
Marine Antimicrobial Peptides-Based Strategies for Tackling Bacterial Biofilm and Biofouling Challenges
 , and
, and
Abstract
:1. Introduction
2. Biofilm Formation
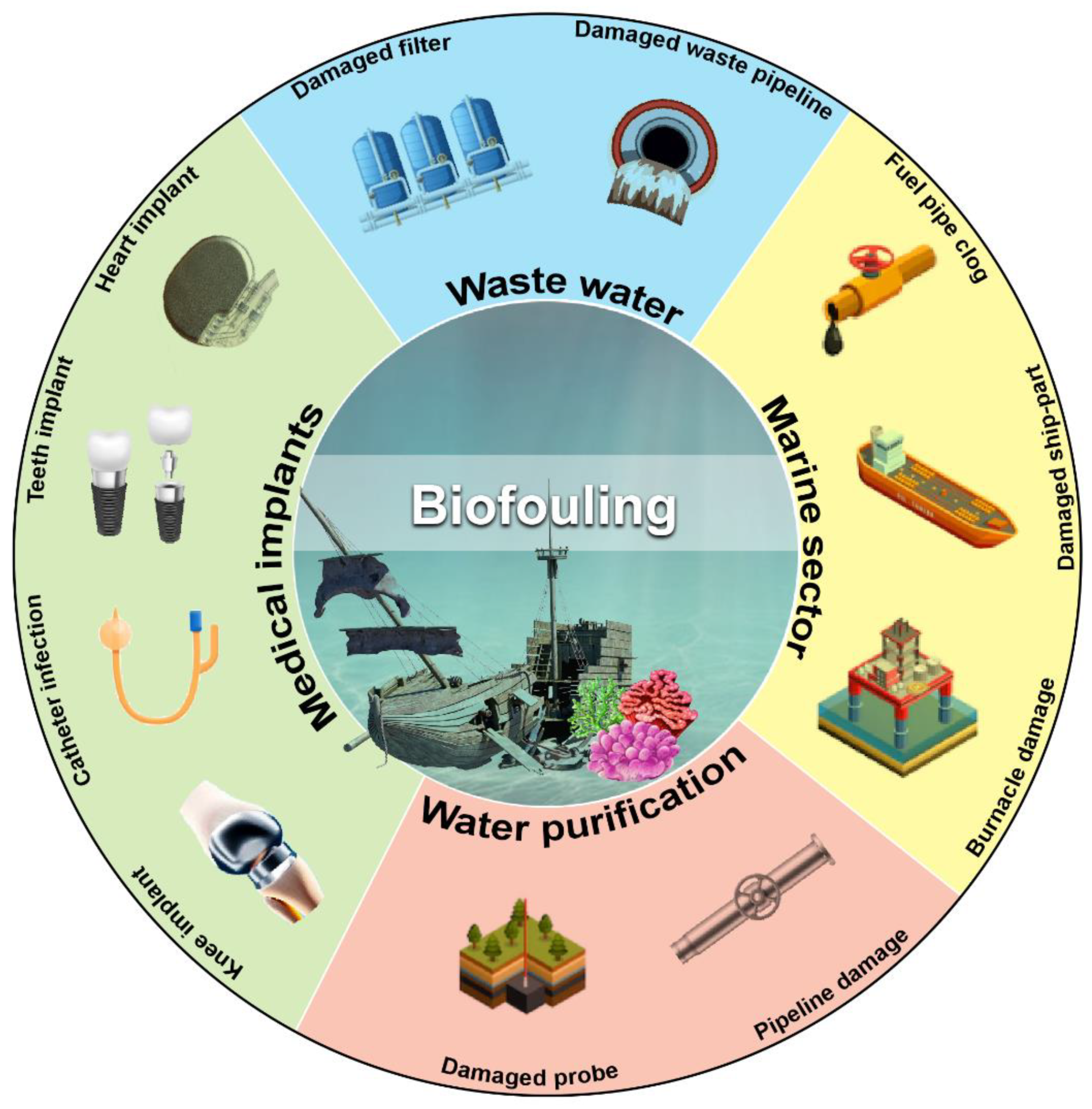
3. Biofilm-Mediated Biofouling
4. Antimicrobial Peptides from Marine Sources
5. Mechanisms of Action
5.1. Membrane Targeting Mechanism
5.1.1. Toroidal Pore Model
5.1.2. Barrel-Stave Model
5.1.3. Carpet-like Model
5.2. Non-Membrane Targeting Mechanism
6. Classification of Marine AMPs
6.1. AMPs from Bacterial Sources
6.2. AMPs from Marine Invertebrates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Peptides | References |
|---|---|---|
| Porifera peptides | Callyaerin A and B (antimicrobial) | [48] |
| Theonellamide F(antimicrobial) | [49] | |
| Theonellamide G(antimicrobial) | [49] | |
| Koshikamides F and H (antiviral) | [50] | |
| Celebesides A-C (antiviral) | [50] | |
| Mirabamides A–D (antiviral) | [51] | |
| Mirabamides E–H (antiviral) | [51] | |
| Stellettapeptins A and B (antiviral) | [52] | |
| Barettin and 8,9-dihydrobarettin (antifouling) | [53] | |
| Barrettides A and B (antifouling) | [54] | |
| Cnidaria peptides | Aurelin (antimicrobial) | [45] |
| Pd-AMP1 (antimicrobial) | [46] | |
| Mollusca peptides | Myticusin-1 (antimicrobial) | [55] |
| Mytichitin-CB (antimicrobial) | [56] | |
| Myticusin beta (antimicrobial) | [57] | |
| Octominin (antimicrobial) | [47] | |
| RpdefB (antimicrobial) | [58] | |
| VpMacin (antimicrobial) | [58] | |
| Myticin C (antiviral) | [59] | |
| Annelida peptides | Arenicin-1, 2, and 3 (antimicrobial) | [60] |
| Hedistin (antimicrobial) | [61] | |
| Nicomicin -1 (antimicrobial) | [62] | |
| Capitellacin (antimicrobial) | [62] | |
| Perinerin (antimicrobial) | [63] | |
| Arthropoda peptides | rSs-arasin (antimicrobial) | [64] |
| Sphistin (antimicrobial) | [65] | |
| Anti-lipopolysaccharides (ALFs)(ALFPm11) (antimicrobial) | [66] | |
| Crustin (antimicrobial) | [67] | |
| paralithocins 1–3 (antimicrobial) | [68] | |
| Echinodermata peptides | PpCrAMP (antimicrobial) | [69] |
| SdStrongylocin 1 and 2 (antimicrobial) | [70] | |
| EeCentrocin 1 and 2 (antimicrobial) | [71] | |
| EeStrongylocin 2 (antimicrobial) | [71] | |
| Chordata peptides | Pc-pis (antimicrobial) | [72] |
| CodCath (antimicrobial) | [73] | |
| RbLEAP-2 (antimicrobial) | [74] | |
| Styelin D (antimicrobial) | [75] |
6.3. AMPs from Marine Algae
7. Strategies for AMPs Extraction from Marine Sources
8. Advantages of Marine AMPs
8.1. Exhibit Early-Stage Killing Potential
8.2. Act in Varied Micro-Environment and Niches in Biofilm
8.3. Hinder Cell Propagation through Extracellular Matrix Interference
8.4. Stands in the Way of Bacterial Communication
8.5. Synergistic Action with Other Antimicrobial Drugs
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Li, F.; Wu, C.; Yu, R.; Wu, X.; Shen, L.; Liu, Y.; Qiu, G.; Li, J. Role of extracellular polymeric substance (EPS) in toxicity response of soil bacteria Bacillus sp. S3 to multiple heavy metals. Bioprocess. Biosyst. Eng. 2020, 43, 153–167. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C.C.R. Marine Biofilms: A successful microbial strategy with economic implications. Front. Mar. Sci. 2018, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control. 2019, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Schittek, B.; Hipfel, R.; Sauer, B. Dermcidin: A novel human antibiotic peptide secreted by sweat glands. Nat. Immunol. 2001, 2, 1133–1137. [Google Scholar] [CrossRef]
- Malkoski, M.; Dashper, S.G.; O’Brien-Simpson, N.M. Kappacin, a novel antibacterial peptide from bovine milk. Antimicrob. Agents Chemother. 2001, 45, 2309. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Villaruz, A.E.; Li, M. The human anionic antimicrobial peptide dermcidin induces proteolytic defence mechanisms in staphylococci. Mol. Microbiol. 2007, 63, 497–506. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.C.; Huang, S. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919. [Google Scholar]
- Koehbach, J.; Craik, D.J. The vast structural diversity of antimicrobial peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 2559. [Google Scholar] [CrossRef] [PubMed]
- Subbalakshmi, C.; Sitaram, N. Mechanism of antimicrobial action of indolicidin. FEMS Microbiol. Lett. 1998, 160, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, B.; Munoz-Garay, C. Marine antimicrobial peptides: A promising source of new generation antibiotics and other bio-active molecules. Int. J. Pept. Res. Ther. 2019, 25, 1441–1450. [Google Scholar] [CrossRef]
- Dalton, H.M.; Goodman, A.E.; Marshall, K.C. Diversity in surface colonization behavior in marine bacteria. J. Ind. Microbiol. 1996, 17, 228–234. [Google Scholar] [CrossRef]
- Johnson, B.D.; Azetsu-Scott, K. Adhesion force and the character of surfaces immersed in seawater. Limnol. Oceanogr. 1995, 40, 802–808. [Google Scholar] [CrossRef]
- Wahl, M. Marine epibiosis. I. Fouling and antifouling: Some basic aspects. Mar. Ecol. Prog. Ser. 1989, 58, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Railkin, A.I. Marine Biofouling: Colonization Processes and Defenses; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [Green Version]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S. Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Wang, J.; Chen, H.; Chen, D. Progress of marine biofouling and antifouling technologies. Chin. Sci. Bull. 2011, 56, 598–612. [Google Scholar] [CrossRef] [Green Version]
- Karpov, V.A.; Kovalchuk, Y.L.; Kharchenko, U.V.; Beleneva, I.A. The effect of microfouling on marine corrosion of metals and destruction of protective coatings. Prot. Met. Phys. Chem. Surf. 2012, 48, 803–809. [Google Scholar] [CrossRef]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic impact of biofouling on a naval surface ship. Biofouling 2011, 27, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Fitridge, I.; Dempster, T.; Guenther, J.; de Nys, R. The impact and control of biofouling in marine aquaculture: A review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Floerl, O.; Sunde, L.; Bloecher, N. Potential environmental risks associated with biofouling management in salmon aquaculture. Aquac. Environ. Interact. 2016, 8, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [Green Version]
- Mora, C.; Tittensor, D.P.; Adl, S. How many species are there on earth and in the Ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Brown, K.L.; Mookherjee, N. Host defence peptides from invertebrates—Emerging antimicrobial strategies. Immunobiology 2006, 211, 315–322. [Google Scholar] [CrossRef]
- Yin, Z.X.; He, W.; Chen, W.J. Cloning, expression and antimicrobial activity of an antimicrobial peptide, epinecidin-1, from the orange-spotted grouper, Epinephelus coioides. Aquaculture 2006, 253, 204–211. [Google Scholar] [CrossRef]
- Acosta, J.; Montero, V.; Carpio, Y. Cloning and functional characterization of three novel antimicrobial peptides from tilapia (Oreochromis niloticus). Aquaculture 2013, 372–375, 9–18. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Semreen, M.H.; El-Gamal, M.I.; Abdin, S. Recent updates of marine antimicrobial peptides. Saudi Pharm. J. 2018, 26, 396. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E. Cationic peptides: Effectors in innate immunity and novel antimicrobials. Lancet Infect. Dis. 2001, 1, 156–164. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Murase, O.; Fujii, N.; Miyajima, K. Translocation of a channel-forming antimicrobial peptide, magainin 2, across lipid bilayers by forming a pore. Biochemistry 1995, 34, 6521–6526. [Google Scholar] [CrossRef] [PubMed]
- Omardien, S.; Drijfhout, J.W.; Vaz, F.M. Bactericidal activity of amphipathic cationic antimicrobial peptides involves altering the membrane fluidity when interacting with the phospholipid bilayer. Biochim. Biophys. Acta Biomembr. 2018, 1860, 2404–2415. [Google Scholar] [CrossRef] [PubMed]
- Lohner, K.; Prossnigg, F. Biological activity and structural aspects of PGLa interaction with membrane mimetic systems. Biochim. Biophys. Acta 2009, 1788, 1656–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipkin, R.B.; Lazaridis, T. Implicit membrane investigation of the stability of antimicrobial peptide β-barrels and arcs. J. Membr. Biol. 2015, 248, 469–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolym. Pept. Sci. Sect. 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Corrêa, J.A.F.; Evangelista, A.G.; de Melo Nazareth, T.; Luciano, F.B. Fundamentals on the molecular mechanism of action of antimicrobial peptides. Materialia 2019, 8, 100494. [Google Scholar] [CrossRef]
- Lyu, Y.; Fitriyanti, M.; Narsimhan, G. Nucleation and growth of pores in 1,2-Dimyristoyl-sn-glycero-3-phosphocholine (DMPC)/cholesterol bilayer by antimicrobial peptides melittin, its mutants and cecropin P1. Colloids Surf. B Biointerfaces 2019, 173, 121–127. [Google Scholar] [CrossRef]
- Mardirossian, M.; Grzela, R.; Giglione, C. The host antimicrobial peptide Bac71-35 binds to bacterial ribosomal proteins and inhibits protein synthesis. Chem. Biol. 2014, 21, 1639–1647. [Google Scholar] [CrossRef] [Green Version]
- Shu, G.; Chen, Y.; Liu, T.; Ren, S.; Kong, Y. Antimicrobial peptide cathelicidin-bf inhibits platelet aggregation by blocking protease-activated receptor 4. Int. J. Pept. Res. Ther. 2019, 25, 349–358. [Google Scholar] [CrossRef]
- Desriac, F.; Jégou, C.; Balnois, E. Antimicrobial peptides from marine proteobacteria. Mar. Drugs 2013, 11, 3632–3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klompen, A.M.L.; Macrander, J.; Reitzel, A.M.; Stampar, S.N. transcriptomic analysis of four cerianthid (cnidaria, ceriantharia) venoms. Mar. Drugs 2020, 18, 413. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Balandin, S.V.; Aleshina, G.M. Aurelin, a novel antimicrobial peptide from jellyfish Aurelia aurita with structural features of defensins and channel-blocking toxins. Biochem. Biophys. Res. Commun. 2006, 348, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.; Migliolo, L.; Castro, C. Identification of a novel antimicrobial peptide from Brazilian coast coral Phyllogorgia dilatata. Protein Pept. Lett. 2013, 20, 1153–1158. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dananjaya, S.H.S.; Chandrarathna, H.P.S.U. Octominin: A novel synthetic anticandidal peptide derived from defense protein of octopus minor. Mar. Drugs 2020, 18, 56. [Google Scholar] [CrossRef] [Green Version]
- Daletos, G.; Kalscheuer, R.; Koliwer-Brandl, H. Callyaerins from the marine sponge Callyspongia aerizusa: Cyclic peptides with antitubercular activity. J. Nat. Prod. 2015, 78, 1910–1925. [Google Scholar] [CrossRef]
- Nishimura, S.; Arita, Y.; Honda, M. Marine antifungal theonellamides target 3beta-hydroxysterol to activate Rho1 signaling. Nat. Chem. Biol. 2010, 6, 519–526. [Google Scholar] [CrossRef]
- Plaza, A.; Bifulco, G.; Keffer, J.L. Celebesides A-C and theopapuamides B-D, depsipeptides from an indonesian sponge that inhibit HIV-1 entry. J. Org. Chem. 2009, 74, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Van Wagoner, R.M.; Harper, M.K. Mirabamides, E-H, HIV-inhibitory depsipeptides from the sponge Stelletta clavosa. J. Nat. Prod. 2011, 74, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.J.; Rashid, M.A.; Cartner, L.K.; Bokesch, H.R.; Wilson, J.A.; McMahon, J.B.; Gustafson, K.R. Stellettapeptins A and B, HIV-inhibitory cyclic depsipeptides from the marine sponge Stelletta sp. Tetrahedron Lett. 2015, 56, 4215–4219. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, M.; Göransson, U.; Johnson, A.L. Antifouling activity of brominated cyclopeptides from the marine sponge Geodia barretti. J. Nat. Prod. 2004, 67, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, M.; Johnson, A.L.; Hedner, E. Antifouling activity of synthesized peptide analogs of the sponge metabolite barettin. Peptides 2006, 27, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Wang, X.C.; Liu, H.H.; Fan, M.H.; Sun, J.J.; Shen, W. Molecular characterization of a novel antimicrobial peptide from Mytilus coruscus. Fish Shellfish Immunol. 2013, 34, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Oh, R.; Lee, M.J.; Kim, Y.O. Myticusin-beta, antimicrobial peptide from the marine bivalve, Mytilus coruscus. Fish Shellfish Immunol. 2020, 99, 342–352. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Q.; Wang, Q. A defensin-like antimicrobial peptide from the manila clam Ruditapes philippinarum: Investigation of the antibacterial activities and mode of action. Fish Shellfish Immunol. 2018, 80, 274–280. [Google Scholar] [CrossRef]
- Yang, D.; Han, Y.; Chen, L.; Cao, R.; Wang, Q.; Dong, Z.; Liu, H.; Zhang, X.; Zhang, Q.; Zhao, J. A macin identified from Venerupis philippinarum: Investigation on antibacterial activities and action mode. Fish Shellfish Immunol. 2019, 92, 897–904. [Google Scholar] [CrossRef]
- Novoa, B.; Romero, A.; Álvarez, Á.L. Antiviral activity of myticin C peptide from mussel: An ancient defense against herpesviruses. J. Virol. 2016, 90, 7692–7702. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, X.; Teng, D. Candidacidal mechanism of the arenicin-3-derived peptide NZ17074 from Arenicola marina. Appl. Microbiol. Biotechnol. 2014, 98, 7387–7398. [Google Scholar] [CrossRef]
- Tasiemski, A.; Schikorski, D.; Le Marrec-Croq, F.; Pontoire-Van Camp, C.; Boidin-Wichlacz, C.; Sautière, P.E. Hedistin: A novel antimicrobial peptide containing bromotryptophan constitutively expressed in the NK cells-like of the marine annelid, Nereis diversicolor. Dev. Comp. Immunol. 2007, 31, 749–762. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Tsarev, A.V.; Safronova, V.N. Structure elucidation and functional studies of a novel β-hairpin antimicrobial peptide from the marine polychaeta Capitella teleta. Mar. Drugs 2020, 18, 620. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Liu, X.; Ge, F. Perinerin, a novel antimicrobial peptide purified from the clamworm Perinereis aibuhitensis grube and its partial characterization. J. Biochem. 2004, 135, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Anju, A.; Smitha, C.K.; Preetha, K. Molecular characterization, recombinant expression and bioactivity profile of an antimicrobial peptide, Ss-arasin from the Indian mud crab, Scylla serrata. Fish Shellfish Immunol. 2019, 88, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Fan, D.Q.; Zhu, K.X. Mechanism study on a new antimicrobial peptide Sphistin derived from the N-terminus of crab histone H2A identified in haemolymphs of Scylla paramamosain. Fish Shellfish Immunol. 2015, 47, 833–846. [Google Scholar] [CrossRef]
- Zhou, L.; Li, G.; Jiao, Y.; Huang, D.; Li, A.; Chen, H.; Liu, Y.; Li, S.; Li, H.; Wang, C. Molecular and antimicrobial characterization of a group G anti-lipopolysaccharide factor (ALF) from Penaeus monodon. Fish Shellfish Immunol. 2019, 94, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Antony, S.P.; Singh, I.S.B.; Sudheer, N.S. Molecular characterization of a crustin-like antimicrobial peptide in the giant tiger shrimp, Penaeus monodon, and its expression profile in response to various immunostimulants and challenge with WSSV. Immunobiology 2011, 216, 184–194. [Google Scholar] [CrossRef]
- Moe, M.K.; Haug, T.; Sydnes, M.O. Paralithocins, antimicrobial peptides with unusual disulfide connectivity from the red king crab, Paralithodes camtschaticus. J. Nat. Prod. 2018, 81, 140–150. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Go, H.J.; Oh, H.Y. Identification of a novel antimicrobial peptide from the sea star Patiria pectinifera. Dev. Comp. Immunol. 2018, 86, 203–213. [Google Scholar] [CrossRef]
- Solstad, R.G.; Li, C.; Isaksson, J. Novel antimicrobial peptides eecentrocins 1, 2 and eestrongylocin 2 from the edible sea urchin Echinus esculentus have 6-br-trp post-translational modifications. PLoS ONE 2016, 11, e0151820. [Google Scholar] [CrossRef] [Green Version]
- Solstad, R.G.; Johansen, C.; Stensvåg, K. Structure-activity relationship studies of shortened analogues of the antimicrobial peptide EeCentrocin 1 from the sea urchin Echinus esculentus. J. Pept. Sci. 2020, 26, e3233. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.F.; Jin, Y.; Xu, X. Characterization of a novel piscidin-like antimicrobial peptide from Pseudosciaena crocea and its immune response to Cryptocaryon irritans. Fish Shellfish Immunol. 2013, 35, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Broekman, D.C.; Zenz, A.; Gudmundsdottir, B.K. Functional characterization of codCath, the mature cathelicidin antimicrobial peptide from Atlantic cod (Gadus morhua). Peptides 2011, 32, 2044–2051. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.D.; Joo, M.S.; Hwang, J.Y. Molecular characterization and gene expression data of liver expressed antimicrobial Peptide-2 (LEAP-2) isolated from rock bream (Oplegnathus fasciatus). Data Br. 2019, 26, 104538. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.W.; Craig, A.G.; Fischer, W.H.; Park, M.; Lehrer, R.I. Styelin D, an extensively modified antimicrobial peptide from ascidian hemocytes. J. Biol. Chem. 2000, 275, 38417–38426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.K.; Seo, C.H.; Park, Y. Marine peptides and their anti-infective activities. Mar. Drugs 2015, 13, 618–654. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K. Antimicrobial bioactive compounds from marine algae: A mini review. Rev. Artic. Indian J. Geo-Mar. Sci. 2016, 45, 1076–1085. [Google Scholar]
- Smith, D.; Buddie, A.G.; Goss, R.J.M. Discovery pipelines for marine resources: An ocean of opportunity for biotechnology? World J. Microbiol. Biotechnol. 2019, 35, 107. [Google Scholar] [CrossRef] [Green Version]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Xing, R.; Li, P. Characterization, preparation, and purification of marine bioactive peptides. Biomed. Res. Int. 2017, 2017, 9746720. [Google Scholar] [CrossRef] [Green Version]
- Charlet, M.; Chernysh, S.; Philippe, H. Innate immunity: Isolation of several cysteine-rich antimicrobial peptides from the blood of a mollusc, Mytilus edulis. J. Biol. Chem. 1996, 271, 21808–21813. [Google Scholar] [CrossRef] [Green Version]
- Relf, J.M.; Chisholm, J.R.S.; Kemp, G.D.; Smith, V.J. Purification and characterization of a cysteine-rich 11.5-kDa antibacterial protein from the granular haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1999, 264, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, C.; Chen, A. Molecular characterization of a novel big defensin from clam Venerupis philippinarum. PLoS ONE 2010, 5, e13480. [Google Scholar] [CrossRef] [PubMed]
- Shike, H.; Lauth, X.; Westerman, M.E. Bass hepcidin is a novel antimicrobial peptide induced by bacterial challenge. Eur. J. Biochem. 2002, 269, 2232–2237. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chang, Q.; Wu, M.; Zhao, X. Total synthesis of five proline-enriched cyclic heptapeptides from the marine sponge Stylissa carteri. Tetrahedron Lett. 2018, 59, 1828–1831. [Google Scholar] [CrossRef]
- Chen, T.; Zhou, J.; Qu, Z. Administration of dietary recombinant hepcidin on grass carp (Ctenopharyngodon idella) against Flavobacterium columnare infection under cage aquaculture conditions. Fish Shellfish Immunol. 2020, 99, 27–34. [Google Scholar] [CrossRef]
- Sperstad, S.V.; Haug, T.; Blencke, H.M. Antimicrobial peptides from marine invertebrates: Challenges and perspectives in marine antimicrobial peptide discovery. Biotechnol. Adv. 2011, 29, 519–530. [Google Scholar] [CrossRef]
- Lebeaux, D.; Ghigo, J.-M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.; Pulicherla, K.K. Psychrophiles as the Source for Potential Industrial Psychrozymes. In Recent Developments in Microbial Technologies; Prasad, R., Kumar, V., Singh, J., Upadhyaya, C.P., Eds.; Environmental and Microbial Biotechnology; Springer: Singapore, 2021. [Google Scholar] [CrossRef]
- Ghosh, M.; Gera, M.; Singh, J.; Prasad, R.; Pulicherla, K.K. A Comprehensive Investigation of Potential Novel Marine Psychrotolerant Actinomycetes sp. Isolated from the Bay-of-Bengal. Curr. Genom. 2020, 21, 271–282. [Google Scholar] [CrossRef]
- Sun, H.; Hong, Y.; Xi, Y. Synthesis, self-assembly, and biomedical applications of antimicrobial peptide-polymer conjugates. Biomacromolecules 2018, 19, 1701–1720. [Google Scholar] [CrossRef]
- Shang, D.; Han, X.; Du, W. Trp-containing antibacterial peptides impair quorum sensing and biofilm development in multidrug-resistant Pseudomonas aeruginosa and exhibit synergistic effects with antibiotics. Front. Microbiol. 2021, 12, 185. [Google Scholar] [CrossRef]




| Functional Type | Number of AMPs | Examples |
|---|---|---|
| Antibiofilm peptide | 76 | Pleurocidin, Nisin A, Gramicidin S |
| Antibacterial peptide | 2900 | Ericin S, Bactericidin B-3, Pleurocidin |
| Antifungal peptide | 1257 | Catfish PACAP38, Trematocine, Moronecidin-like |
| Antiparasitic peptide | 140 | Bombinin H4, HbbetaP-1, Piscidin 2 |
| Insecticidal peptide | 41 | Magainin 2, Esculentin-1 |
| Ion-channel inhibitor | 7 | Microcin H47, Bldesin |
| Protease inhibitor | 33 | Odorranain-B1, Microcin H47, Kunitzin-OS |
| Surface immobilized peptides | 31 | Magainin 2, Nisin A, Chrysophsin-1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patra, A.; Das, J.; Agrawal, N.R.; Kushwaha, G.S.; Ghosh, M.; Son, Y.-O. Marine Antimicrobial Peptides-Based Strategies for Tackling Bacterial Biofilm and Biofouling Challenges. Molecules 2022, 27, 7546. https://doi.org/10.3390/molecules27217546
Patra A, Das J, Agrawal NR, Kushwaha GS, Ghosh M, Son Y-O. Marine Antimicrobial Peptides-Based Strategies for Tackling Bacterial Biofilm and Biofouling Challenges. Molecules. 2022; 27(21):7546. https://doi.org/10.3390/molecules27217546
Chicago/Turabian StylePatra, Anupam, Jhilik Das, Nupur Rani Agrawal, Gajraj Singh Kushwaha, Mrinmoy Ghosh, and Young-Ok Son. 2022. "Marine Antimicrobial Peptides-Based Strategies for Tackling Bacterial Biofilm and Biofouling Challenges" Molecules 27, no. 21: 7546. https://doi.org/10.3390/molecules27217546