
The Effect of Selected Bee Products on Adhesion and Biofilm of Clostridioides difficile Strains Belonging to Different Ribotypes


Abstract
:1. Introduction
2. Results
2.1. MIC and MBC Values of C. difficile with Bee Products
2.2. Effect of Bee Products on Adhesive Properties of C. difficile
2.3. Effect of Bee Products on C. difficile Biofilm Formation
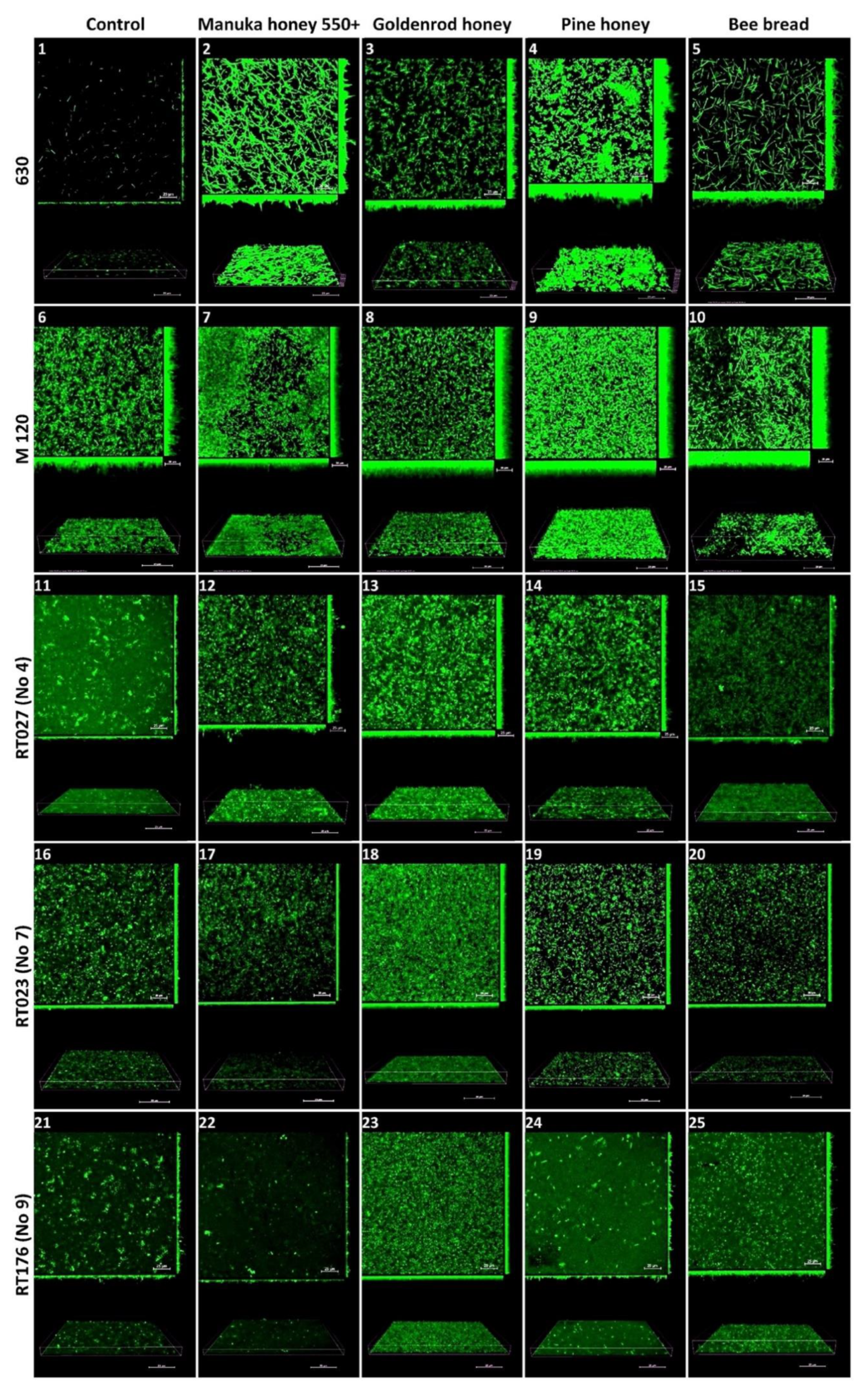
2.4. Confocal Laser Scanning Microscopy
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Bee Products
4.3. Determination of the Minimum Inhibitory and Minimum Bactericidal Concentrations of Bee Products for C. difficile Strains Belonging to Different PCR-RTs
4.4. Cell Cultures
4.5. The Effect of Bee Products on the Adhesion of the Tested C. difficile Strains to Cell Lines
4.6. Testing the Formation of Biofilm of C. difficile Strains under the Influence of Bee Products
4.7. Confocal Laser Scanning Microscopy
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Van Prehn, J.; Reigadas, E.; Vogelzang, E.H.; Bouza, E.; Hristea, A.; Guery, B.; Krutova, M.; Noren, T.; Allerberger, F.; Coia, J.E.; et al. European Society of Clinical Microbiology and Infectious Diseases: 2021 update on the treatment guidance document for Clostridioides difficile infection in adults. Clin. Microbiol. Infect. 2021, 27, S1–S21. [Google Scholar] [CrossRef] [PubMed]
- Ajami, N.J.; Cope, J.L.; Wong, M.C.; Petrosino, J.F.; Chesnel, L. Impact of Oral Fidaxomicin Administration on the Intestinal Microbiota and Susceptibility to Clostridium difficile Colonization in Mice. Antimicrob. Agents Chemother. 2018, 62, e02112-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dapa, T.; Unnikrishnan, M. Biofilm formation by Clostridium difficile. Gut Microbes 2013, 4, 397–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, L.R.; Cheng, J.K.J.; Unnikrishnan, M. Clostridioides difficile biofilms: A mechanism of persistence in the gut? PLoS Pathog. 2021, 17, e1009348. [Google Scholar] [CrossRef]
- Semenyuk, E.G.; Laning, M.L.; Foley, J.; Johnston, P.F.; Knight, K.L.; Gerding, D.N.; Driks, A. Spore Formation and Toxin Production in Clostridium difficile Biofilms. PLoS ONE 2014, 9, e87757. [Google Scholar] [CrossRef] [Green Version]
- Pepin, J.; Valiquette, L.; Alary, M.E.; Villemure, P.; Pelletier, A.; Forget, K.; Pepin, K.; Chouinard, D. Clostridium difficile-associated diarrhea in a region of Quebec from 1991 to 2003: A changing pattern of disease severity. Can. Med. Assoc. J. 2004, 171, 466–472. [Google Scholar] [CrossRef] [Green Version]
- Loo, V.G.; Poirier, L.; Miller, M.A.; Oughton, M.; Libman, M.D.; Michaud, S.; Bourgault, A.M.; Nguyen, T.; Frenette, C.; Kelly, M.; et al. A predominantly clonal multi-institutional outbreak of Clostridium difficile-associated diarrhea with high morbidity and mortality. N. Engl. J. Med. 2005, 353, 2442–2449. [Google Scholar] [CrossRef] [Green Version]
- McDonald, L.C.; Killgore, G.E.; Thompson, A.; Owens, R.C.; Kazakova, S.V.; Sambol, S.P.; Johnson, S.; Gerding, D.N. An epidemic, toxin gene-variant strain of Clostridium difficile. N. Engl. J. Med. 2005, 353, 2433–2441. [Google Scholar] [CrossRef] [Green Version]
- Brazier, J.; Raybould, R.; Patel, B.; Duckworth, G.; Pearson, A.; Charlett, A.; Duerden, B. Distribution and antimicrobial susceptibility patterns of Clostridium difficile PCR ribotypes in English hospitals, 2007–2008. Eurosurveillance 2008, 13, 19000. [Google Scholar] [CrossRef] [Green Version]
- Indra, A.; Huhulescu, S.; Schneeweis, M.; Hasenberger, P.; Kernbichler, S.; Fiedler, A.; Wewalka, G.; Allerberger, F.; Kuijper, E.J. Characterization of Clostridium difficile isolates using capillary gel electrophoresis-based PCR ribotyping. J. Med. Microbiol. 2008, 57, 1377–1382. [Google Scholar] [CrossRef]
- Valiente, E.; Cairns, M.D.; Wren, B.W. The Clostridium difficile PCR ribotype 027 lineage: A pathogen on the move. Clin. Microbiol. Infect. 2014, 20, 396–404. [Google Scholar] [CrossRef] [Green Version]
- Pituch, H.; Bakker, D.; Kuijper, E.; Obuch-Woszczatynski, P.; Wultanska, D.; Nurzynska, G.; Bielec, A.; Bar-Andziak, E.; Luczak, M. First isolation of Clostridium difficile PCR-ribotype 027/toxinotype III in Poland. Pol. J. Microbiol. 2008, 57, 267–268. [Google Scholar]
- Lachowicz, D.; Szulencka, G.; Obuch-Woszczatynski, P.; van Belkum, A.; Pituch, H. First Polish outbreak of Clostridium difficile ribotype 027 infections among dialysis patients. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Hubert, B.; Loo, V.G.; Bourgault, A.M.; Poirier, L.; Dascal, A.; Fortin, E.; Dionne, M.; Lorange, M. A portrait of the geographic dissemination of the Clostridium difficile North American pulsed-field type 1 strain and the epidemiology of C-difficile-associated disease in Quebec. Clin. Infect. Dis. 2007, 44, 238–244. [Google Scholar] [CrossRef] [Green Version]
- Mooney, H. Annual incidence of MRSA falls in England, but C difficile continues to rise, report shows. Br. Med. J. 2007, 335, 958. [Google Scholar] [CrossRef] [Green Version]
- Redelings, M.D.; Sorvillo, F.; Mascola, L. Increase in Clostridium difficile-related mortality rates, United States, 1999–2004. Emerg. Infect. Dis. 2007, 13, 1417–1419. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Miyajima, F.; Roberts, P.; Ellison, L.; Pickard, D.J.; Martin, M.J.; Connor, T.R.; Harris, S.R.; Fairley, D.; Bamford, K.B.; et al. Emergence and global spread of epidemic healthcare-associated Clostridium difficile. Nat. Genet. 2013, 45, U109–U155. [Google Scholar] [CrossRef]
- Arvand, M.; Bettge-Weller, G. Clostridium difficile ribotype 027 is not evenly distributed in Hesse, Germany. Anaerobe 2016, 40, 1–4. [Google Scholar] [CrossRef]
- Arvand, M.; Vollandt, D.; Bettge-Weller, G.; Harmanus, C.; Kuijper, E. Increased incidence of Clostridium difficile PCR ribotype 027 in Hesse, Germany, 2011 to 2013. Eurosurveillance 2014, 19, 20732. [Google Scholar] [CrossRef] [Green Version]
- Pituch, H.; Obuch-Woszczatynski, P.; Lachowicz, D.; Wultanska, D.; Karpinski, P.; Mlynarczyk, G.; van Dorp, S.M.; Kuijper, E.J.; Polish Clostridium Difficile, S. Hospital-based Clostridium difficile infection surveillance reveals high proportions of PCR ribotypes 027 and 176 in different areas of Poland, 2011 to 2013. Eurosurveillance 2015, 20, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Obuch-Woszczatynski, P.; Lachowicz, D.; Schneider, A.; Mol, A.; Pawlowska, J.; Ozdzenska-Milke, E.; Pruszczyk, P.; Wultanska, D.; Mlynarczyk, G.; Harmanus, C.; et al. Occurrence of Clostridium difficile PCR-ribotype 027 and it’s closely related PCR-ribotype 176 in hospitals in Poland in 2008–2010. Anaerobe 2014, 28, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Shaw, H.A.; Preston, M.D.; Vendrik, K.E.W.; Cairns, M.D.; Browne, H.P.; Stabler, R.A.; Crobach, M.J.T.; Corver, J.; Pituch, H.; Ingebretsen, A.; et al. The recent emergence of a highly related virulent Clostridium difficile clade with unique characteristics. Clin. Microbiol. Infect. 2020, 26, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Boateng, J.; Diunase, K.N. Comparing the Antibacterial and Functional Properties of Cameroonian and Manuka Honeys for Potential Wound Healing-Have We Come Full Cycle in Dealing with Antibiotic Resistance? Molecules 2015, 20, 16068–16084. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, B.; Lazim, N.M.; Salim, R. The effectiveness of Tualang honey in reducing post-tonsillectomy pain. Turk. J. Ear Nose Throat 2015, 25, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnwath, R.; Graham, E.M.; Reynolds, K.; Pollock, P.J. The antimicrobial activity of honey against common equine wound bacterial isolates. Vet. J. 2014, 199, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Hammond, E.N.; Donkor, E.S. Antibacterial effect of Manuka honey on Clostridium difficile. BMC Res. Notes 2013, 6, 188. [Google Scholar] [CrossRef] [Green Version]
- Hawley, P.; Hovan, A.; McGahan, C.E.; Saunders, D. A randomized placebo-controlled trial of manuka honey for radiation-induced oral mucositis. Support. Care Cancer 2014, 22, 751–761. [Google Scholar] [CrossRef]
- Piotrowski, M.; Karpinski, P.; Pituch, H.; van Belkum, A.; Obuch-Woszczatynski, P. Antimicrobial effects of Manuka honey on in vitro biofilm formation by Clostridium difficile. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1661–1664. [Google Scholar] [CrossRef]
- Siavash, M.; Shokri, S.; Haghighi, S.; Shahtalebi, M.A.; Farajzadehgan, Z. The efficacy of topical royal jelly on healing of diabetic foot ulcers: A double-blind placebo-controlled clinical trial. Int. Wound J. 2015, 12, 137–142. [Google Scholar] [CrossRef]
- Wallace, A.; Eady, S.; Miles, M.; Martin, H.; McLachlan, A.; Rodier, M.; Willis, J.; Scott, R.; Sutherland, J. Demonstrating the safety of manuka honey UMF (R) 20+in a human clinical trial with healthy individuals. Br. J. Nutr. 2010, 103, 1023–1028. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, J.M.; Cavanagh, H.M.A. Antibacterial activity of 13 honeys against Escherichia coli and Pseudomonas aeruginosa. J. Med. Food 2005, 8, 100–103. [Google Scholar] [CrossRef]
- Van den Berg, A.; Van den Worm, E.; Quarles van Ufford, H.; Halkes, S.; Hoekstra, M.; Beukelman, C. An in vitro examination of the antioxidant and anti-inflammatory properties of buckwheat honey. J. Wound Care 2008, 17, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Umeda, N.; Maeda, A.; Matsumoto, D.; Kitamoto, N.; Kikuzaki, H. Identification of a Novel Glycoside, Leptosin, as a Chemical Marker of Manuka Honey. J. Agric. Food Chem. 2012, 60, 3418–3423. [Google Scholar] [CrossRef]
- Kwakman, P.H.S.; Velde, A.A.T.; de Boer, L.; Vandenbroucke-Grauls, C.; Zaat, S.A.J. Two Major Medicinal Honeys Have Different Mechanisms of Bactericidal Activity. PLoS ONE 2011, 6, e17709. [Google Scholar] [CrossRef] [Green Version]
- Mavric, E.; Wittmann, S.; Barth, G.; Henle, T. Identification and quantification of methylglyoxal as the dominant antibacterial constituent of Manuka (Leptospermum scoparium) honeys from New Zealand. Mol. Nutr. Food Res. 2008, 52, 483–489. [Google Scholar] [CrossRef]
- Molan, P.C. The antibacterial activity of honey.1. the nature of the antibacterial activity. Bee World 1992, 73, 5–28. [Google Scholar] [CrossRef]
- Al-Waili, N.; Al Ghamdi, A.; Ansari, M.J.; Al-Attal, Y.; Al-Mubarak, A.; Salom, K. Differences in Composition of Honey Samples and Their Impact on the Antimicrobial Activities against Drug Multiresistant Bacteria and Pathogenic Fungi. Arch. Med. Res. 2013, 44, 307–316. [Google Scholar] [CrossRef]
- Brudzynski, K.; Miotto, D. The recognition of high molecular weight melanoidins as the main components responsible for radical-scavenging capacity of unheated and heat-treated Canadian honeys. Food Chem. 2011, 125, 570–575. [Google Scholar] [CrossRef]
- Kolayli, S.; Sahin, H.; Can, Z.; Yildiz, O.; Sahin, K. Honey shows potent inhibitory activity against the bovine testes hyaluronidase. J. Enzym. Inhib. Med. Chem. 2016, 31, 599–602. [Google Scholar] [CrossRef]
- Kolayli, S.; Can, Z.; Yildiz, O.; Sahin, H.; Karaoglu, S.A. A comparative study of the antihyaluronidase, antiurease, antioxidant, antimicrobial and physicochemical properties of different unifloral degrees of chestnut (Castanea sativa Mill.) honeys. J. Enzym. Inhib. Med. Chem. 2016, 31, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Ramanauskiene, K.; Stelmakiene, A.; Briedis, V.; Ivanauskas, L.; Jakstas, V. The quantitative analysis of biologically active compounds in Lithuanian honey. Food Chem. 2012, 132, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, J.; Pietrzak, M.; Pomianowski, J.; Wieczorek, Z. Honey as a source of bioactive compounds. Pol. J. Nat. Sci. 2014, 29, 275–285. [Google Scholar]
- Aggad, H.; Guemour, D. Honey antibacterial activity. Med. Arom. Plants 2014, 3, 152. [Google Scholar] [CrossRef] [Green Version]
- Dzugan, M.; Tomczyk, M.; Sowa, P.; Grabek-Lejko, D. Antioxidant Activity as Biomarker of Honey Variety. Molecules 2018, 23, 2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbanna, K.; Attalla, K.; Elbadry, M.; Abdeltawab, A.; Gamal-Eldin, H.; Ramadan, M.F. Impact of floral sources and processing on the antimicrobial activities of different unifloral honeys. Asian Pac. J. Trop. Dis. 2014, 4, 194–200. [Google Scholar] [CrossRef]
- Cornara, L.; Biagi, M.; Xiao, J.B.; Burlando, B. Therapeutic Properties of Bioactive Compounds from Different Honeybee Products. Front. Pharmacol. 2017, 8, 412. [Google Scholar] [CrossRef] [Green Version]
- Majtan, J.; Kumar, P.; Majtan, T.; Walls, A.F.; Klaudiny, J. Effect of honey and its major royal jelly protein 1 on cytokine and MMP-9 mRNA transcripts in human keratinocytes. Exp. Dermatol. 2010, 19, E73–E79. [Google Scholar] [CrossRef]
- Henriques, A.F.; Jenkins, R.E.; Burton, N.F.; Cooper, R.A. The effect of manuka honey on the structure of Pseudomonas aeruginosa. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, R.; Burton, N.; Cooper, R. Manuka honey inhibits cell division in methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2011, 66, 2536–2542. [Google Scholar] [CrossRef] [Green Version]
- Al-Nahari, A.A.M.; Almasaudi, S.B.; El-Ghany, E.; Barbour, E.; Al Jaouni, S.K.; Harakeh, S. Antimicrobial activities of Saudi honey against Pseudomonas aeruginosa. Saudi J. Biol. Sci. 2015, 22, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Kus, P.M.; Szweda, P.; Jerkovic, I.; Tuberoso, C.I.G. Activity of Polish unifloral honeys against pathogenic bacteria and its correlation with colour, phenolic content, antioxidant capacity and other parameters. Lett. Appl. Microbiol. 2016, 62, 269–276. [Google Scholar] [CrossRef]
- Cooper, R.A.; Wigley, P.; Burton, N.F. Susceptibility of multiresistant strains of Burkholderia cepacia to honey. Lett. Appl. Microbiol. 2000, 31, 20–24. [Google Scholar] [CrossRef]
- Cooper, R.A.; Halas, E.; Molan, P.C. The efficacy of honey in inhibiting strains of Pseudomonas aeruginosa from infected burns. J. Burn Care Rehabil. 2002, 23, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Lusby, P.E.; Coombes, A.L.; Wilkinson, J.M. Bactericidal activity of different honeys against pathogenic bacteria. Arch. Med. Res. 2005, 36, 464–467. [Google Scholar] [CrossRef]
- Taormina, P.J.; Niemira, B.A.; Beuchat, L.R. Inhibitory activity of honey against foodborne pathogens as influenced by the presence of hydrogen peroxide and level of antioxidant power. Int. J. Food Microbiol. 2001, 69, 217–225. [Google Scholar] [CrossRef]
- Maddocks, S.E.; Lopez, M.S.; Rowlands, R.S.; Cooper, R.A. Manuka honey inhibits the development of Streptococcus pyogenes biofilms and causes reduced expression of two fibronectin binding proteins. Microbiol. Immunol. 2012, 158, 781–790. [Google Scholar] [CrossRef] [Green Version]
- Brudzynski, K.; Lannigan, R. Mechanism of honey bacteriostatic action against MRSA and VRE involves hydroxyl radicals generated from honey’s hydrogen peroxide. Front. Microbiol. 2012, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Adams, C.J.; Boult, C.H.; Deadman, B.J.; Farr, J.M.; Grainger, M.N.C.; Manley-Harris, M.; Snow, M.J. Isolation by HPLC and characterisation of the bioactive fraction of New Zealand manuka (Leptospermum scoparium) honey. Carbohydr. Res. 2008, 343, 651–659. [Google Scholar] [CrossRef]
- Brudzynski, K.; Sjaarda, C. Antibacterial Compounds of Canadian Honeys Target Bacterial Cell Wall Inducing Phenotype Changes, Growth Inhibition and Cell Lysis That Resemble Action of beta-Lactam Antibiotics. PLoS ONE 2014, 9, e106967. [Google Scholar] [CrossRef] [Green Version]
- Mizzi, L.; Maniscalco, D.; Gaspari, S.; Chatzitzika, C.; Gatt, R.; Valdramidis, V.P. Assessing the individual microbial inhibitory capacity of different sugars against pathogens commonly found in food systems. Lett. Appl. Microbiol. 2020, 71, 251–258. [Google Scholar] [CrossRef]
- Kafantaris, I.; Tsadila, C.; Nikolaidis, M.; Tsavea, E.; Dimitriou, T.G.; Iliopoulos, I.; Amoutzias, G.D.; Mossialos, D. Transcriptomic Analysis of Pseudomonas aeruginosa Response to Pine Honey via RNA Sequencing Indicates Multiple Mechanisms of Antibacterial Activity. Foods 2021, 10, 936. [Google Scholar] [CrossRef] [PubMed]
- Tsavea, E.; Vardaka, F.-P.; Savvidaki, E.; Kellil, A.; Kanelis, D.; Bucekova, M.; Grigorakis, S.; Godocikova, J.; Gotsiou, P.; Dimou, M.; et al. Physicochemical Characterization and Biological Properties of Pine Honey Produced across Greece. Foods 2022, 11, 943. [Google Scholar] [CrossRef] [PubMed]
- Wultańska, D.; Piotrowski, M.; Pituch, H. The effect of berberine chloride and/or its combination with vancomycin on the growth, biofilm formation, and motility of Clostridioides difficile. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1391–1399. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Strain No. | RT | MIC% of Manuka Honey 550+ | MBC% of Manuka Honey 550+ | MIC% of Goldenrod Honey | MBC% of Goldenrod Honey | MIC% of Pine Honey | MBC% of Pine Honey | MIC (mg/L) of Bee Bread | MBC (mg/L) of Bee Bread |
|---|---|---|---|---|---|---|---|---|---|
| 012 | 6.25 | 6.25 | 50 | ≥100 | 25 | ≥100 | 100 | ≥200 |
| 001 | 6.25 | 6.25 | 50 | ≥100 | 25 | ≥100 | 50 | ≥200 |
| 078 | 1.56 | 3.125 | 12.5 | ≥100 | 25 | ≥100 | 100 | ≥200 |
| 027 | 6.25 | 6.25 | 12.5 | 50 | 3.125 | 12.5 | 50 | ≥200 |
| 027 | 6.25 | 6.25 | 25 | ≥100 | 6.25 | ≥100 | 50 | ≥200 |
| 023 | 6.25 | 6.25 | 25 | ≥100 | 12.5 | ≥100 | 25 | ≥200 |
| 023 | 6.25 | 6.25 | 50 | ≥100 | 25 | ≥100 | 25 | ≥200 |
| 176 | 1.56 | 3.125 | 1.56 | 50 | 1.56 | 50 | 50 | ≥200 |
| 176 | 3.125 | 6.25 | 3.125 | 25 | 1.56 | 25 | 100 | ≥200 |
| 176 | 3.125 | 6.25 | 6.25 | 25 | 25 | 50 | 25 | ≥200 |
| 176 | 3.125 | 6.25 | 12.5 | 25 | 25 | 50 | 25 | ≥200 |
| 176 | 6.25 | 12.5 | 12.5 | 25 | 25 | 50 | 25 | ≥200 |
| Strain | CTR | MAN 100% | MAN 50% | MAN 25% | MAN 12.5% | MAN 6.25% | MAN 3.125% | MAN 1.56% |
|---|---|---|---|---|---|---|---|---|
| No 1 (630) | 0.43 ± 0.02 | 0.10 ± 0.01 * | 0.10 ± 0.02 * | 0.11 ± 0.01 * | 0.12 ± 0.01 * | 0.17 ± 0.01 * | 0.50 ± 0.01 * | 0.51 ± 0.01 * |
| No 2 (ATCC 9689) | 0.31 ± 0.02 | 0.10 ± 0.04 * | 0.10 ± 0.02 * | 0.15 ± 0.02 * | 0.16 ± 0 * | 0.16 ± 0.01 * | 0.41 ± 0.06 * | 0.62 ± 0.05 * |
| No 3 (M 120) | 0.40 ± 0.02 | 0.14 ± 0.04 * | 0.21 ± 0.02 * | 0.14 ± 0.02 * | 0.21 ± 0.02 * | 0.23 ± 0.01 * | 0.70 ± 0.01 * | 0.55 ± 0.01 * |
| No 4 (1468/12) RT027 | 0.24 ± 0.02 | 0.20 ± 0.02 | 0.13 ± 0.02 | 0.16 ± 0.04 | 0.17 ± 0.07 | 0.21 ± 0.03 | 0.29 ± 0.07 | 0.29 ± 0.01 |
| No 5 (2292) RT027 | 0.23 ± 0.03 | 0.10 ± 0 * | 0.11 ± 0.02 * | 0.11 ± 0.02 * | 0.14 ± 0.02 * | 0.12 ± 0.02 * | 0.23 ± 0.07 | 0.33 ± 0.01 * |
| No 6 (2333/06) RT023 | 0.22 ± 0.02 | 0.09 ± 0.01 * | 0.10 ± 0.03 | 0.11 ± 0.01 * | 0.11 ± 0.02 | 0.15 ± 0.02 * | 0.16 ± 0.04 | 0.25 ± 0.07 |
| No 7 (25694/12) RT023 | 0.24 ± 0.03 | 0.09 ± 0.02 * | 0.10 ± 0 * | 0.11 ± 0.02 * | 0.15 ± 0.02 * | 0.18 ± 0.02 * | 0.34 ± 0.03 | 0.33 ± 0.02 |
| No 8 (5291/12) RT176 | 0.18 ± 0.04 | 0.09 ± 0.01 * | 0.11 ± 0 * | 0.17 ± 0.03 * | 0.14 ± 0.02 | 0.16 ± 0.03 | 0.19 ± 0.3 | 0.20 ± 0.05 |
| No 9 (2628/12) RT176 | 0.23 ± 0.06 | 0.09 ± 0.02 | 0.13 ± 0.03 | 0.11 ± 0.01 | 0.17 ± 0.03 | 0.13 ± 0.02 | 0.20 ± 0 | 0.29 ± 0.05 |
| No 10 (536/12) RT176 | 0.21 ± 0.01 | 0.10 ± 0.01 * | 0.12 ± 0.01 * | 0.10 ± 0.02 * | 0.11 ± 0.02 * | 0.38 ± 0.02 * | 0.37 ± 0.03 * | 0.38 ± 0.03 * |
| No 11 (1974/12) RT176 | 0.12 ± 0.01 | 0.10 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.11 ± 0.01 |
| No 12 (3136/12) RT176 | 0.14 ± 0.02 | 0.10 ± 0.01 * | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.17 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.10 ± 0.01 |
| Strain | CTR | GOL 100% | GOL 50% | GOL 25% | GOL 12.5% | GOL 6.25% | GOL 3.125% | GOL 1.56% |
|---|---|---|---|---|---|---|---|---|
| No 1 (630) | 0.43 ± 0.04 | 0.11 ± 0.01 * | 0.21 ± 0.06 * | 0.33 ± 0.1 | 0.39 ± 0.14 | 0.43 ± 0.07 | 0.43 ± 0.03 | 0.45 ± 0.03 |
| No 2 (ATCC 9689) | 0.28 ± 0.02 | 0.09 ± 0.02 * | 0.10 ± 0.02 * | 0.17 ± 0.02 | 0.27 ± 0.06 | 0.39 ± 0.06 | 0.41 ± 0.06 | 0.53 ± 0.1 * |
| No 3 (M 120) | 0.43 ± 0.02 | 0.09 ± 0.02 * | 0.16 ± 0.02 * | 0.26 ± 0.04 * | 0.34 ± 0.05 | 0.54 ± 0.08 | 0.49 ± 0.04 | 0.50 ± 0.07 |
| No 4 (1468/12) RT027 | 0.33 ± 0.12 | 0.08 ± 0.02 * | 0.14 ± 0.02 * | 0.15 ± 0.02 * | 0.21 ± 0.02 | 0.21 ± 0.02 | 0.31 ± 0.02 | 0.25 ± 0.09 |
| No 5 (2292) RT027 | 0.20 ± 0.02 | 0.08 ± 0.02 * | 0.09 ± 0.02 * | 0.14 ± 0.03 | 0.21 ± 0.02 | 0.27 ± 0.04 | 0.33 ± 0.02 * | 0.27 ± 0.02 * |
| No 6 (2333/06) RT023 | 0.23 ± 0.01 | 0.09 ± 0.03 | 0.08 ± 0 | 0.24 ± 0.04 | 0.23 ± 0.06 | 0.38 ± 0.05 | 0.40 ± 0.12 | 0.30 ± 0.03 |
| No 7 (25694/12) RT023 | 0.16 ± 0.05 | 0.08 ± 0 | 0.07 ± 0 | 0.17 ± 0 | 0.27 ± 0.06 | 0.34 ± 0.13 * | 0.23 ± 0.15 | 0.22 ± 0.01 |
| No 8 (5291/12) RT176 | 0.16 ± 0.03 | 0.06 ± 0.01 * | 0.10 ± 0.02 * | 0.10 ± 0.05 * | 0.09 ± 0.01 * | 0.18 ± 0.01 | 0.18 ± 0.01 | 0.18 ± 0.01 |
| No 9 (2628/12) RT176 | 0.18 ± 0.04 | 0.10 ± 0 * | 0.10 ± 0 * | 0.12 ± 0.01 | 0.12 ± 0.04 * | 0.17 ± 0.01 | 0.17 ± 0.01 | 0.18 ± 0.01 |
| No 10 (536/12) RT176 | 0.21 ± 0.01 | 0.05 ± 0.01 * | 0.18 ± 0.02 | 0.33 ± 0.12 | 0.41 ± 0.02 * | 0.40 ± 0.02 * | 0.41 ± 0.04 * | 0.40 ± 0.05 * |
| No 11 (1974/12) RT176 | 0.12 ± 0.01 | 0.10 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.11 ± 0.01 |
| No 12 (3136/12) RT176 | 0.15 ± 0.02 | 0.10 ± 0.01 * | 0.10 ± 0.01 * | 0.12 ± 0.01 * | 0.12 ± 0.02 | 0.12 ± 0.01 * | 0.11 ± 0.01 * | 0.13 ± 0.01 |
| Strain | CTR | PIN 100% | PIN 50% | PIN 25% | PIN 12.5% | PIN 6.25% | PIN 3.125% | PIN 1.56% |
|---|---|---|---|---|---|---|---|---|
| No 1 (630) | 0.24 ± 0.02 | 0.11 ± 0.01 * | 0.12 ± 0.06 * | 0.16 ± 0.02 * | 0.18 ± 0.03 | 0.22 ± 0.02 | 0.21 ± 0.01 | 0.39 ± 0.03 * |
| No 2 (ATCC 9689) | 0.38 ± 0.02 | 0.09 ± 0.01 * | 0.10 ± 0.02 * | 0.17 ± 0.02 * | 0.17 ± 0.02 * | 0.18 ± 0 | 0.24 ± 0.16 | 0.28 ± 0.18 |
| No 3 (M 120) | 0.38 ± 0.01 | 0.09 ± 0.02 * | 0.11 ± 0.02 * | 0.14 ± 0.01 * | 0.19 ± 0.01 * | 0.30 ± 0.12 | 0.33 ± 0.14 | 0.42 ± 0.07 |
| No 4 (1468/12) RT027 | 0.11 ± 0.01 | 0.07 ± 0.01 * | 0.07 ± 0.01 * | 0.08 ± 0.01 * | 0.08 ± 0.01 * | 0.08 ± 0.01 * | 0.08 ± 0.01 * | 0.08 ± 0.01 * |
| No 5 (2292) RT027 | 0.12 ± 0.02 | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.17 ± 0.01 | 0.25 ± 0.12 * | 0.23 ± 0.02 * |
| No 6 (2333/06) RT023 | 0.1 ± 0.01 | 0.07 ± 0.01 * | 0.09 ± 0 | 0.09 ± 0.01 | 0.1 ± 0.01 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.09 ± 0.01 |
| No 7 (25694/12) RT023 | 0.11 ± 0.01 | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.09 ± 0.02 | 0.17 ± 0.03 | 0.30 ± 0.16 * | 0.2 ± 0.13 | 0.21 ± 0.13 |
| No 8 (5291/12) RT176 | 0.11 ± 0.01 | 0.11 ± 0.02 | 0.09 ± 0.01 | 0.12 ± 0.02 | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 |
| No 9 (2628/12) RT176 | 0.12 ± 0.01 | 0.10 ± 0.01 | 0.10 ± 0 | 0.12 ± 0 | 0.12 ± 0.01 | 0.13 ± 0.02 | 0,18 ± 0.01 * | 0,17 ± 0.02 * |
| No 10 (536/12) RT176 | 0.18 ± 0.02 | 0.10 ± 0.01 | 0.18 ± 0.02 | 0.38 ± 0.12 * | 0.41 ± 0.12 * | 0.36 ± 0.07 * | 0.39 ± 0.06 * | 0.35 ± 0.05 |
| No 11 (1974/12) RT176 | 0.11 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.13 ± 0.01 * | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 |
| No 12 (3136/12) RT176 | 0.11 ± 0.02 | 0.1 ± 0.02 | 0.1 ± 0.01 | 0.07 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.02 | 0.1 ± 0.02 |
| Strain | CTR | BEB 200% | BEB 100% | BEB 50% | BEB 25% | BEB 12.5% | BEB 6.25% | BEB 3.125% | BEB 1.56% |
|---|---|---|---|---|---|---|---|---|---|
| No 1 (630) | 0.11 ± 0 | 0.17 ± 0.01 * | 0.18 ± 0.02 * | 0.11 ± 0.01 | 0.15 ± 0.01 | 0.10 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 |
| No 2 (ATCC 9689) | 0.11 ± 0.01 | 0.18 ± 0.01 * | 0.19 ± 0.01 * | 0.17 ± 0.02 * | 0.11 ± 0.02 | 0.11 ± 0 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 |
| No 3 (M 120) | 0.31 ± 0.11 | 0.15 ± 0.01 * | 0.19 ± 0.01 | 0.12 ± 0.01 * | 0.45 ± 0.07 | 0.31 ± 0.01 | 0.33 ± 0.03 | 0.32 ± 0.03 | 0.32 ± 0.03 |
| No 4 (1468/12) RT027 | 0.11 ± 0.01 | 0.13 ± 0.01 | 0.16 ± 0.01 | 0.18 ± 0.01 | 0.15 ± 0.01 | 0.15 ± 0.01 | 0.17 ± 0.01 | 0.18 ± 0.01 | 0.18 ± 0.01 |
| No 5 (2292) RT027 | 0.15 ± 0.02 | 0.13 ± 0.01 | 0.08 ± 0.01 | 0.07 ± 0.01 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.17 ± 0.01 | 0.25 ± 0.12 | 0.23 ± 0.02 |
| No 6 (2333/06) RT023 | 0.25 ± 0 | 0.1 ± 0.01 * | 0.17 ± 0.1 | 0.21 ± 0.12 | 0.2 ± 0.1 | 0.17 ± 0.07 | 0.3 ± 0.04 | 0.32 ± 0.02 | 0.21 ± 0.09 |
| No 7 (25694/12) RT023 | 0.18 ± 0.01 | 0.16 ± 0.01 | 0.18 ± 0.01 | 0.18 ± 0.01 | 0.17 ± 0.05 | 0.16 ± 0.03 | 0.18 ± 0.01 | 0.17 ± 0.01 | 0.18 ± 0.04 |
| No 8 (5291/12) RT176 | 0.11 ± 0.01 | 0.15 ± 0.01 | 0.16 ± 0.01 * | 0.14 ± 0.03 | 0.13 ± 0.01 | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 |
| No 9 (2628/12) RT176 | 0.14 ± 0.01 | 0.13 ± 0.01 | 0.18 ± 0.03 * | 0.10 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.13 ± 0.02 | 0.13 ± 0.01 | 0.13 ± 0.01 |
| No 10 (536/12) RT176 | 0.14 ± 0.01 | 0.13 ± 0.01 | 0.14 ± 0.03 | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.14 ± 0.01 | 0.14 ± 0.02 | 0.14 ± 0.01 | 0.13 ± 0.01 |
| No 11 (1974/12) RT176 | 0.14 ± 0.01 | 0.15 ± 0.01 | 0.14 ± 0.03 | 0.14 ± 0.01 | 0.14 ± 0.03 | 0.09 ± 0.01 | 0.14 ± 0.02 | 0.14 ± 0.01 | 0.13 ± 0.01 |
| No 12 (3136/12) RT176 | 0.14 ± 0.03 | 0.14 ± 0.01 | 0.14 ± 0.03 | 0.12 ± 0 * | 0.14 ± 0.03 | 0.14 ± 0.01 | 0.14 ± 0.02 | 0.14 ± 0.01 | 0.13 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wultańska, D.; Paterczyk, B.; Nowakowska, J.; Pituch, H. The Effect of Selected Bee Products on Adhesion and Biofilm of Clostridioides difficile Strains Belonging to Different Ribotypes. Molecules 2022, 27, 7385. https://doi.org/10.3390/molecules27217385
Wultańska D, Paterczyk B, Nowakowska J, Pituch H. The Effect of Selected Bee Products on Adhesion and Biofilm of Clostridioides difficile Strains Belonging to Different Ribotypes. Molecules. 2022; 27(21):7385. https://doi.org/10.3390/molecules27217385
Chicago/Turabian StyleWultańska, Dorota, Bohdan Paterczyk, Julita Nowakowska, and Hanna Pituch. 2022. "The Effect of Selected Bee Products on Adhesion and Biofilm of Clostridioides difficile Strains Belonging to Different Ribotypes" Molecules 27, no. 21: 7385. https://doi.org/10.3390/molecules27217385