only difference between two compounds is the position of one hydroxyl group. That is, they exhibit very close biological properties but with slight activity difference. For example, in 2,2′-Azobis (2-amidinopropane) dihydrochloride (AAPH)-induced DNA oxidation, both magnolol and honokiol demonstrated good antioxidant capacities and could trap 1.8 and 2.5 radicals in protecting DNA, respectively [6]. Their activity difference was suggested to be related to the intramolecular hydrogen bond formed between di-ortho-hydroxyl groups, which affected the hydrogen atom in hydroxyl group to be abstracted by radicals [6]. Magnolol and honokiol both have showed good anticancer effects in a number of studies, but are with variable activities in different types of cancer cells. There are already several literature reviews to summarize the anticancer role of honokiol against cancer [7,8,9,10,11,12], but few for magnolol. This paper highlights the potential anticancer effect of magnolol. In recent years, there are a number of studies reporting the anticancer effect of magnolol, which exhibited a wide range of pharmacological mechanisms against cancer, including proliferation inhibition, cell cycle arrest, apoptosis induction, metastasis blockage, angiogenesis inhibition and signaling pathways involving phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin (PI3K/Akt/mTOR) signaling, mitogen-activated protein kinase (MAPK) signaling and nuclear factor kappa-B (NF-κB) signaling. In this review, we will summarize the latest advances in the anticancer properties of magnolol and its potential mechanistic actions.
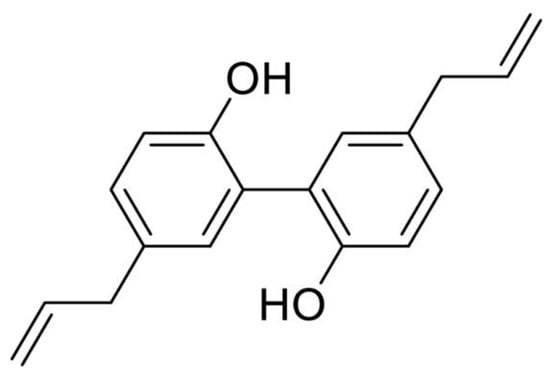
Figure 1.
Chemical structure of magnolol.
2. Search Syntax and Inclusion Criteria
Relevant published studies were identified for the years 2002–2022 by searching PubMed database with general string of “(magnolol) AND (cancer OR tumor OR tumour OR carcinoma)”. The citations used in this study were searched by August 31, 2022. The pre-set criteria for inclusion were: (i) the study investigated the anticancer effect of magnolol, not its analogues or derivatives; (ii) the study attempted to elucidate the underlying anticancer mechanism of magnolol; (iii) the study was pharmacological investigation, not pharmaceutical research (e.g., drug delivery system); (iv) only publication in English was included; and (v) other relevant citations not included in the search syntax were manually searched.
3. Magnolol Inhibits Proliferation and Growth
Unlimited proliferation is one of the fundamental cancer characteristics, thus restricting tumor cell proliferation represents a promising direction for developing anticancer therapeutics. In recent years, magnolol has exhibited the antiproliferative property against various cancer types. Magnolol effectively inhibited tumor cell growth in bladder cancer [13,14], breast cancer [15,16], cholangiocarcinoma [17], colon cancer [18,19,20,21,22,23], esophagus cancer [24], fibrosarcoma [25], gallbladder cancer [26], gastric cancer [27], glioblastoma [28,29,30,31,32], leukemia [25], liver cancer [21,33,34,35,36], lung cancer [37,38,39,40], lymphoma [41], melanoma [25,42,43], myeloma [44], oral carcinoma [45,46], osteosarcoma [47], ovarian cancer [48], pancreatic cancer [49], prostate cancer [50,51,52,53], renal cancer [54,55], skin cancer [56,57] and thyroid cancer [58] (Table 1). In terms of treatment efficacy, the in vitro IC50 values of magnolol for the majority of cancer types varied from 20 to 100 μM under 24-h treatment. The minimum effective inhibitory dose of magnolol in vitro was 2.4 μM as demonstrated in cancer stem cells enriched from oral squamous cell carcinoma [45]. For in vivo mouse studies, 5 mg/kg magnolol was sufficient to restrict tumor growth in bladder cancer [14], colon cancer [19] and gallbladder cancer [26]. The IC50 values of magnolol on different cancer cell lines as well as the effective doses used in tumor-bearing mice were summarized in Table 1. Several cellular signaling have been uncovered to participate in antiproliferative effect of Magnolol against cancer, as summarized in Table 1 and Figure 2, mainly involving with PI3K/Akt/mTOR and MAPK/ERK signaling. Meanwhile, the provocation of cell cycle arrest and apoptosis also demonstrate crucial roles on suppressing tumor growth, they are associated with magnolol-induced MAPK/JNK and MAPK/p38 cascades activation, as well as NF-κB signaling blockage. We introduce them in the following paragraphs.

Table 1.
The anticancer effect of magnolol against various cancer types.
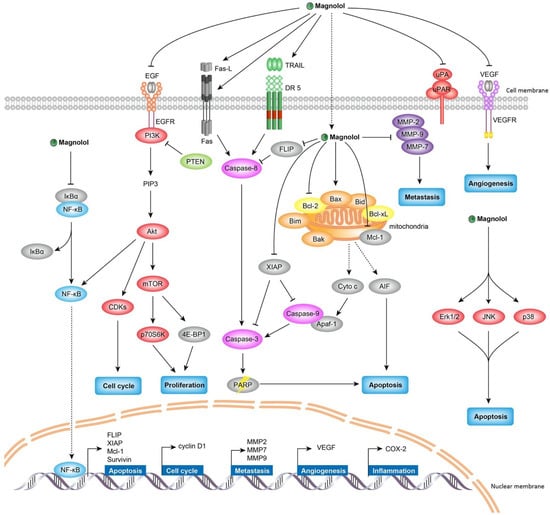
Figure 2.
The anticancer mechanism of magnolol. Magnolol exerts anticancer effects through inhibiting proliferation, inducing cell cycle arrest, provoking apoptosis, restraining migration and invasion, and suppressing angiogenesis. Multiple signaling pathways are involved in the pharmacological actions of magnolol against cancer, such as PI3K/Akt/mTOR signaling, MAPK signaling, and NF-κB signaling.
4. Magnolol Inhibits PI3K/Akt/mTOR Signaling
The PI3K/Akt/mTOR signaling has been found to be frequently dysregulated pathways of cancer cell proliferation and survival [59]. PI3K catalyzes the conversion of phosphatidylinositol (3,4)-bisphosphate into phosphatidylinositol (3,4,5)-trisphosphate (PIP3), therefore triggers the phosphorylation of Akt [60], subsequently activates mTOR complex 1 (mTORC1) via phosphorylation [61]. It is known that mTOR controls the expression of many proliferation promoting proteins via phosphorylating the p70S6 kinase (p70S6K) and the eukaryotic translation initiation factor 4E binding protein 1 (4E-BP1) [62]. Many studies have reported that magnolol inhibited tumor cell proliferation through attenuating the PI3K/Akt/mTOR signaling. Magnolol has been observed to diminish the phosphorylation of PI3K, Akt, mTOR, p70S6K and 4E-BP1, and this magnolol-induced inactivation of PI3K/Akt/mTOR signaling seems to be cancer type-independent, across a variety of cancer types covering bladder cancer [14], colon cancer [20], gastric cancer [27], glioblastoma [31], lung cancer [37,40], melanoma [43], ovarian cancer [48], prostate cancer [50] and skin cancer [57] (Table 1 and Figure 2). The PI3K/Akt/mTOR signaling usually is activated by ligand binding tyrosine kinase receptors, and PTEN negatively regulates the cascade. Magnolol inactivated the tyrosine kinase receptor EGFR and VEGFR2 [14,50] as well as upregulating PTEN activity, thereby abolishing the abnormal PI3K/Akt/mTOR signaling in cancer cells [38,58]. It is considered that the PI3K/Akt/mTOR cascade serves as a major antiproliferative signaling pathway responsible for magnolol-induced tumor growth inhibition.
5. Magnolol Inhibits MAPK Signaling
The MAPK signaling pathway is an important signal transduction pathway that regulates many cellular processes, and its dysregulation leads to aberrant control of cell proliferation, survival and apoptosis [63]. There are three main classical MAPK cascades involving MAPK/extracellular signal-regulated kinase (ERK), MAPK/C-Jun N-Terminal Kinase (JNK) and MAPK/p38 mitogen-activated protein kinase (p38) [64]. All three of these cascades have been reported to be targeted by magnolol to exert anticancer properties as revealed by multiple studies [13,18,22,23,24,30,34,35,36,37,41,43,46,48,56,57], however whether the modulating effect of magnolol on different MAPK cascades is upregulated or downregulated remains controversial (Table 1 and Figure 2). Magnolol could activate MAPK/ERK cascade by increasing the phosphorylation of ERK to provoke and apoptosis and cell cycle arrest in bladder cancer [13], colon cancer [18], esophagus cancer [24], glioblastoma [30], lymphoma [41] and skin cancer [56]. The similar programmed cell death was also observed mediating by magnolol-induced MAPK/JNK and MAPK/p38 activation in bladder cancer [13], esophagus cancer [24], glioblastoma [30], lung cancer [37] and oral carcinoma [46]. However, the MAPK signaling pathway could play a dual role in regulating cancer inhibition process. As reported by other studies, magnolol was capable of hindering NF-κB activation and cell survival through the downregulation of MAPK/ERK cascade [23,34,36,43,48,57]. Additionally, magnolol was also found to block cancer migration and invasion via suppressing ERK and p38 cascades [22,23,34,48]. Therefore, how MAPK cascades acts on magnolol-induced anticancer process remains controversial, it might be due to the reason that the MAPK pathway is predominantly stimulated by various stress signaling and plays a central role in regulating diverse downstream.
6. Magnolol Inhibits NF-κB Signaling
NF-κB signaling plays a fundamental role in cancer development and progression, its aberrant activation has been found to be associated with multiple pathways in malignant tumors, including apoptosis inhibition, survival promotion and angiogenesis induction [65,66]. NF-κB activation also triggers epithelial-mesenchymal transition (EMT) to promote cancer migration and invasion [67]. The classic form of NF-κB is the heterodimer of the p50 and p65 subunits, which is controlled by its biological inhibitor IκBs by modulating the translocation of NF-κB from cytoplasm to nucleus. NF-κB is activated by the phosphorylation-induced degradation of IκBs, which facilitates the dissociation of NF-κB from IκB family proteins in cytoplasm [67]. In various cancer types, magnolol exerted the inhibitory effect on NF-κB activity, which is demonstrated by studies of breast cancer [15], cholangiocarcinoma [17], colon cancer [23], liver cancer [34,36], lung cancer [39], myeloma [44], ovarian cancer [48] and skin cancer [57] (Table 1 and Figure 2). For the majority of cases, the inactivation of NF-κB signaling induced by magnolol was to block the phosphorylation of IκBα and/or p65 subunit of NF-κB. Followed by direct NF-κB activity inactivation, magnolol also affected the expression of NF-κB targeting genes involved in apoptosis (Survivin, XIAP, c-FLIP and Mcl-1) [23,34,36], cell cycle (cyclin D1) [17,23,36], metastasis (MMP2, MMP7 and MMP9) [17,23,34,36,44], angiogenesis (VEGF) [23] and inflammation (COX-2) [57]. In general, the magnolol-affected NF-κB signaling inhibition plays crucial functions on regulating tumor growth and metastasis. In particular, it is considered as the primary anti-metastatic signaling of magnolol [15,17,34,44].
7. Magnolol Induces Cell Cycle Arrest
Cell cycle is tightly regulated by organized checkpoints to maintain normal cell division and proliferation. Many key checkpoint regulators of cell cycle have been identified, which can be classified as three main categories: cyclin, cyclin-dependent kinase (CDK) and cyclin-dependent kinase inhibitor (CKI). They coordinate with each other to constitute a complex regulatory network in the cell cycle. There are four major cyclins involved in cell cycle control, including cyclin A, B1, D1 and E. Entry into the S phase from G1 phase requires cyclin E and D1 activation, cyclin A facilitates the progression through S phase. Cyclin B1 is an essential cyclin to bring cells into mitosis (M) stage. The cyclins form a complex with its corresponding CDKs (CDK1, 2, 4 or 6) to promote cell cycle progression, whereas CKIs, such as p15, p16, p21 and p27, negatively regulate the cell cycle through competing with corresponding cyclins for the binding to CDKs [68]. Abnormal cell cycle regulatory protein expression would lead to unlimited cell replication, which is a hallmark of cancer [69]. Magnolol has been shown to induce cell cycle arrest, consequently leading to apoptosis and cell death in multiple cancer types. Interestingly, it was found that magnolol could trigger cell cycle arrest at three different phases involving sub-G1 [27,34,39,46,55], G0/G1 [13,17,18,21,26,29,30,35,43,47,52,54] and G2/M [13,16,40,56] (Table 1 and Figure 2). Their difference seemed to be magnolol dose-dependent and cancer type-related. As demonstrated in bladder cancer, magnolol at low dose (20 and 40 μM) could induce cell cycle arrest at G0/G1 phase, only when the dose increasing to 60 μM, cell cycle of G2/M phase started to be arrested [13]. Consistent with this finding, the majority of magnolol doses which could induce G0/G1 cell cycle arrest were at the range of 10–40 μM in various other cancer types, including cholangiocarcinoma [17], colon cancer [21], gallbladder cancer [26], glioblastoma [29], liver cancer [35], melanoma [43], osteosarcoma [47] and renal cancer [54]. By comparison, the doses of magnolol trigging G2/M arrest were much higher, with 75–100 μM in skin cancer and 80–100 μM in breast cancer [16,56]. In addition, magnolol could also induce cell cycle arrest at sub-G1 phase in gastric cancer [27], liver cancer [34], lung cancer [39], oral cancer [46] and renal cancer [55]. There was no specific effective dose preference for sub-G1 arrest induction found from these studies, indicating the cancer type difference, mainly caused by genetic background heterogeneity, might also contribute to important influence on how magnolol regulates cell cycle arrest.
8. Magnolol Induces Apoptosis
Apoptosis is a strictly programmed cell death process that plays a critical role in physiological and pathological activities. Resisting cell death is a hallmark of cancer development and progression [69]. Therefore, promoting cancer cell death by triggering apoptosis has become a feasible strategy for anticancer therapy. Apoptotic pathways generally can be divided by intrinsic (mitochondrial-mediated) and extrinsic (death receptor-mediated) pathways. The intrinsic pathway is activated by cytochrome c released from mitochondria, which is regulated by the Bcl-2 family members and caspases. The Bcl-2 family includes both pro-apoptotic (e.g., Bax, Bak, Bid, Bad and Bim) and anti-apoptotic (e.g., Bcl-2, Bcl-x and Bcl-XL) proteins [70,71,72]. The intrinsic pathway is usually initiated by intracellular stress stimuli and requires the activation of caspase-9. The extrinsic pathway is initiated by the activation of death receptors, such as the receptors of FASL, TNF, and TRAIL, which are associated with procaspase-8 through the dimerization of the death effect domain, leading to autocatalytic activation of procaspase-8 and triggering apoptosis [73]. Both intrinsic and extrinsic apoptosis induce cleavage caspase family, such as caspase-3, caspase-6 and caspase-7. The apoptotic effects of magnolol have been widely investigated in many studies, meanwhile different cellular mechanisms elucidating magnolol-triggered apoptosis have been explored. The major apoptotic process induced by magnolol was initiated by the mitochondria-dependent intrinsic pathway. Magnolol induced intrinsic apoptosis in various types of cancer (Table 1 and Figure 2), including breast cancer [16], colon cancer [20], glioblastoma [31], liver cancer [34,35,36], lung cancer [37,39,40], lymphoma [41], melanoma [42], osteosarcoma [47], ovarian cancer [48], and thyroid cancer [58]. In general, magnolol significantly decreased Bcl-2 expression and increased Bax expression, leading to an upregulation of the Bax/Bcl-2 ratio; it also induced the release of cytochrome c, increased PARP cleavage, and consequently induced apoptosis in cancer cells. Several studies had found that magnolol-induced intrinsic apoptosis via cleaved-caspase-9, for example, in glioblastoma [31], liver cancer [36], lung cancer [40] and renal cancer [54]. The activated caspase-9 induced the cleavage of downstream caspases-3, which subsequently induced apoptosis. In addition, magnolol also induced the mitochondria to nucleus translocation of apoptosis-inducing factor (AIF) and resulted in apoptosis [16,37]. In addition to the intrinsic apoptosis, magnolol could also provoke the extrinsic apoptotic pathway in fibrosarcoma [25], glioblastoma [28], leukemia [25], melanoma [42], renal cancer [55] and skin cancer [56] (Table 1 and Figure 2). Magnolol was able to induce death receptor mediated caspase-8 activation in a concentration-dependent manner, subsequently activated caspase-3 and PARP by cleavage and resulted in apoptosis [28,55,56]. Furthermore, the downregulated expression of the anti-apoptotic protein Mcl-1, c-FLIP, XIAP and the upregulated expression of pro-apoptotic protein Apaf-1, ATF4 also have been demonstrated to involve in magnolol-induced apoptosis [33,34,47,54,55]. Interestingly, it was found that magnolol could simultaneously activate both intrinsic mitochondria-dependent and extrinsic death receptor-mediated apoptotic pathways against cancer, such as in colon cancer [23], esophagus cancer [24], glioblastoma [28], liver cancer [33], melanoma [42] and oral carcinoma [46]. The intrinsic apoptotic signaling transduction (the loss of mitochondrial membrane potential and cleaved caspase-9) and the extrinsic apoptotic signaling transduction (the activation of FAS, FASL, DR-4, DR-5 and cleaved caspase-8) could be significantly triggered by magnolol at a same time [28,55].
9. Magnolol Inhibits Migration and Invasion
Metastasis is the main lethal factor in cancer patients. It involves a sequential cascade, including the migration and invasion of regional tumor cells into neighboring tissues, intravasation into blood or lymphatic vessels, and extravasation into distant organs [74]. To degrade extracellular matrix (ECM) of surrounding tissues and transform tumor cells into invasive mesenchymal phenotype are essential processes involved in cancer cell migration and invasion [75]. Matrix metalloproteinases (MMPs) are calcium endopeptidases that promote ECM degradation [76]. Overexpression of MMPs has been found to be highly associated with tumor metastasis progression [77]. EMT is a critical process to make epithelial tumor cells obtaining the features of invasiveness [78]. The EMT processes manifest as upregulation of mesenchymal markers (α-SMA, fibronectin, N-cadherin, Snail, Slug, Twist and Vimentin) and downregulation of epithelial markers (E-cadherin, cytokeratin, and occludin) [79]. Various studies have reported that magnolol had the capability to suppress cancer cell migration and invasion in different cancer types, including breast cancer [15], cholangiocarcinoma [17], colon cancer [19,20,22,23], esophagus cancer [24], glioblastoma [28,32], liver cancer [33,34,35], myeloma [44], oral carcinoma [45], osteosarcoma [47], ovarian cancer [48], pancreatic cancer [49], prostate cancer [53], and renal cancer [54] (Table 1). These studies proved that magnolol inhibited migration and invasion by decreasing the expressions and activities of MMPs, such as MMP-2, MMP-7, MMP-9, and urokinase-type plasminogen activator (uPA) [15,17,19,23,24,28,33,34,44]. Magnolol could remarkably block EMT by downregulating Vimentin, N-Cadherin, Twist, Slug, Snail and upregulating E-cadherin [19,22,32,49]. A number of studies indicated that several upstream signaling cascades were involving in the modulation of MMPs and EMT by magnolol. Magnolol diminished NF-κB activation by downregulating phosphorylation of p65 and IκBα, which was considered as the primary anti-metastatic signaling of magnolol as evidenced in multiple cancer types [15,17,34,44]. Magnolol also inhibited transforming growth factor beta 1 (TGF-β1)/Smad signaling [22,49], β-Catenin/transcription factor (TCF) signaling [19], protein kinase C delta (PKCδ)/signal transducer and activator of transcription 3 (STAT3) signaling [28]. Additionally, magnolol decreased focal adhesion-related protein expression through attenuating focal adhesion kinase (FAK) and paxillin phosphorylation [32].
10. Magnolol Suppresses Angiogenesis
Angiogenesis, the formation of new blood vessels, is critical for supplying nutrients and oxygen required for tumor progression [80]. Tumor angiogenesis is a complex process in which tumor cells interact with endothelial cells. Vascular endothelial growth factor (VEGF) has been described as an essential angiogenic factor that plays a fundamental role in new blood vessel generation [10]. Secreted VEGF interacts with its specific receptors (VEGFR) to initiate endothelial cell proliferation, migration, and colonization, thus to promote angiogenesis. Intratumoral hypoxia is believed to be a main cause of angiogenesis that is closely associated with tumor growth and metastasis [81,82]. Under hypoxic condition, hypoxia-inducible factor 1α (HIF-1α) has been regarded as the primary regulator facilitating angiogenesis by upregulating pro-angiogenic genes in tumor cells, such as VEGF [83]. Thus, the inhibition of hypoxia-mediated tumor angiogenesis is considered a feasible approach to attenuating cancer progression. Magnolol has been found to have effective antiangiogenic activities (Table 1). Specifically, magnolol suppressed the proliferation and tube formation of human umbilical endothelial cells (HUVECs) and mouse embryonic stem (MES)/embryoid body (EB)-derived endothelial-like cells [14,84,85] and inhibited vascular branch formation in chicken chorioallantoic membrane (CAM) [14]. Similar anti-angiogenic evidence was also proved by in vivo studies that magnolol blocked vascular outgrowth in bladder cancer xenografts and subsequently restrained tumor growth [14]. In endothelial cells, magnolol targeted VEGFR-dependent angiogenesis via repressing PI3K/Akt/mTOR and MAPK signaling pathway and inducing ROS-mediated apoptosis [85]. On the other hand, magnolol also acted as an important role in tumor cells through reducing hypoxia /HIF-1α-mediated VEGF secretion [14]. Hypoxia-associated HIF-1α was the primary transcription factor of VEGF. In bladder cancer cells, the hypoxia-induced HIF-1α activation and VEGF secretion were abolished by magnolol through suppressing VEGFR2/PI3K/Akt/mTOR/p70S6K/4E-BP1 cascade [14]. Meanwhile, the magnolol-mediated inhibition of HIF-1α protein expression is associated with increased HIF-1α protein degradation and protein synthesis blockage [14].
11. Synthetic Analogues of Magnolol and Their Anticancer Activities
To enhance the anticancer efficacy of magnolol, several structural modification studies have been investigated (Figure 3). Magnolol analogue 5,5′-dibutyl-3,3′-diiodo-[1,1′-biphenyl]-2,2′-diol (named as Ery5, (a)) exhibited strong antiproliferative activity with over 20-fold increase against HL-60 and PC-3 cancer cell lines compared with magnolol [86]. Ery5 could effectively induce cell cycle arrest at Sub-G1, trigger autophagic cancer cell death and inhibit angiogenesis [86,87,88]. Xu et al. synthesized a serious of magnolol-based Mannich base derivatives by introducing Mannich base groups at the ortho position of the phenolic hydroxyl at C-2′ [89]. Most of these magnolol derivatives displayed more potent antiproliferative activity against cancer cells than magnolol. Among them, 5,5′-diallyl-3-(morpholino(3,4,5-trimethoxyphenyl)methyl)-[1,1′-biphenyl]-2,2′-diol (compound 3p, (b)) had the most potent anticancer activity against T47D cancer cells with 76.1-fold enhancement of cytotoxic effect compared with magnolol [89]. Tang et al. introduced nitrogen heterocycle at C-3 position of magnolol in order to improve the water solubility and antitumor activity [90]. Magnolol analogue 5,5′-diallyl-3-((4-aminopiperidin-1-yl)methyl)-[1,1′-biphenyl]-2,2′-diol (compound C2, (c)) exhibited significant and broad-spectrum antiproliferative activity with approximately 8-fold more potent than magnolol in NSCLC cell lines [90]. Based on the core of compound C2, Zhao et al. continued to synthesize 5,5′-diallyl-3-((4-aminopiperidin-1-yl)methyl)-2′-isopropoxy-[1,1′-biphenyl]-2-ol (compound 30, (d)) with a piperityl group substituting the hydrogen atom at C-3 position of magnolol, which exhibited potent antiproliferative activities on H460, HCC827 and H1975 cancer cell lines with approximately 100-fold more potent than magnolol [91]. Chen et al. synthesized magnolol analogue 2-(4-((2-((5′,6-diallyl-2′-hydroxy-[1,1′-biphenyl]-3-yl)oxy)ethyl)sulfinyl)butyl)isoindoline-1,3-dione (compound CT2-3, €), which showed significant cytotoxicity against A549 and H460 cancer cell lines [92]. But the antiproliferative activity of compound CT2-3 was not as strong as the analogues synthesized by other research groups, only 2-fold better than magnolol [92]. Tao et al. synthesized a magnolol-sulforaphane hybrid 2′,5-diallyl-5′-(2-((4-isothiocyanatobutyl)sulfinyl)ethoxy)-[1,1′-biphenyl]-2-ol (compound CT1-3, (f)), exhibiting at least 5-fold more efficient anticancer activity than magnolol in multiple cancer cell lines [93]. More recently, Sun et al. designed and synthesized a series of magnolol derivatives showing better cytotoxic activity compared to magnolol [94]. Among them, 5,5′-diallyl-2′-((4-fluorobenzyl)oxy)-[1,1′-biphenyl]-2-ol (compound 6a, (g)) was the most potent one to inhibit cancer cell proliferation, migration and invasion. The results suggested that substitution of the benzyl group with F atoms at the C2 position on magnolol was a viable strategy for structural optimization [94].
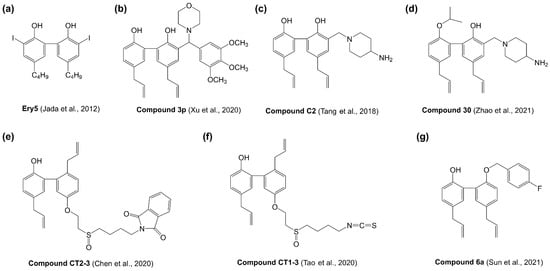
Figure 3.
The structure of magnolol synthetic analogues. (a) 5,5′-dibutyl-3,3′-diiodo-[1,1′-biphenyl]-2,2′-diol; (b) 5,5′-diallyl-3-(morpholino(3,4,5-trimethoxyphenyl)methyl)-[1,1′-biphenyl]-2,2′-diol; (c) 5,5′-diallyl-3-((4-aminopiperidin-1-yl)methyl)-[1,1′-biphenyl]-2,2′-diol; (d) 5,5′-diallyl-3-((4-aminopiperidin-1-yl)methyl)-2′-isopropoxy-[1,1′-biphenyl]-2-ol; (e) 2-(4-((2-((5′,6-diallyl-2′-hydroxy-[1,1′-biphenyl]-3-yl)oxy)ethyl)sulfinyl)butyl)isoindoline-1,3-dione; (f) 2′,5-diallyl-5′-(2-((4-isothiocyanatobutyl)sulfinyl)ethoxy)-[1,1′-biphenyl]-2-ol; (g) 5,5′-diallyl-2′-((4-fluorobenzyl)oxy)-[1,1′-biphenyl]-2-ol.
12. Conclusions and Prospects
Cancer is a serious malignant disease with high morbidity and mortality worldwide. As far as we know, the complexity of tumor heterogeneity brings cancer to be a multifactorial disease, which involves in various signaling pathways during tumor initiation, progression and recurrence [95]. Therefore, to develop new drugs with multiple targets against cancer has become a promising and popular trend. Chinese herbal medicines as anticancer drug discovery sources have attracted increasing attentions in recent years because they have been demonstrated with advantages of evidence-based effectiveness, multi-target therapy, and few side effects. Accumulating preclinical studies have proved the tremendous anticancer potential of magnolol, a representative compound derived from Chinese herbal medicines, as summarized in the present review. Magnolol exerted anticancer effects through various molecular mechanisms, including proliferation, cell cycle, apoptosis, metastasis, angiogenesis and so on. Multiple signaling pathways were also involved in the pharmacological mechanisms of magnolol against cancer, such as PI3K/Akt/mTOR signaling, MAPK signaling and NF-κB signaling (Table 1 and Figure 2). Based on this evidence, the multi-target anti-cancer properties of magnolol have been initially confirmed as supporting its druggable potential against heterogeneous cancer. Furthermore, unlike chemotherapy, either magnolol or Magnolia bark extract had no known toxicities and side effects reported so far, which makes this natural agent a potentially safe drug candidate. In addition, magnolol has the ability to successfully cross the blood-brain barrier and rapidly distribute into different brain regions [96], which expands its pharmacological potential against brain tumors. However, it is worth noting that the water solubility, gastrointestinal absorption and first-pass metabolism of magnolol are poor, with oral bioavailability less than 10% [97], which restricts its clinical application. Despite the promising results on anticancer activity of magnolol, further research is required to improve its bioavailability, including to investigate appropriate liposomal formulation, nanocarrier delivery system and chemical structure modification to enhance magnolol in vivo efficacy [98,99,100]. And so far, no clinical evidence has been obtained to show the efficacy of magnolol. To fully realize the therapeutic potential of magnolol, more clinical investigations are necessary. Nevertheless, based on existing preclinical studies, we have reason to believe that magnolol may potentially be developed as a promising natural agent to fight against cancer. To sum up, we have reviewed the anticancer effects of magnolol against various cancer types and elucidated its main underlying pharmacological mechanisms, including proliferation inhibition, cell cycle arrest, apoptosis induction, metastasis blockage, angiogenesis inhibition, and pathways involving in PI3K/Akt/mTOR, MAPK and NF-κB signaling (Table 1 and Figure 2). The preclinical studies on magnolol are intriguing and suggest it as a promising multi-target drug candidate for cancer therapy.
Author Contributions
Conceptualization, J.B. and Z.S.; writing—original draft preparation, X.W. and Q.L.; writing—review and editing, J.B., Z.S., X.W., Q.L., Y.F., R.-B.D., X.Q. and X.Z.; funding acquisition, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82104057 and 82203206), Hainan Provincial Natural Science Foundation of China (No. 820QN248, 822RC649 and 822QN300), the Scientific Research Foundation of Hainan University (No. KYQD(ZR)-21030 and KYQD(ZR)-21169), the Open Research Project Program of the State Key Laboratory of Quality Research in Chinese Medicine (University of Macau) (No. SKL-QRCM-OP21009) and the Hainan Association for Science and Technology Innovation Program of Youth Talents (QCQTXM202210).
Acknowledgments
We thank members of the Bao and Sun laboratories for helpful advice and discussion.
Conflicts of Interest
The authors declare no competing interests.
Abbreviations
| PI3K | phosphoinositide 3-kinase |
| Akt | protein kinase B |
| mTOR | mammalian target of rapamycin |
| MAPK | mitogen-activated protein kinase |
| NF-κB | nuclear factor kappa-B |
| PIP3 | phosphatidylinositol (3,4,5)-trisphosphate |
| mTORC1, | mTOR complex 1 |
| p70S6K | p70S6 kinase |
| 4E-BP1 | eukaryotic translation initiation factor 4E binding protein 1 |
| ERK | extracellular signal-regulated kinase |
| JNK | C-Jun N-Terminal Kinase |
| EMT | epithelial-mesenchymal transition |
| CDK | cyclin dependent kinase |
| CKI | cyclin dependent kinase inhibitor |
| AIF | apoptosis-inducing factor |
| ECM | extracellular matrix |
| MMPs | matrix metalloproteinases |
| uPA | urokinase-type plasminogen activator |
| TGF-β1 | transforming growth factor beta 1 |
| TCF | transcription factor |
| PKCδ | protein kinase C delta |
| STAT3 | signal transducer and activator of transcription 3 |
| FAK | focal adhesion kinase |
| VEGF | vascular endothelial growth factor |
| HIF-1α | hypoxia-inducible factor 1α |
| HUVECs | human umbilical endothelial cells |
| MES | mouse embryonic stem |
| EB | embryoid body |
| CAM | chicken chorioallantoic membrane |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Cao, W.; Chen, H.D.; Yu, Y.W.; Li, N.; Chen, W.Q. Changing profiles of cancer burden worldwide and in China: A secondary analysis of the global cancer statistics 2020. Chin. Med. J. (Engl. Ed.) 2021, 134, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, G.; Gulder, T.; Gulder, T.A.; Breuning, M. Atroposelective total synthesis of axially chiral biaryl natural products. Chem. Rev. 2011, 111, 563–639. [Google Scholar] [CrossRef] [PubMed]
- Hajduk, P.J.; Bures, M.; Praestgaard, J.; Fesik, S.W. Privileged molecules for protein binding identified from NMR-based screening. J. Med. Chem. 2000, 43, 3443–3447. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, Z.Q. Comparison of antioxidant abilities of magnolol and honokiol to scavenge radicals and to protect DNA. Biochimie 2011, 93, 1755–1760. [Google Scholar] [CrossRef] [PubMed]
- Banik, K.; Ranaware, A.M.; Deshpande, V.; Nalawade, S.P.; Padmavathi, G.; Bordoloi, D.; Sailo, B.L.; Shanmugam, M.K.; Fan, L.; Arfuso, F.; et al. Honokiol for cancer therapeutics: A traditional medicine that can modulate multiple oncogenic targets. Pharmacol. Res. 2019, 144, 192–209. [Google Scholar] [CrossRef]
- Pan, J.; Lee, Y.; Wang, Y.; You, M. Honokiol targets mitochondria to halt cancer progression and metastasis. Mol. Nutr Food Res. 2016, 60, 1383–1395. [Google Scholar] [CrossRef]
- Arora, S.; Singh, S.; Piazza, G.A.; Contreras, C.M.; Panyam, J.; Singh, A.P. Honokiol: A novel natural agent for cancer prevention and therapy. Curr. Mol. Med. 2012, 12, 1244–1252. [Google Scholar] [CrossRef]
- Ong, C.P.; Lee, W.L.; Tang, Y.Q.; Yap, W.H. Honokiol: A Review of Its Anticancer Potential and Mechanisms. Cancers 2019, 12, 48. [Google Scholar] [CrossRef]
- Rauf, A.; Patel, S.; Imran, M.; Maalik, A.; Arshad, M.U.; Saeed, F.; Mabkhot, Y.N.; Al-Showiman, S.S.; Ahmad, N.; Elsharkawy, E. Honokiol: An anticancer lignan. Biomed. Pharmacother. 2018, 107, 555–562. [Google Scholar] [CrossRef]
- Fried, L.E.; Arbiser, J.L. Honokiol, a multifunctional antiangiogenic and antitumor agent. Antioxid. Redox Signal. 2009, 11, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, Y.H.; Park, K.; Kim, E.J.; Jung, K.H.; Park, S.S.; Kim, W.J.; Moon, S.K. Magnolol elicits activation of the extracellular signal-regulated kinase pathway by inducing p27KIP1-mediated G2/M-phase cell cycle arrest in human urinary bladder cancer 5637 cells. Biochem. Pharmacol. 2008, 75, 2289–2300. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Lee, C.F.; Huang, W.H.; Chou, T.C. Magnolol suppresses hypoxia-induced angiogenesis via inhibition of HIF-1alpha/VEGF signaling pathway in human bladder cancer cells. Biochem. Pharmacol. 2013, 85, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, W.; Zhang, B.; Liu, Y.Q.; Wang, Z.Y.; Wu, Y.P.; Yu, X.J.; Zhang, X.D.; Ming, P.H.; Zhou, G.B.; et al. The natural compound magnolol inhibits invasion and exhibits potential in human breast cancer therapy. Sci. Rep. 2013, 3, 3098. [Google Scholar] [CrossRef]
- Zhou, Y.; Bi, Y.; Yang, C.; Yang, J.; Jiang, Y.; Meng, F.; Yu, B.; Khan, M.; Ma, T.; Yang, H. Magnolol induces apoptosis in MCF-7 human breast cancer cells through G2/M phase arrest and caspase-independent pathway. Pharmazie 2013, 68, 755–762. [Google Scholar]
- Zhang, F.H.; Ren, H.Y.; Shen, J.X.; Zhang, X.Y.; Ye, H.M.; Shen, D.Y. Magnolol suppresses the proliferation and invasion of cholangiocarcinoma cells via inhibiting the NF-κB signaling pathway. Biomed. Pharmacother. 2017, 94, 474–480. [Google Scholar] [CrossRef]
- Hsu, Y.F.; Lee, T.S.; Lin, S.Y.; Hsu, S.P.; Juan, S.H.; Hsu, Y.H.; Zhong, W.B.; Lee, W.S. Involvement of Ras/Raf-1/ERK actions in the magnolol-induced upregulation of p21 and cell-cycle arrest in colon cancer cells. Mol. Carcinog. 2007, 46, 275–283. [Google Scholar] [CrossRef]
- Kang, Y.J.; Park, H.J.; Chung, H.J.; Min, H.Y.; Park, E.J.; Lee, M.A.; Shin, Y.; Lee, S.K. Wnt/beta-catenin signaling mediates the antitumor activity of magnolol in colorectal cancer cells. Mol. Pharmacol. 2012, 82, 168–177. [Google Scholar] [CrossRef]
- Park, J.B.; Lee, M.S.; Cha, E.Y.; Lee, J.S.; Sul, J.Y.; Song, I.S.; Kim, J.Y. Magnolol-induced apoptosis in HCT-116 colon cancer cells is associated with the AMP-activated protein kinase signaling pathway. Biol. Pharm. Bull. 2012, 35, 1614–1620. [Google Scholar] [CrossRef]
- Lin, S.Y.; Liu, J.D.; Chang, H.C.; Yeh, S.D.; Lin, C.H.; Lee, W.S. Magnolol suppresses proliferation of cultured human colon and liver cancer cells by inhibiting DNA synthesis and activating apoptosis. J. Cell Biochem. 2002, 84, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Chei, S.; Oh, H.J.; Song, J.H.; Seo, Y.J.; Lee, K.; Lee, B.Y. Magnolol Suppresses TGF-β-Induced Epithelial-to-Mesenchymal Transition in Human Colorectal Cancer Cells. Front. Oncol. 2019, 9, 752. [Google Scholar] [CrossRef] [PubMed]
- Su, C.M.; Weng, Y.S.; Kuan, L.Y.; Chen, J.H.; Hsu, F.T. Suppression of PKCδ/NF-κB Signaling and Apoptosis Induction through Extrinsic/Intrinsic Pathways Are Associated Magnolol-Inhibited Tumor Progression in Colorectal Cancer In Vitro and In Vivo. Int. J. Mol. Sci. 2020, 21, 3527. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, K.; Ding, X.; Tang, H.; Xu, Z. Magnolol inhibits growth and induces apoptosis in esophagus cancer KYSE-150 cell lines via the MAP kinase pathway. J. Thorac. Dis. 2019, 11, 3030–3038. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Nagase, H. Magnolol has the ability to induce apoptosis in tumor cells. Biol. Pharm. Bull. 2002, 25, 1546–1549. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, F.; Wang, X.; Wu, X.; Zhang, B.; Zhang, N.; Wu, W.; Wang, Z.; Weng, H.; Liu, S.; et al. Magnolol inhibits growth of gallbladder cancer cells through the p53 pathway. Cancer Sci. 2015, 106, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Rasul, A.; Yu, B.; Khan, M.; Zhang, K.; Iqbal, F.; Ma, T.; Yang, H. Magnolol, a natural compound, induces apoptosis of SGC-7901 human gastric adenocarcinoma cells via the mitochondrial and PI3K/Akt signaling pathways. Int. J. Oncol. 2012, 40, 1153–1161. [Google Scholar] [CrossRef]
- Yueh, P.F.; Lee, Y.H.; Fu, C.Y.; Tung, C.B.; Hsu, F.T.; Lan, K.L. Magnolol Induces the Extrinsic/Intrinsic Apoptosis Pathways and Inhibits STAT3 Signaling-Mediated Invasion of Glioblastoma Cells. Life 2021, 11, 1399. [Google Scholar] [CrossRef]
- Chen, L.C.; Liu, Y.C.; Liang, Y.C.; Ho, Y.S.; Lee, W.S. Magnolol inhibits human glioblastoma cell proliferation through upregulation of p21/Cip1. J. Agric. Food Chem. 2009, 57, 7331–7337. [Google Scholar] [CrossRef]
- Chen, L.C.; Lee, W.S. P27/Kip1 is responsible for magnolol-induced U373 apoptosis in vitro and in vivo. J. Agric. Food Chem. 2013, 61, 2811–2819. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Hueng, D.Y.; Huang, H.Y.; Chen, J.Y.; Chen, Y. Magnolol and honokiol exert a synergistic anti-tumor effect through autophagy and apoptosis in human glioblastomas. Oncotarget 2016, 7, 29116–29130. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Tsao, M.J.; Chiu, C.Y.; Kan, P.C.; Chen, Y. Magnolol Inhibits Human Glioblastoma Cell Migration by Regulating N-Cadherin. J. Neuropathol. Exp. Neurol. 2018, 77, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Hsu, F.T.; Chen, W.L.; Chen, J.H. Induction of Apoptosis, Inhibition of MCL-1, and VEGF-A Expression Are Associated with the Anti-Cancer Efficacy of Magnolol Combined with Regorafenib in Hepatocellular Carcinoma. Cancers 2021, 13, 2066. [Google Scholar] [CrossRef] [PubMed]
- Kuan, L.Y.; Chen, W.L.; Chen, J.H.; Hsu, F.T.; Liu, T.T.; Chen, W.T.; Wang, K.L.; Chen, W.C.; Liu, Y.C.; Wang, W.S. Magnolol Induces Apoptosis and Inhibits ERK-modulated Metastatic Potential in Hepatocellular Carcinoma Cells. In Vivo 2018, 32, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.D.; Sun, X.J.; Yang, W.J.; Li, J.; Yin, J.J. Magnolol exerts anticancer activity in hepatocellular carcinoma cells through regulating endoplasmic reticulum stress-mediated apoptotic signaling. Onco Targets Ther. 2018, 11, 5219–5226. [Google Scholar] [CrossRef]
- Tsai, J.J.; Chen, J.H.; Chen, C.H.; Chung, J.G.; Hsu, F.T. Apoptosis induction and ERK/NF-κB inactivation are associated with magnolol-inhibited tumor progression in hepatocellular carcinoma in vivo. Environ. Toxicol. 2020, 35, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.R.; Chong, I.W.; Chen, Y.H.; Hwang, J.J.; Yin, W.H.; Chen, H.L.; Chou, S.H.; Chiu, C.C.; Liu, P.L. Magnolol induces apoptosis via caspase-independent pathways in non-small cell lung cancer cells. Arch. Pharm. Res. 2014, 37, 548–557. [Google Scholar] [CrossRef]
- Li, H.B.; Yi, X.; Gao, J.M.; Ying, X.X.; Guan, H.Q.; Li, J.C. Magnolol-induced H460 cells death via autophagy but not apoptosis. Arch. Pharm. Res. 2007, 30, 1566–1574. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.U.; Kim, M.H.; Kim, H.M.; Jeong, H.J. Anticancer potential of magnolol for lung cancer treatment. Arch. Pharm. Res. 2011, 34, 625–633. [Google Scholar] [CrossRef]
- Shen, J.; Ma, H.; Zhang, T.; Liu, H.; Yu, L.; Li, G.; Li, H.; Hu, M. Magnolol Inhibits the Growth of Non-Small Cell Lung Cancer via Inhibiting Microtubule Polymerization. Cell Physiol. Biochem. 2017, 42, 1789–1801. [Google Scholar] [CrossRef]
- Ikai, T.; Akao, Y.; Nakagawa, Y.; Ohguchi, K.; Sakai, Y.; Nozawa, Y. Magnolol-induced apoptosis is mediated via the intrinsic pathway with release of AIF from mitochondria in U937 cells. Biol. Pharm. Bull. 2006, 29, 2498–2501. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Li, M.; Jiao, G. Magnolol induces apoptosis via activation of both mitochondrial and death receptor pathways in A375-S2 cells. Arch. Pharm. Res. 2009, 32, 1789–1794. [Google Scholar] [CrossRef] [PubMed]
- Emran, A.A.; Chinna Chowdary, B.R.; Ahmed, F.; Hammerlindl, H.; Huefner, A.; Haass, N.K.; Schuehly, W.; Schaider, H. Magnolol induces cell death through PI3K/Akt-mediated epigenetic modifications boosting treatment of BRAF- and NRAS-mutant melanoma. Cancer Med. 2019, 8, 1186–1196. [Google Scholar] [CrossRef]
- Jin, W.; Wang, X.; Zeng, Y.; Lan, Y.; Wang, X. Magnolol suppressed cell migration and invasion and induced cell apoptosis via inhibition of the NF-κB signaling pathway by upregulating microRNA-129 in multiple myeloma. Neoplasma 2021, 68, 404–415. [Google Scholar] [CrossRef]
- Peng, C.Y.; Yu, C.C.; Huang, C.C.; Liao, Y.W.; Hsieh, P.L.; Chu, P.M.; Yu, C.H.; Lin, S.S. Magnolol inhibits cancer stemness and IL-6/Stat3 signaling in oral carcinomas. J. Formos. Med. Assoc. 2022, 121, 51–57. [Google Scholar] [CrossRef]
- Chen, Y.T.; Lin, C.W.; Su, C.W.; Yang, W.E.; Chuang, C.Y.; Su, S.C.; Hsieh, M.J.; Yang, S.F. Magnolol Triggers Caspase-Mediated Apoptotic Cell Death in Human Oral Cancer Cells through JNK1/2 and p38 Pathways. Biomedicines 2021, 9, 1295. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wen, H.; Li, H. Magnolol induces apoptosis in osteosarcoma cells via G0/G1 phase arrest and p53-mediated mitochondrial pathway. J. Cell Biochem. 2019, 120, 17067–17079. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.C.; Hsu, S.C.; Cheng, Y.T.; Shao, W.S.; Wu, K.; Fang, G.S.; Ou, C.C.; Wang, V. Magnolol down-regulates HER2 gene expression, leading to inhibition of HER2-mediated metastatic potential in ovarian cancer cells. Cancer Lett. 2011, 311, 11–19. [Google Scholar] [CrossRef]
- Chen, S.; Shen, J.; Zhao, J.; Wang, J.; Shan, T.; Li, J.; Xu, M.; Chen, X.; Liu, Y.; Cao, G. Magnolol Suppresses Pancreatic Cancer Development In Vivo and In Vitro via Negatively Regulating TGF-β/Smad Signaling. Front. Oncol. 2020, 10, 597672. [Google Scholar] [CrossRef]
- Lee, D.H.; Szczepanski, M.J.; Lee, Y.J. Magnolol induces apoptosis via inhibiting the EGFR/PI3K/Akt signaling pathway in human prostate cancer cells. J. Cell Biochem. 2009, 106, 1113–1122. [Google Scholar] [CrossRef]
- McKeown, B.T.; Hurta, R.A. Magnolol affects expression of IGF-1 and associated binding proteins in human prostate cancer cells in vitro. Anticancer Res. 2014, 34, 6333–6338. [Google Scholar] [PubMed]
- McKeown, B.T.; McDougall, L.; Catalli, A.; Hurta, R.A. Magnolol causes alterations in the cell cycle in androgen insensitive human prostate cancer cells in vitro by affecting expression of key cell cycle regulatory proteins. Nutr. Cancer 2014, 66, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Park, K.K. Magnolol suppresses metastasis via inhibition of invasion, migration, and matrix metalloproteinase-2/-9 activities in PC-3 human prostate carcinoma cells. Biosci. Biotechnol. Biochem. 2010, 74, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Zhou, S.; Song, J. Induction of apoptosis by magnolol via the mitochondrial pathway and cell cycle arrest in renal carcinoma cells. Biochem. Biophys. Res. Commun. 2019, 508, 1271–1278. [Google Scholar] [CrossRef]
- Woo, S.M.; Min, K.J.; Kwon, T.K. Magnolol Enhances the Therapeutic Effects of TRAIL through DR5 Upregulation and Downregulation of c-FLIP and Mcl-1 Proteins in Cancer Cells. Molecules 2020, 25, 4591. [Google Scholar] [CrossRef] [PubMed]
- Chilampalli, C.; Guillermo, R.; Zhang, X.; Kaushik, R.S.; Young, A.; Zeman, D.; Hildreth, M.B.; Fahmy, H.; Dwivedi, C. Effects of magnolol on UVB-induced skin cancer development in mice and its possible mechanism of action. BMC Cancer 2011, 11, 456. [Google Scholar] [CrossRef]
- Kuo, D.H.; Lai, Y.S.; Lo, C.Y.; Cheng, A.C.; Wu, H.; Pan, M.H. Inhibitory effect of magnolol on TPA-induced skin inflammation and tumor promotion in mice. J. Agric. Food Chem. 2010, 58, 5777–5783. [Google Scholar] [CrossRef]
- Huang, S.H.; Chen, Y.; Tung, P.Y.; Wu, J.C.; Chen, K.H.; Wu, J.M.; Wang, S.M. Mechanisms for the magnolol-induced cell death of CGTH W-2 thyroid carcinoma cells. J. Cell Biochem. 2007, 101, 1011–1022. [Google Scholar] [CrossRef]
- Hassan, B.; Akcakanat, A.; Holder, A.M.; Meric-Bernstam, F. Targeting the PI3-kinase/Akt/mTOR signaling pathway. Surg. Oncol. Clin. North Am. 2013, 22, 641–664. [Google Scholar] [CrossRef]
- Alessi, D.R.; James, S.R.; Downes, C.P.; Holmes, A.B.; Gaffney, P.R.; Reese, C.B.; Cohen, P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Balpha. Curr. Biol. 1997, 7, 261–269. [Google Scholar] [CrossRef]
- Memmott, R.M.; Dennis, P.A. Akt-dependent and -independent mechanisms of mTOR regulation in cancer. Cell Signal. 2009, 21, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Dufour, M.; Dormond-Meuwly, A.; Demartines, N.; Dormond, O. Targeting the Mammalian Target of Rapamycin (mTOR) in Cancer Therapy: Lessons from Past and Future Perspectives. Cancers 2011, 3, 2478–2500. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sung, B. NF-κB in cancer: A matter of life and death. Cancer Discov. 2011, 1, 469–471. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Xia, Y.; Shen, S.; Verma, I.M. NF-κB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef]
- Law, M.E.; Corsino, P.E.; Narayan, S.; Law, B.K. Cyclin-Dependent Kinase Inhibitors as Anticancer Therapeutics. Mol. Pharmacol. 2015, 88, 846–852. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Liu, X.; Kim, C.; Yang, J.; Jemmerson, R.; Wang, X. Induction of apoptotic program in cell-free extracts: Requirement for dATP and cytochrome c. Cell 1996, 86, 147–157. [Google Scholar] [CrossRef]
- Kovarik, J.; Siegrist, C.-A. Immunity in early life. Immunol. Today 1998, 19, 150–152. [Google Scholar] [CrossRef]
- Saelens, X.; Festjens, N.; Vande Walle, L.; van Gurp, M.; van Loo, G.; Vandenabeele, P. Toxic proteins released from mitochondria in cell death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, T.N.; Huysentruyt, L.C. On the origin of cancer metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef]
- Fidler, I.J.; Kripke, M.L. The challenge of targeting metastasis. Cancer Metastasis Rev. 2015, 34, 635–641. [Google Scholar] [CrossRef]
- Singh, D.; Srivastava, S.K.; Chaudhuri, T.K.; Upadhyay, G. Multifaceted role of matrix metalloproteinases (MMPs). Front. Mol. Biosci. 2015, 2, 19. [Google Scholar] [CrossRef]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef]
- Mittal, V. Epithelial Mesenchymal Transition in Tumor Metastasis. Annu. Rev. Pathol. 2018, 13, 395–412. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Invest. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef]
- Höckel, M.; Vaupel, P. Biological consequences of tumor hypoxia. Semin. Oncol. 2001, 28, 36–41. [Google Scholar] [CrossRef]
- Le, Q.T.; Denko, N.C.; Giaccia, A.J. Hypoxic gene expression and metastasis. Cancer Metastasis Rev. 2004, 23, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Wang, L.; Esko, J.; Giordano, F.J.; Huang, Y.; Gerber, H.P.; Ferrara, N.; Johnson, R.S. Loss of HIF-1alpha in endothelial cells disrupts a hypoxia-driven VEGF autocrine loop necessary for tumorigenesis. Cancer Cell 2004, 6, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Kuk, H.; Arnold, C.; Meyer, R.; Hecker, M.; Korff, T. Magnolol inhibits venous remodeling in mice. Sci. Rep. 2017, 7, 17820. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D.; Oh, J.; Park, H.J.; Bae, K.; Lee, S.K. Magnolol inhibits angiogenesis by regulating ROS-mediated apoptosis and the PI3K/AKT/mTOR signaling pathway in mES/EB-derived endothelial-like cells. Int. J. Oncol. 2013, 43, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Jada, S.; Doma, M.R.; Singh, P.P.; Kumar, S.; Malik, F.; Sharma, A.; Khan, I.A.; Qazi, G.N.; Kumar, H.M. Design and synthesis of novel magnolol derivatives as potential antimicrobial and antiproliferative compounds. Eur. J. Med. Chem. 2012, 51, 35–41. [Google Scholar] [CrossRef]
- Kumar, S.; Guru, S.K.; Pathania, A.S.; Kumar, A.; Bhushan, S.; Malik, F. Autophagy triggered by magnolol derivative negatively regulates angiogenesis. Cell Death Dis. 2013, 4, e889. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, A.; Pathania, A.S.; Guru, S.K.; Jada, S.; Sharma, P.R.; Bhushan, S.; Saxena, A.K.; Kumar, H.M.; Malik, F. Tiron and trolox potentiate the autophagic cell death induced by magnolol analog Ery5 by activation of Bax in HL-60 cells. Apoptosis 2013, 18, 605–617. [Google Scholar] [CrossRef]
- Xu, T.; Zheng, Z.; Guo, Y.; Bai, L.P. Semisynthesis of novel magnolol-based Mannich base derivatives that suppress cancer cells via inducing autophagy. Eur. J. Med. Chem. 2020, 205, 112663. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, Y.; Li, D.; Fu, S.; Tang, M.; Wan, L.; Chen, K.; Liu, Z.; Xue, L.; Peng, A.; et al. Discovery and synthesis of novel magnolol derivatives with potent anticancer activity in non-small cell lung cancer. Eur. J. Med. Chem. 2018, 156, 190–205. [Google Scholar] [CrossRef]
- Zhao, M.; Zheng, Y.H.; Zhao, Q.Y.; Zheng, W.; Yang, J.H.; Pei, H.Y.; Liu, L.; Liu, K.J.; Xue, L.L.; Deng, D.X.; et al. Synthesis and evaluation of new compounds bearing 3-(4-aminopiperidin-1-yl)methyl magnolol scaffold as anticancer agents for the treatment of non-small cell lung cancer via targeting autophagy. Eur. J. Med. Chem. 2021, 209, 112922. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tao, C.; Huang, X.; Chen, Z.; Wang, L.; Li, X.; Ma, M.; Wu, Z. CT2-3, a novel magnolol analogue suppresses NSCLC cells through triggering cell cycle arrest and apoptosis. Bioorganic Med. Chem. 2020, 28, 115352. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Chen, J.; Huang, X.; Chen, Z.; Li, X.; Li, Y.; Xu, Y.; Ma, M.; Wu, Z. CT1-3, a novel magnolol-sulforaphane hybrid suppresses tumorigenesis through inducing mitochondria-mediated apoptosis and inhibiting epithelial mesenchymal transition. Eur. J. Med. Chem. 2020, 199. [Google Scholar] [CrossRef]
- Sun, X.-L.; Zhu, M.-L.; Dai, Y.-Q.; Li, H.-M.; Li, B.-H.; Ma, H.; Zhang, C.-H.; Wu, C.-Z. Semi-Synthesis and In Vitro Anti-Cancer Evaluation of Magnolol Derivatives. Molecules 2021, 26, 4302. [Google Scholar] [CrossRef]
- Ding, R.B.; Chen, P.; Rajendran, B.K.; Lyu, X.; Wang, H.; Bao, J.; Zeng, J.; Hao, W.; Sun, H.; Wong, A.H.; et al. Molecular landscape and subtype-specific therapeutic response of nasopharyngeal carcinoma revealed by integrative pharmacogenomics. Nat. Commun. 2021, 12, 3046. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, D.; Yang, G.; Shi, Q.; Feng, F. Comparative pharmacokinetics and brain distribution of magnolol and honokiol after oral administration of Magnolia officinalis cortex extract and its compatibility with other herbal medicines in Zhi-Zi-Hou-Po Decoction to rats. Biomed. Chromatogr. 2016, 30, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.H.; Chou, C.J.; Lee, T.F.; Wang, L.C.H.; Chen, C.F. Pharmacokinetic and Pharmacodynamic Studies of Magnolol after Oral Administration in Rats. Pharm. Pharmacol. Commun. 1996, 2, 191–193. [Google Scholar]
- Lin, Y.; Li, Y.; Zeng, Y.; Tian, B.; Qu, X.; Yuan, Q.; Song, Y. Pharmacology, Toxicity, Bioavailability, and Formulation of Magnolol: An Update. Front. Pharmacol. 2021, 12, 632767. [Google Scholar] [CrossRef]
- Lin, H.L.; Cheng, W.T.; Chen, L.C.; Ho, H.O.; Lin, S.Y.; Hsieh, C.M. Honokiol/Magnolol-Loaded Self-Assembling Lecithin-Based Mixed Polymeric Micelles (lbMPMs) for Improving Solubility to Enhance Oral Bioavailability. Int. J. Nanomed. 2021, 16, 651–665. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chien, Y.C.; Wu, C.H.; Liu, D.M. Magnolol-loaded core-shell hydrogel nanoparticles: Drug release, intracellular uptake, and controlled cytotoxicity for the inhibition of migration of vascular smooth muscle cells. Mol. Pharm. 2011, 8, 2339–2349. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).