

Inhibition of NF-kB/IL-6/JAK2/STAT3 Pathway and Epithelial-Mesenchymal Transition in Breast Cancer Cells by Azilsartan
 , ,
, , 
Abstract
:1. Introduction
2. Results
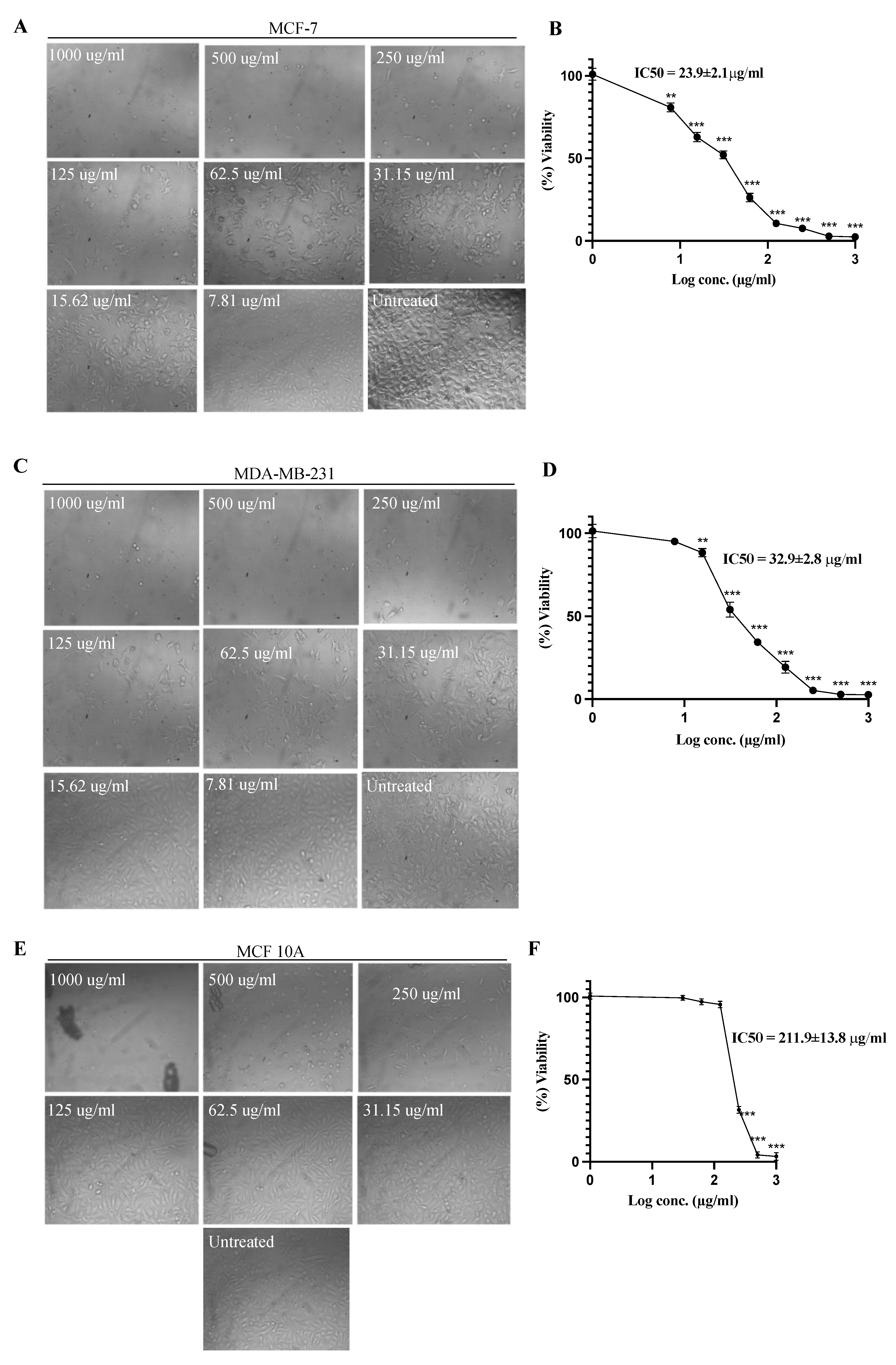
2.1. Cell Viability Assay
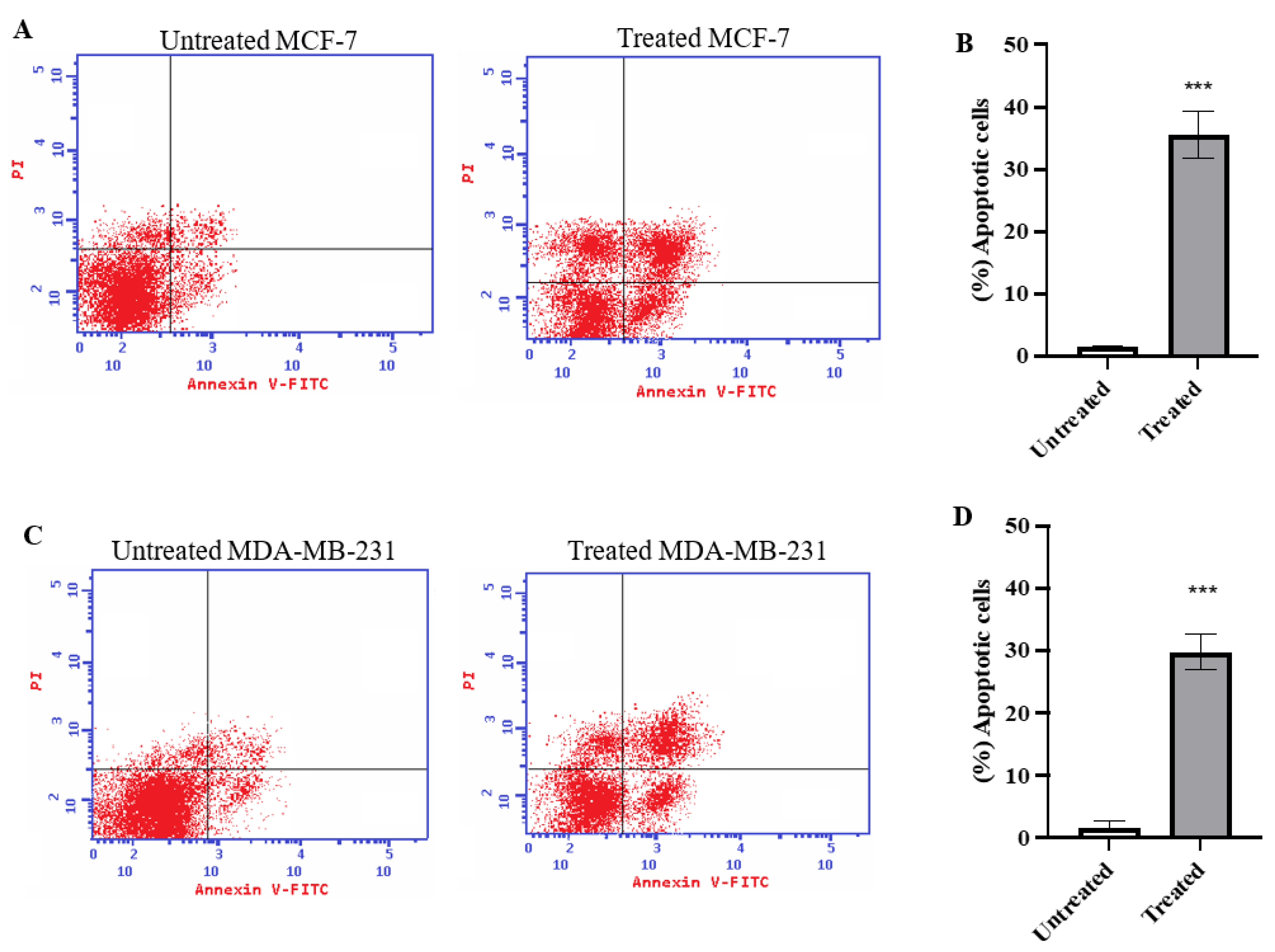
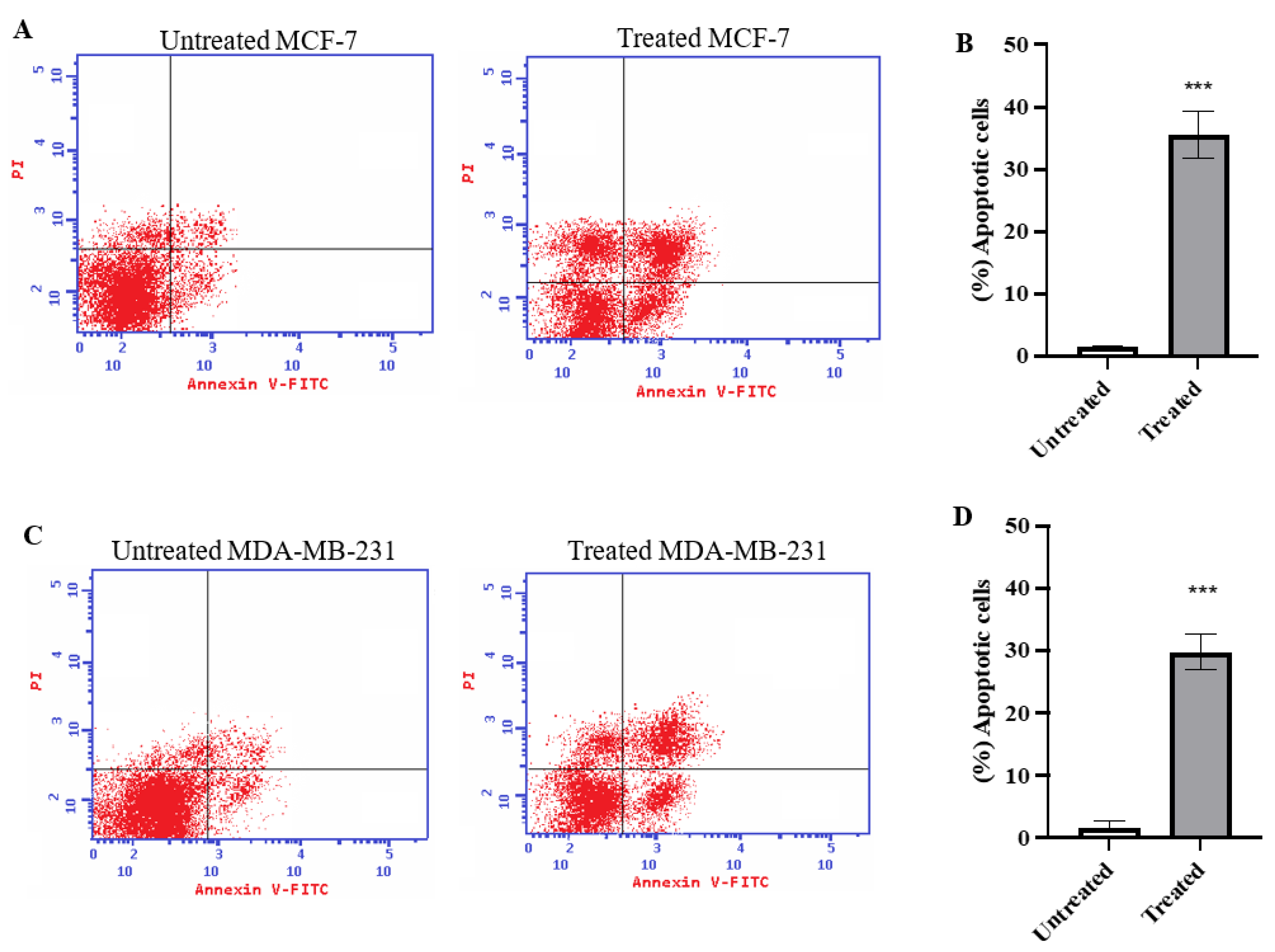
2.2. Annexin V Analysis
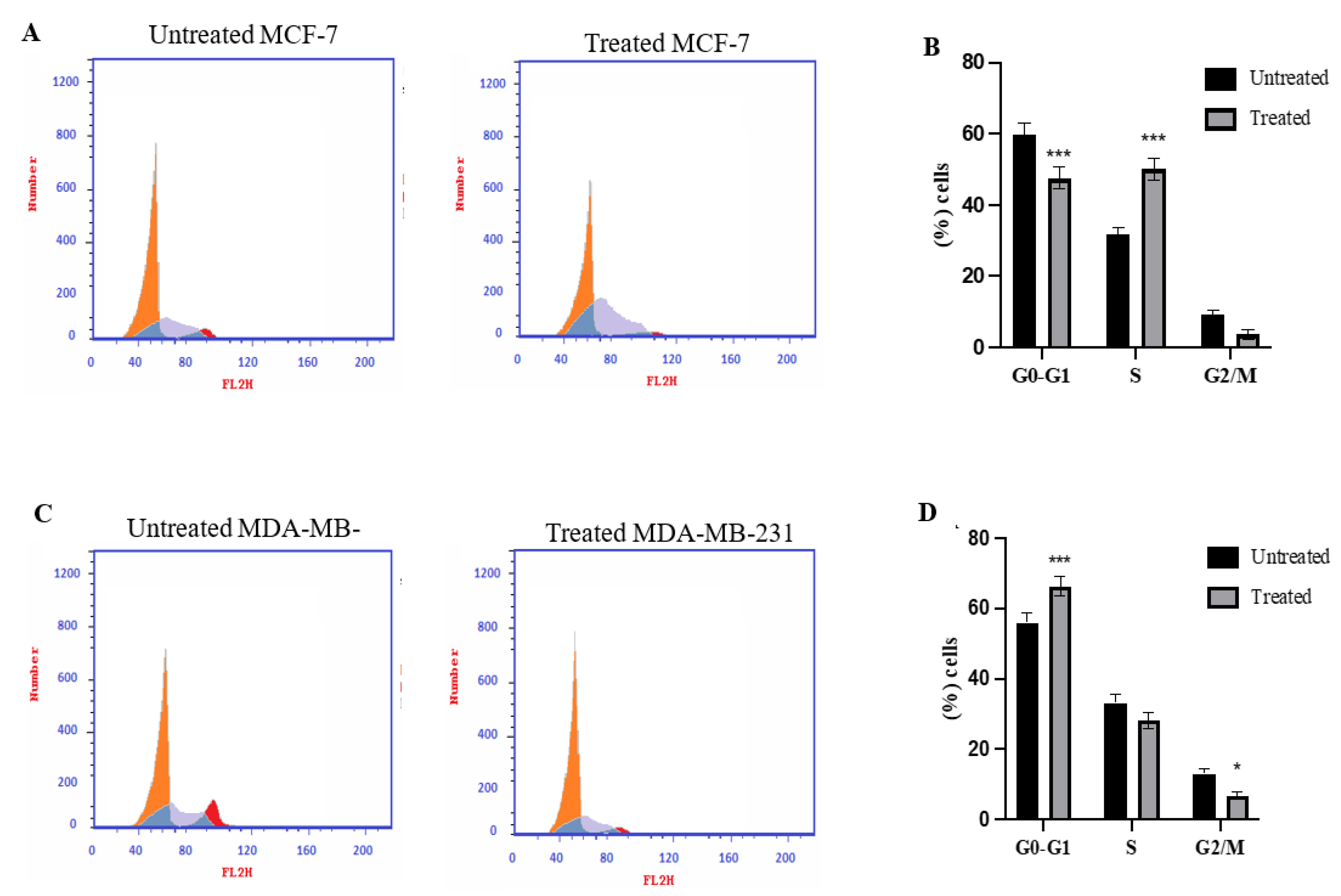
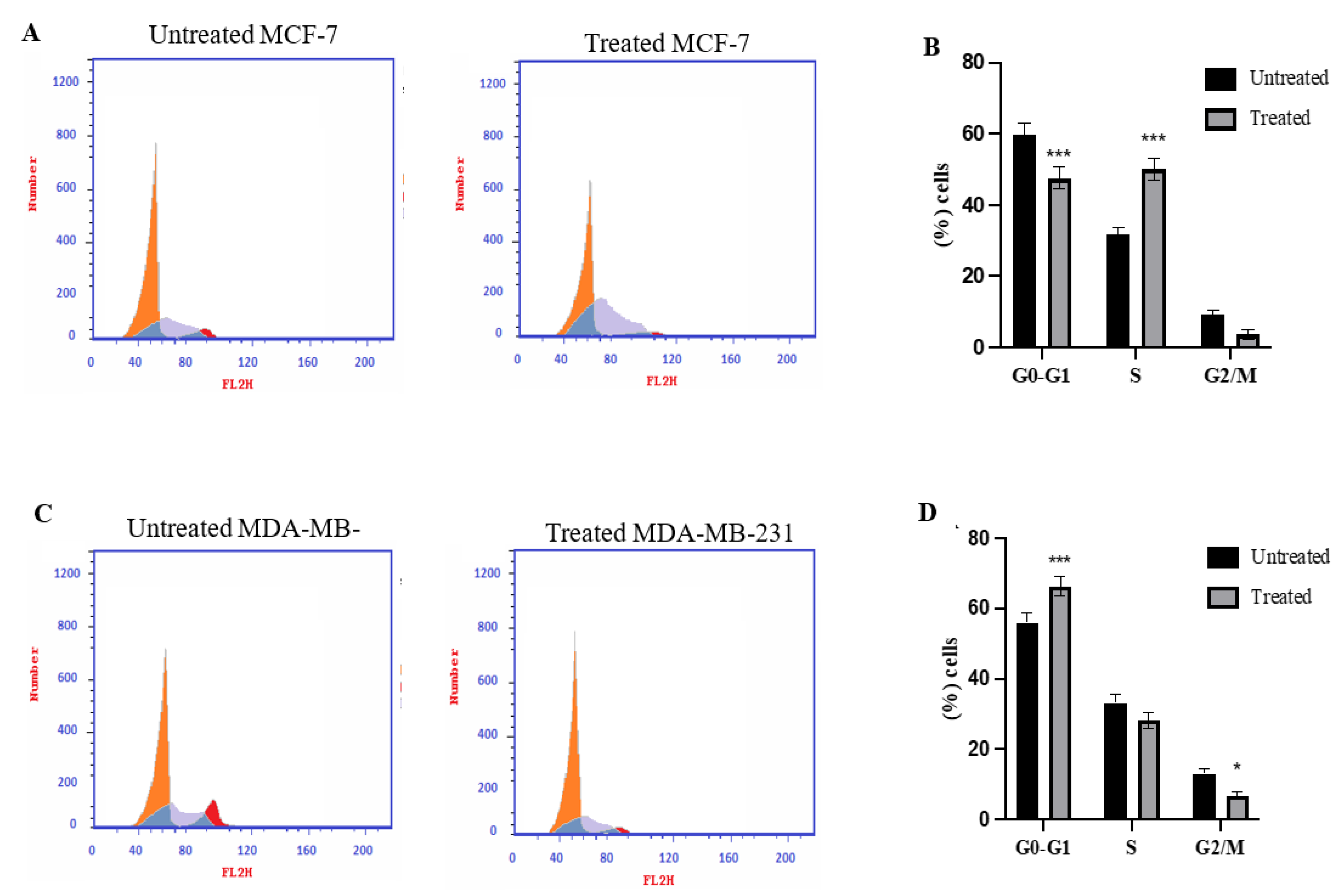
2.3. Cell Cycle Analysis Using Flowcytometry
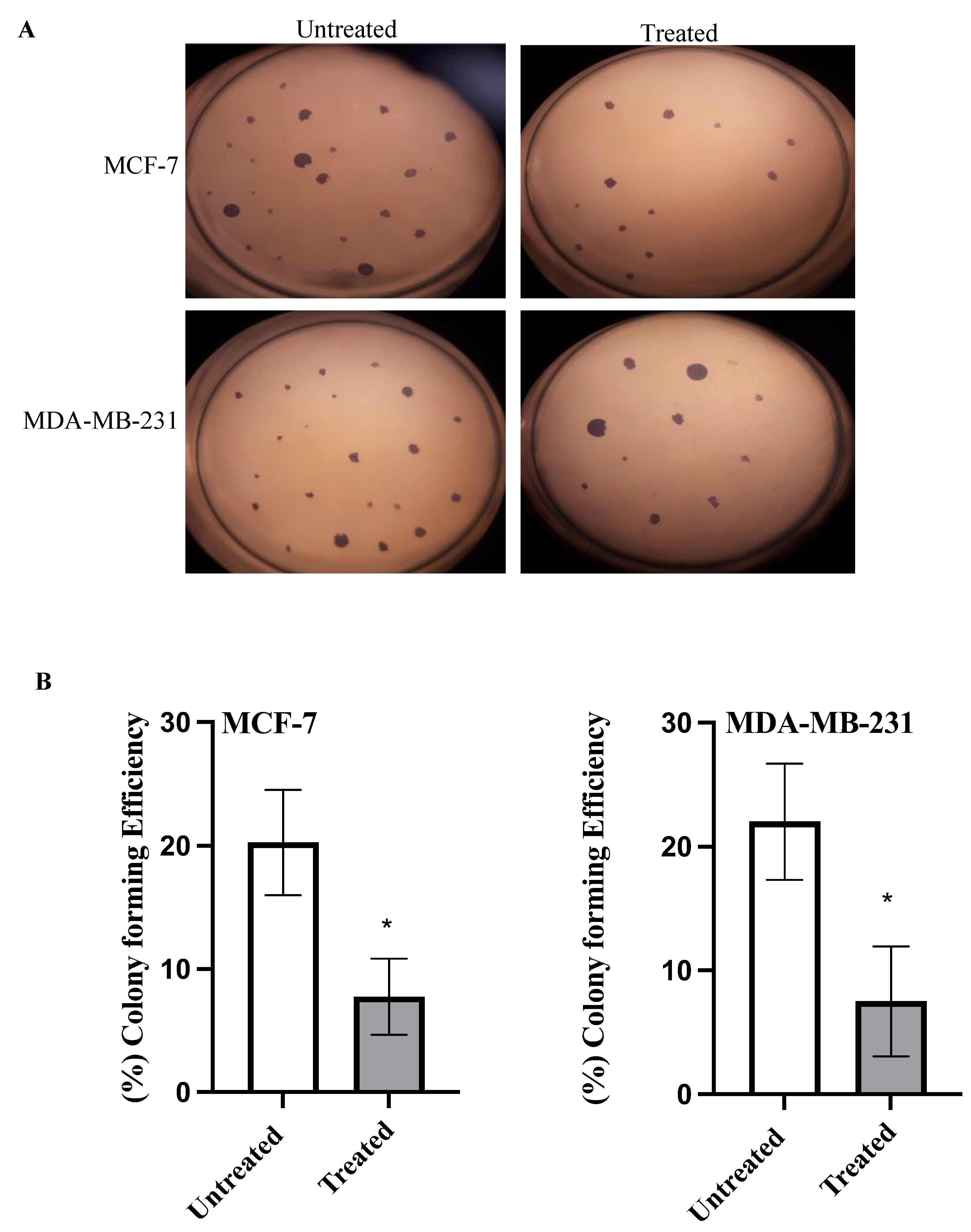
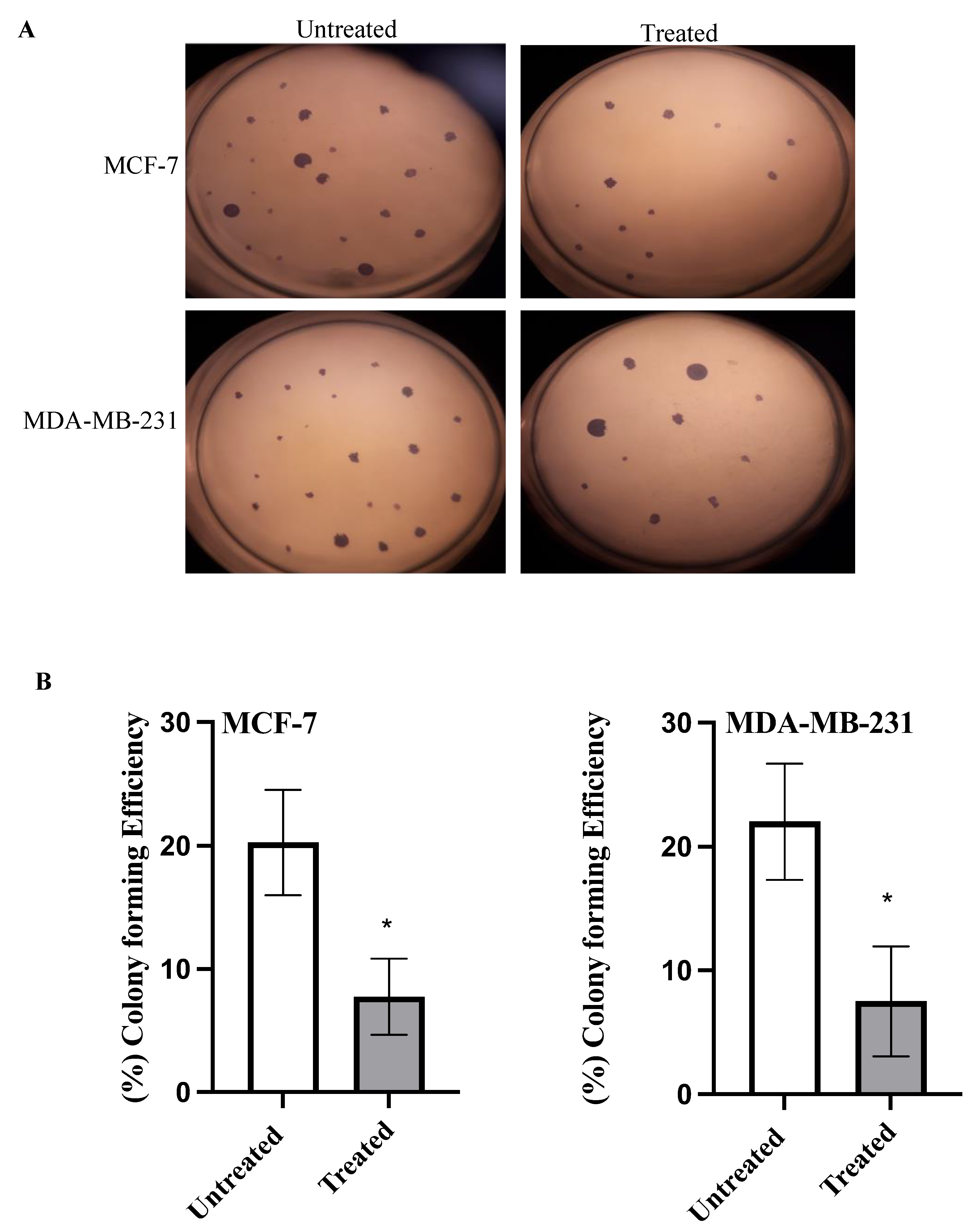
2.4. Colony Formation Assay
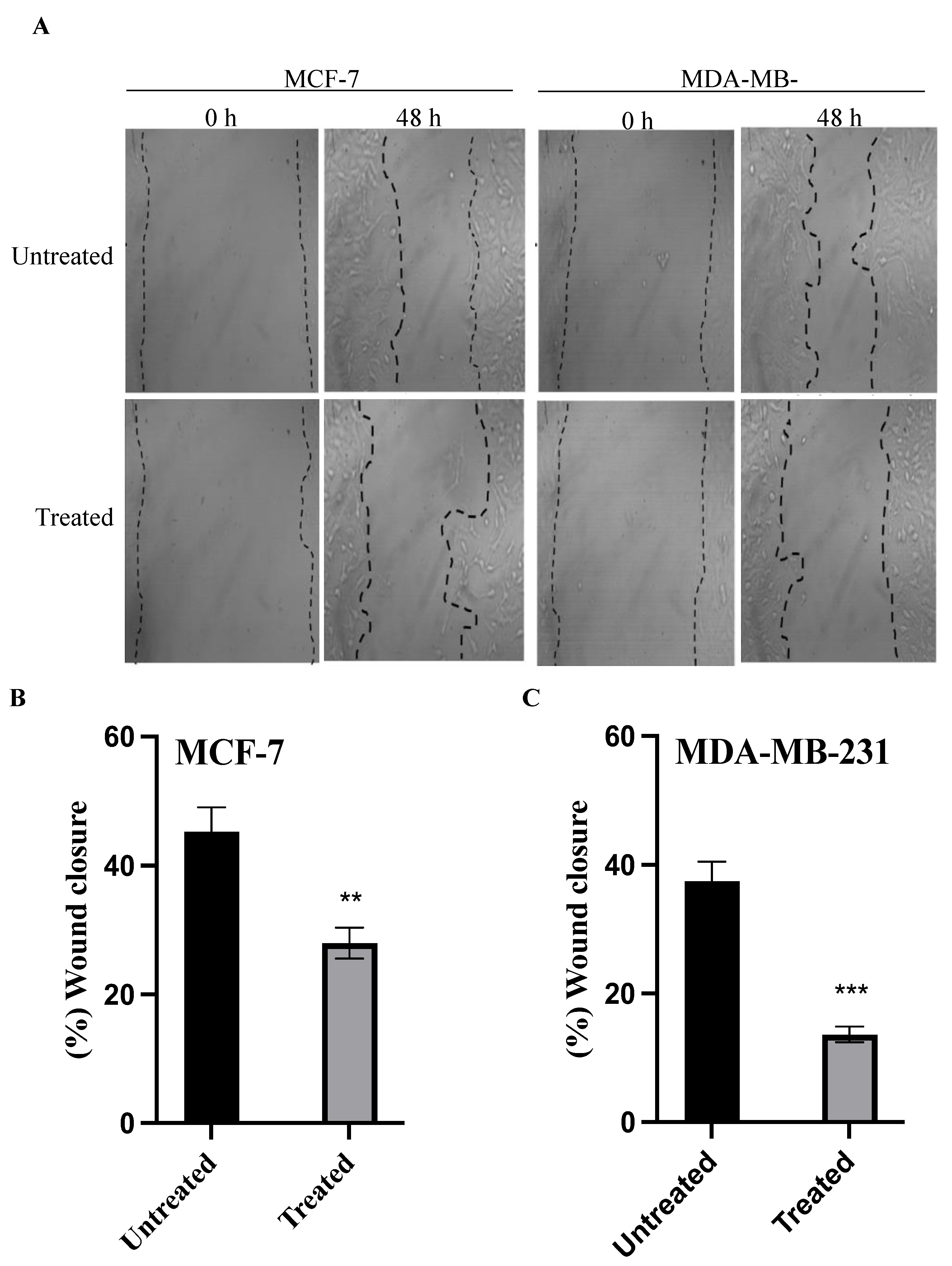
2.5. Cell Migration Assay
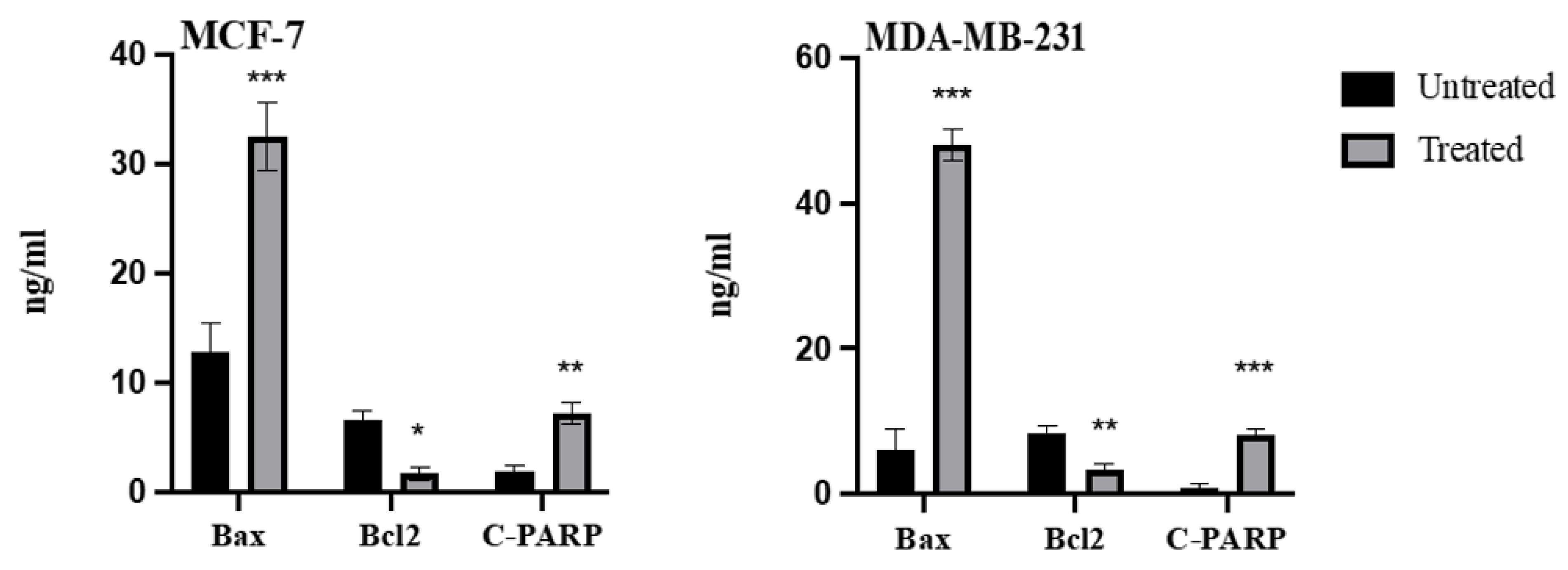
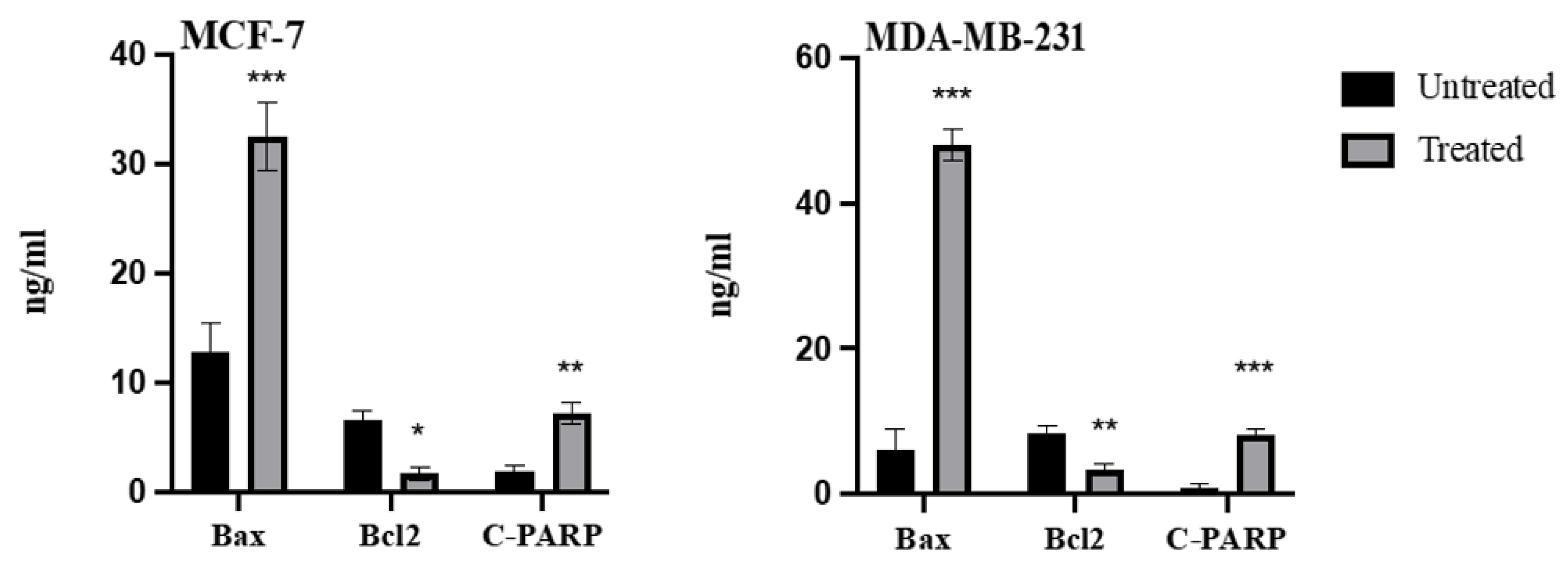
2.6. Effect of Azilsartan on the Levels of Bax, Bcl2, Cleaved PARP Proteins
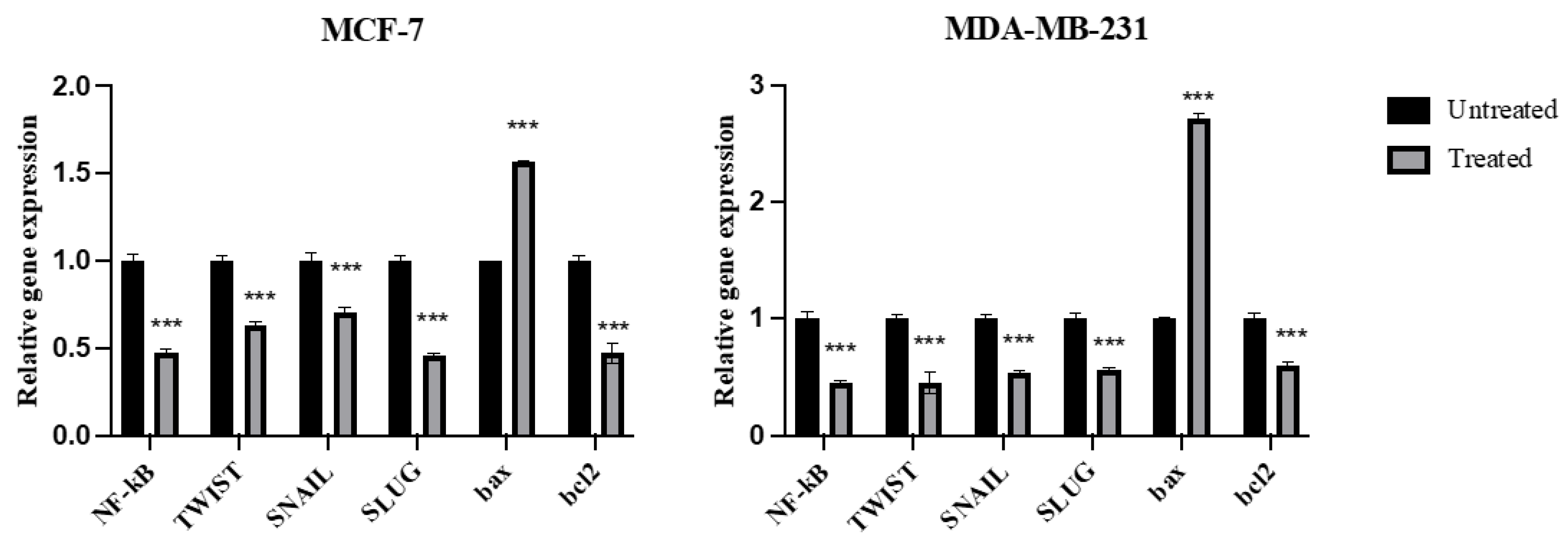
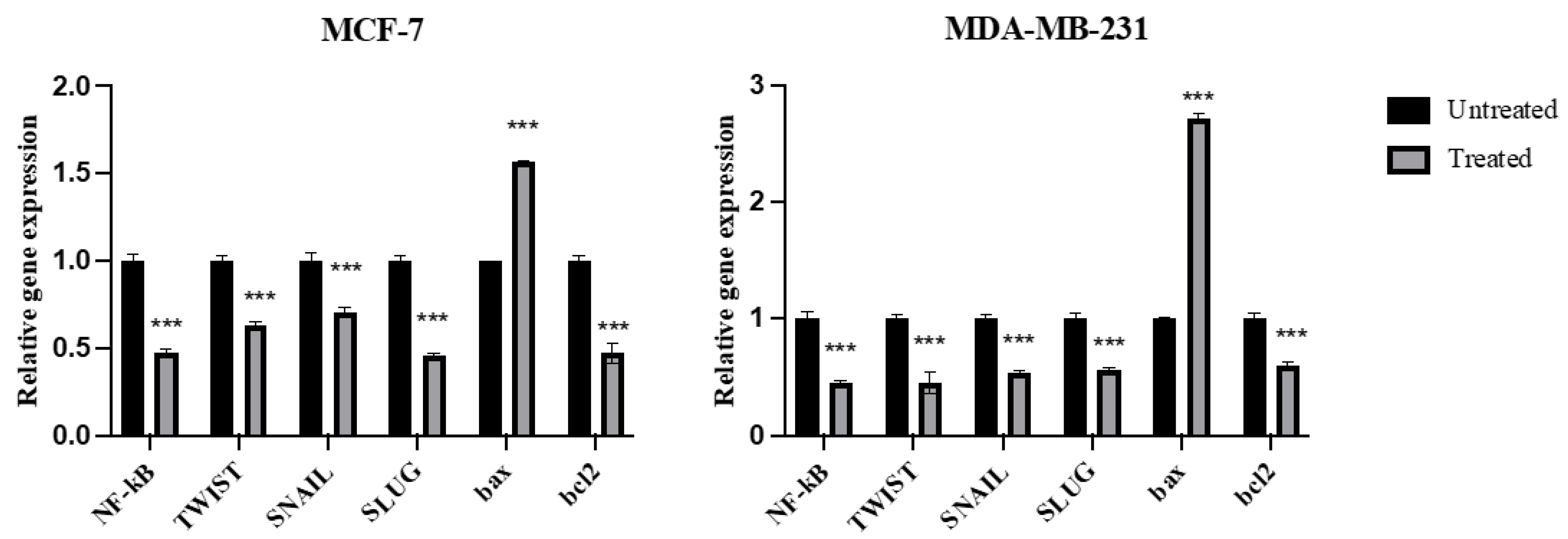
2.7. Expression of NF-kB, TWIST, SNAIL, SLUG, Bax, and Bcl2 Genes
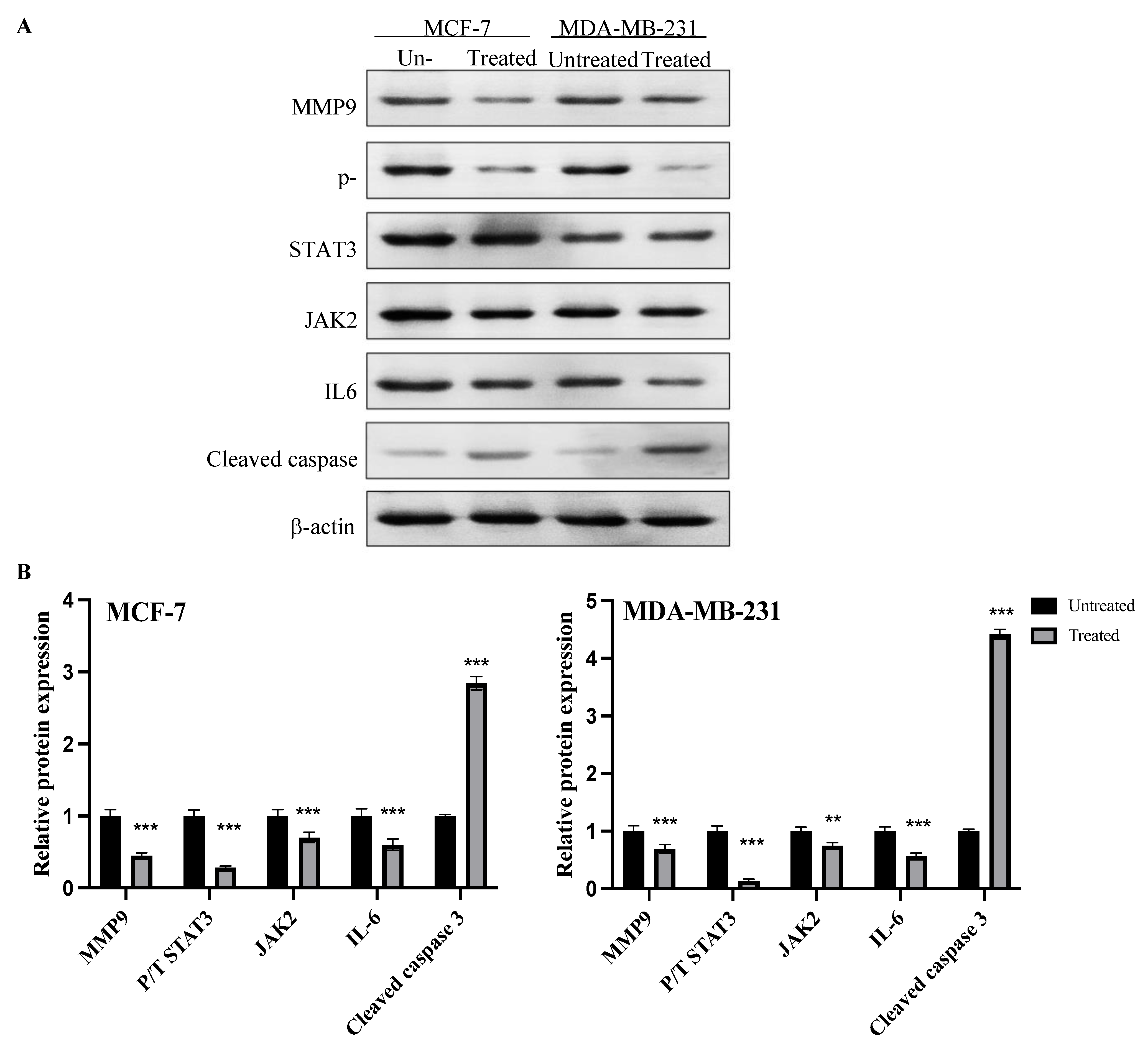
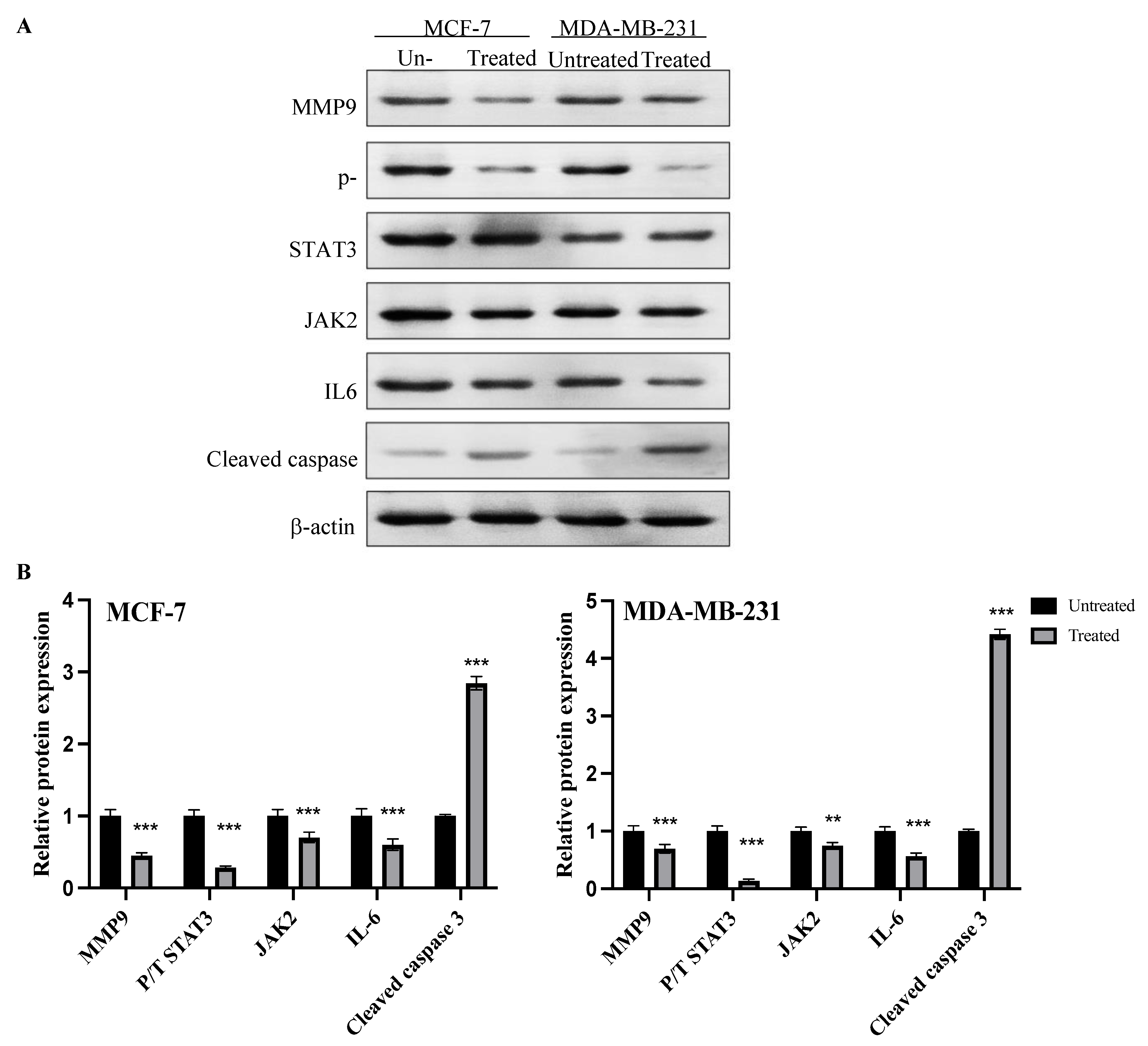
2.8. Expression of MMP9, p-STAT3, JAK2, IL6 and Cleaved Caspase Proteins
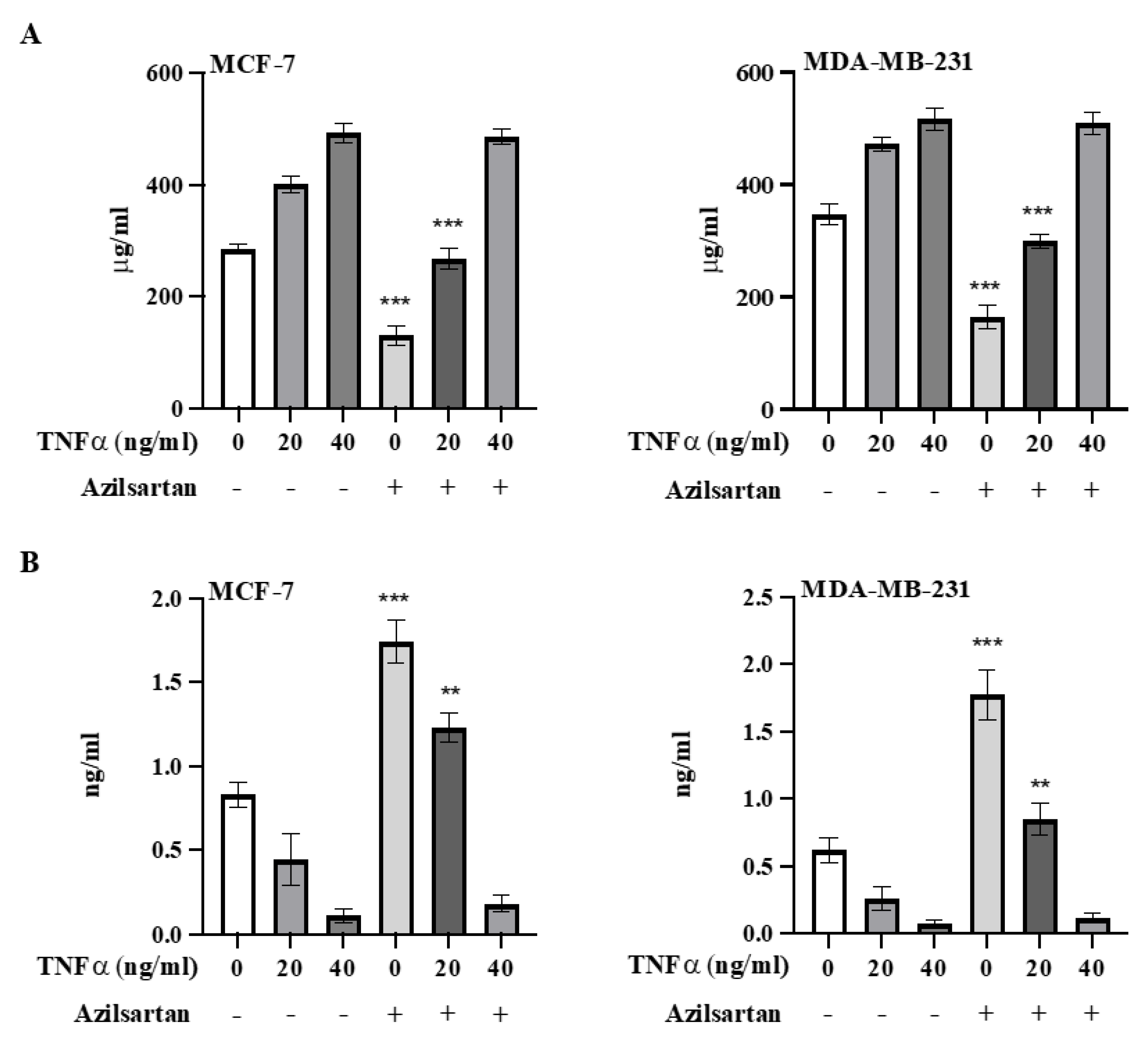
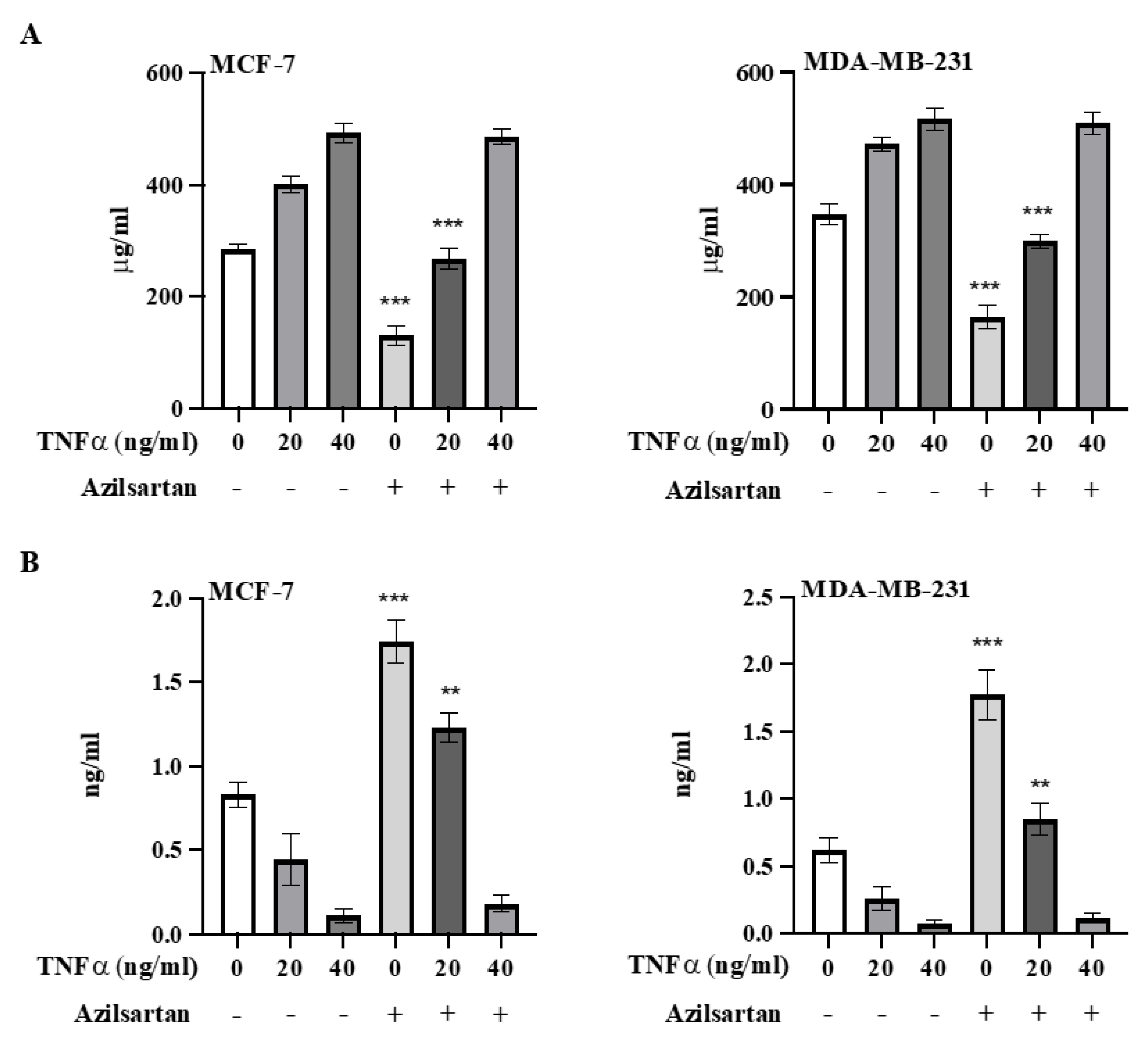
2.9. Effect of Azilsartan on TNFα-Induced NF-kB Activation
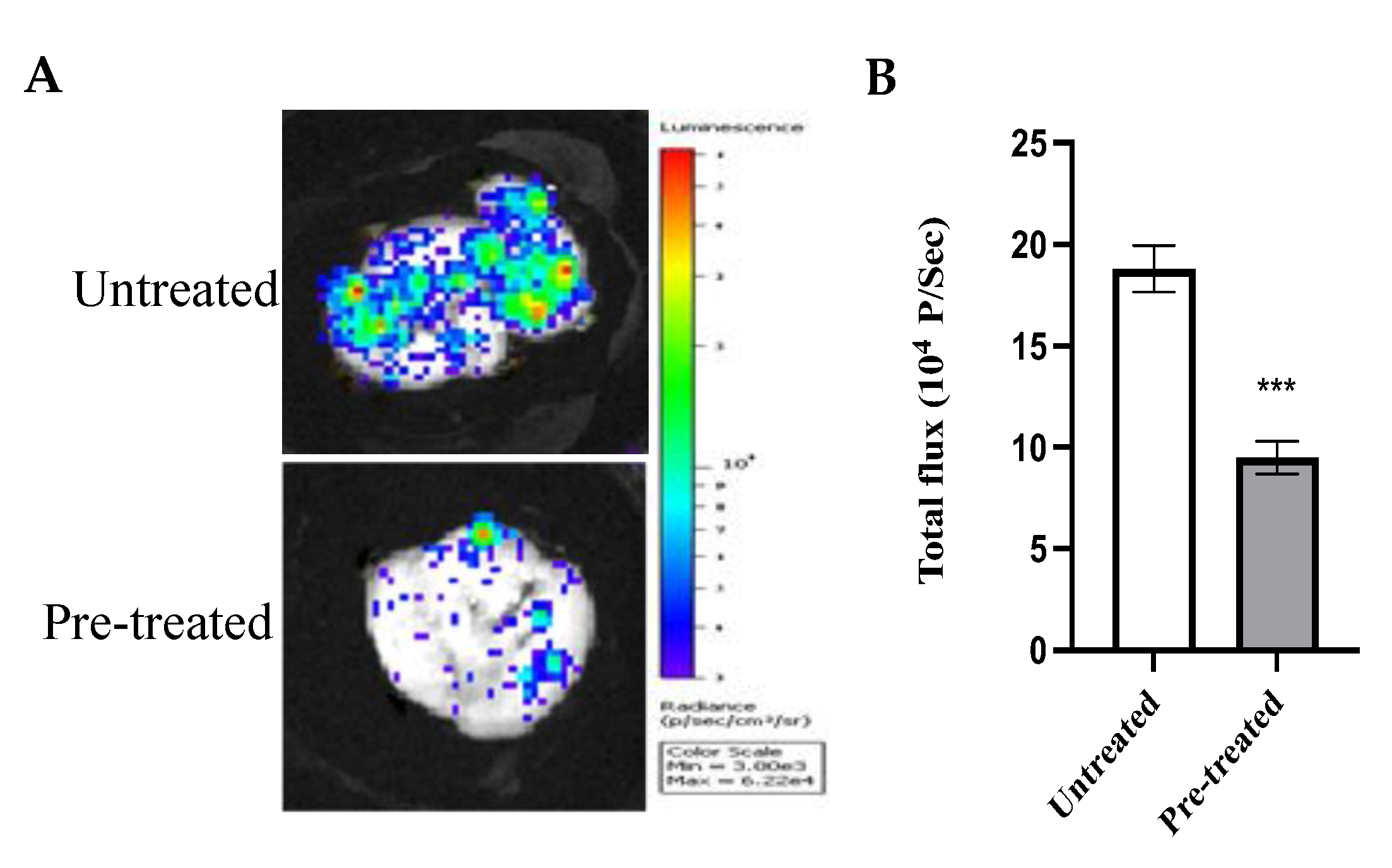
2.10. Effect of Azilsartan on the In Vivo Tumor Metastatic Activity
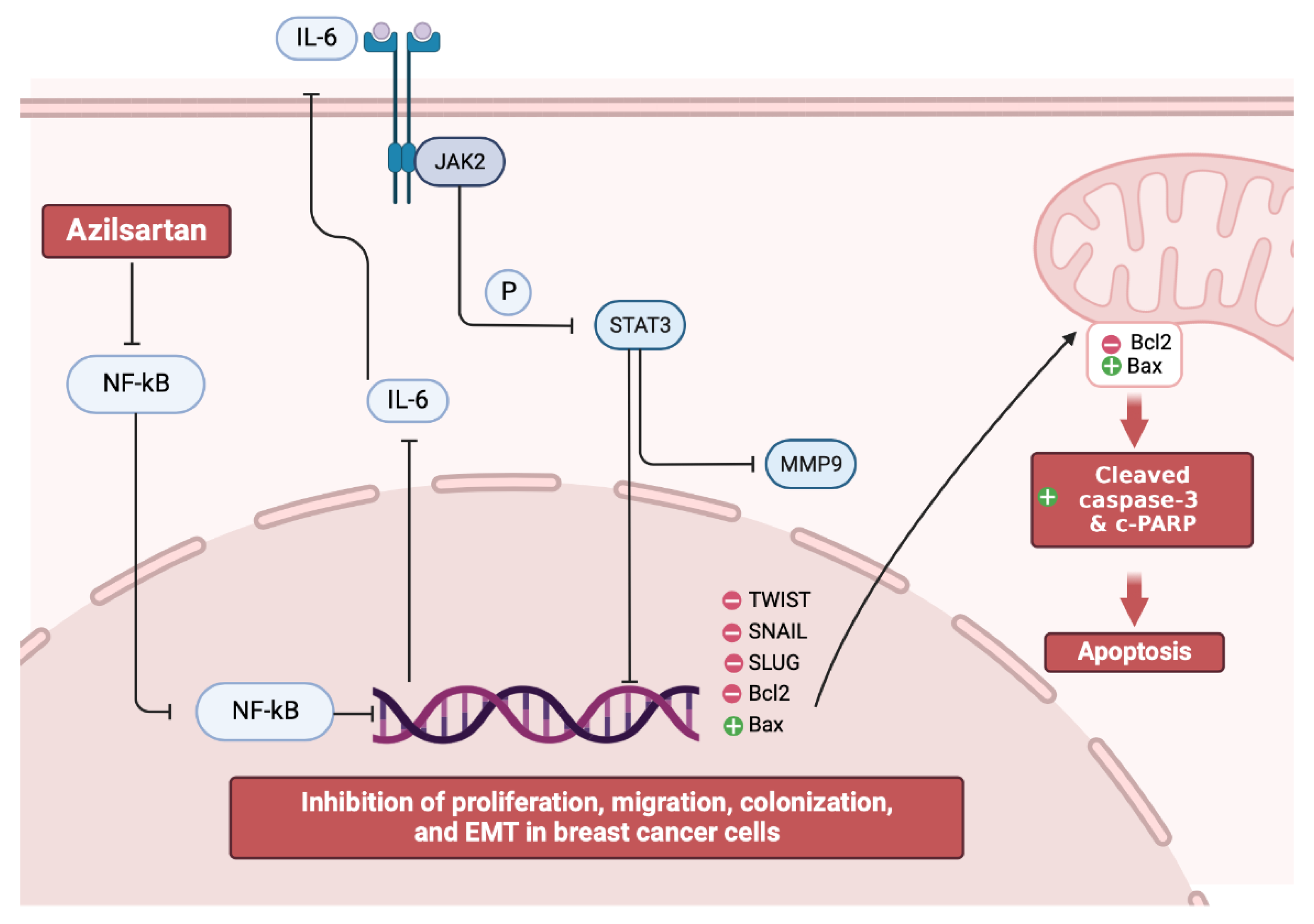
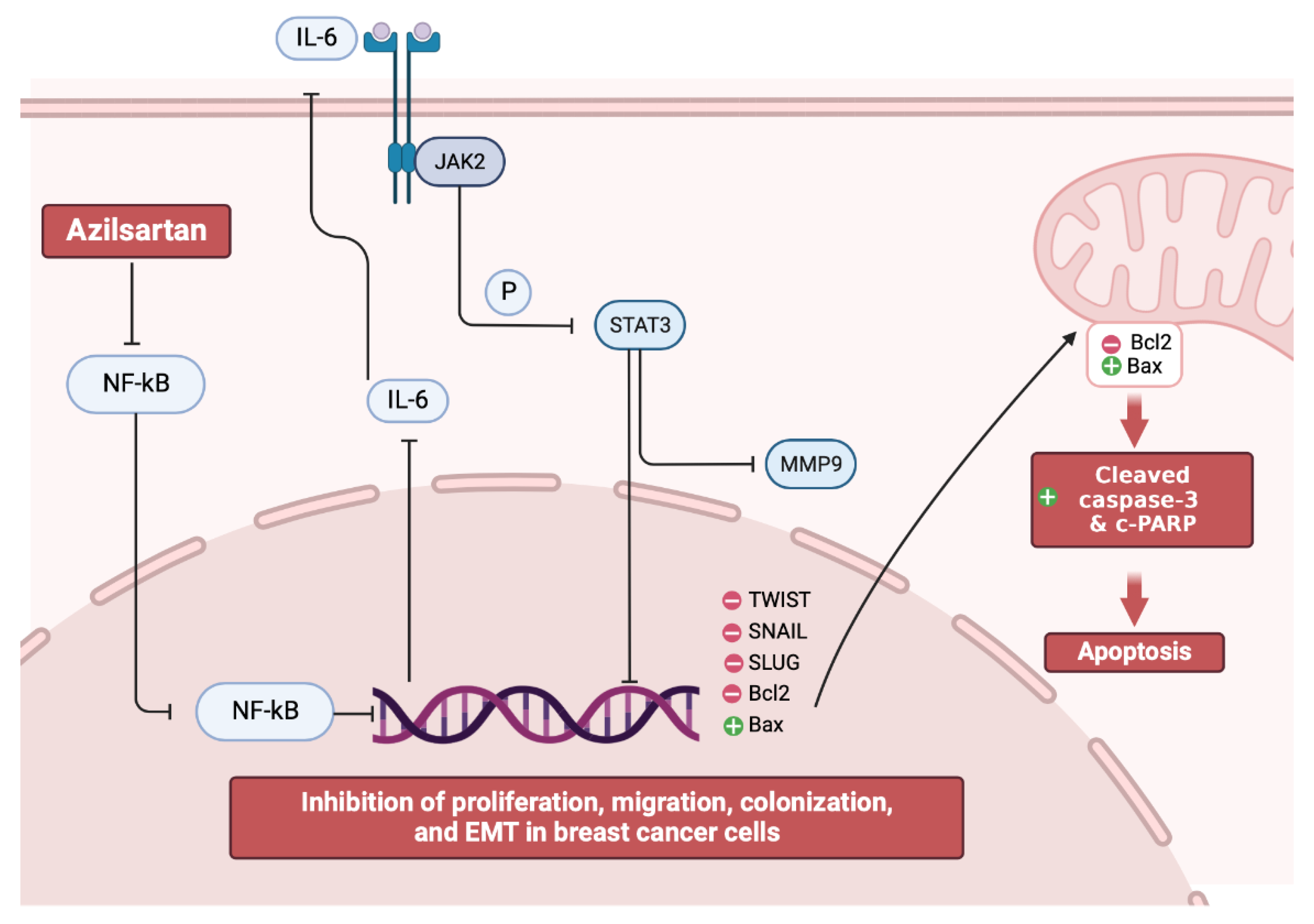
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability
4.3. Annexin V Assay
4.4. Cell Cycle Analysis
4.5. Colony Formation Assay
4.6. Cell Migration Assay
4.7. Measurement the Levels of Bax, Bcl2, Cleaved PARP Proteins
4.8. RNA Isolation and Real-Time PCR Assay
4.9. Analysis of Protein Expression via Western Blotting
4.10. Evaluation of Azilsartan Activity on TNFα-Induced NF-kB Activation in MCF-7 and MDA-MB-231 Cancer Cell Lines
4.11. The In Vivo Tumor Metastasis Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilkinson, L.; Gathani, T. Understanding breast cancer as a global health concern. Br. J. Radiol. 2021, 95, 20211033. [Google Scholar] [CrossRef]
- Nazmy, M.H.; Abu-baih, D.H.; El-Rehany, M.A.-A.; Fathy, M. Pathways of triple negative breast cancer. Minia J. Med. Res. 2021, 32, 1–3. [Google Scholar] [CrossRef]
- Gámez-Pozo, A.; Trilla-Fuertes, L.; Berges-Soria, J.; Selevsek, N.; López-Vacas, R.; Díaz-Almirón, M.; Nanni, P.; Arevalillo, J.M.; Navarro, H.; Grossmann, J.; et al. Functional proteomics outlines the complexity of breast cancer molecular subtypes. Sci. Rep. 2017, 7, 10100. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Yoshida, T.; Okabe, M.; Fathy, M.; Sun, Y.; Koike, C.; Salto, S.; Nikaido, T. CD24+SSEA4+cells in Ovarian Carcinoma Cells Demonstrated the Characteristics as Cancer Stem Cells. J. Cancer Sci. Ther. 2017, 9, 343–352. [Google Scholar] [CrossRef]
- Esteva, F.J.; Hubbard-Lucey, V.M.; Tang, J.; Pusztai, L. Immunotherapy and targeted therapy combinations in metastatic breast cancer. Lancet Oncol. 2019, 20, e175–e186. [Google Scholar] [CrossRef]
- Ahmadian, E.; Khosroushahi, A.Y.; Eftekhari, A.; Farajnia, S.; Babaei, H.; Eghbal, M.A. Novel angiotensin receptor blocker, azilsartan induces oxidative stress and NFkB-mediated apoptosis in hepatocellular carcinoma cell line HepG2. Biomed. Pharmacother. 2018, 99, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Alaaeldin, R.; Nazmy, M.H.; Abdel-Aziz, M.; Abuo-Rahma, G.E.-D.A.; Fathy, M. Cell Cycle Arrest and Apoptotic Effect of 7-(4-(N-substituted carbamoylmethyl) piperazin-1-yl) Ciprofloxacin-derivative on HCT 116 and A549 Cancer Cells. Anticancer Res. 2020, 40, 2739–2749. [Google Scholar] [CrossRef]
- Deshayes, F.; Nahmias, C. Angiotensin receptors: A new role in cancer? Trends Endocrinol. Metab. 2005, 16, 293–299. [Google Scholar] [CrossRef]
- Naugler, W.E.; Karin, M. NF-κB and cancer—identifying targets and mechanisms. Curr. Opin. Genet. Dev. 2008, 18, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Ferreira, S.; Nahmias, C. G-protein coupled receptors of the renin-angiotensin system: New targets against breast cancer? Front. Pharmacol. 2015, 6, 24. [Google Scholar] [CrossRef]
- Nagai, N.; Izumi-Nagai, K.; Oike, Y.; Koto, T.; Satofuka, S.; Ozawa, Y.; Yamashiro, K.; Inoue, M.; Tsubota, K.; Umezawa, K. Suppression of diabetes-induced retinal inflammation by blocking the angiotensin II type 1 receptor or its downstream nuclear factor-κB pathway. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4342–4350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdellatef, A.A.; Fathy, M.; Mohammed, A.E.I.; Bakr, M.S.A.; Ahmed, A.H.; Abbass, H.S.; El-Desoky, A.H.; Morita, H.; Nikaido, T.; Hayakawa, Y. Inhibition of cell-intrinsic NF-kappaB activity and metastatic abilities of breast cancer by aloe-emodin and emodic-acid isolated from Asphodelus microcarpus. J. Nat. Med. 2021, 75, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Alaaeldin, R.; Abdel-Rahman, I.A.; Hassan, H.A.; Youssef, N.; Allam, A.E.; Abdelwahab, S.F.; Zhao, Q.-L.; Fathy, M. Carpachromene Ameliorates Insulin Resistance in HepG2 Cells via Modulating IR/IRS1/PI3k/Akt/GSK3/FoxO1 Pathway. Molecules 2021, 26, 7629. [Google Scholar] [CrossRef] [PubMed]
- Alaaeldin, R.; Hassan, H.A.; Abdel-Rahman, I.M.; Mohyeldin, R.H.; Youssef, N.; Allam, A.E.; Abdelwahab, S.F.; Zhao, Q.-L.; Fathy, M. A New EGFR Inhibitor from Ficus benghalensis Exerted Potential Anti-Inflammatory Activity via Akt/PI3K Pathway Inhibition. Curr. Issues Mol. Biol. 2022, 44, 2967–2981. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Baky, R.M.; Hetta, H.F.; Fawzy, M.A.; Fathy, M. The association of interleukin 6 single nucleotide polymorphism with the susceptibility of Egyptians to HCV infection. Minia J. Med. Res. 2020, 31, 112–118. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Maher, S.A.; Bakkar, S.M.; El-Rehany, M.A.; Fathy, M. Pantoprazole Attenuates MAPK (ERK1/2, JNK, p38)-NF-kappaB and Apoptosis Signaling Pathways after Renal Ischemia/Reperfusion Injury in Rats. Int. J. Mol. Sci. 2021, 22, 10669. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Maher, S.A.; El-Rehany, M.A.; Welson, N.N.; Albezrah, N.K.A.; Batiha, G.E.-S.; Fathy, M. Vincamine Modulates the Effect of Pantoprazole in Renal Ischemia/Reperfusion Injury by Attenuating MAPK and Apoptosis Signaling Pathways. Molecules 2022, 27, 1383. [Google Scholar] [CrossRef]
- Ma, J.-h.; Qin, L.; Li, X. Role of STAT3 signaling pathway in breast cancer. Cell Commun. Signal. 2020, 18, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Balko, J.; Schwarz, L.; Luo, N.; Estrada, M.; Giltnane, J.; Dávila-González, D.; Wang, K.; Sánchez, V.; Dean, P.; Combs, S. Triple-negative breast cancers with amplification of JAK2 at the 9p24 locus demonstrate JAK2-specific dependence. Sci. Transl. Med. 2016, 8, 334ra53. [Google Scholar] [CrossRef] [Green Version]
- Fathy, M.; Nikaido, T. In vivo modulation of iNOS pathway in hepatocellular carcinoma by Nigella sativa. Environ. Health Prev. Med. 2013, 18, 377–385. [Google Scholar] [CrossRef]
- Fathy, M.; Nikaido, T. In vivo attenuation of angiogenesis in hepatocellular carcinoma by Nigella sativa. Turk. J. Med. Sci. 2018, 48, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Naseem, M.; Othman, E.M.; Fathy, M.; Iqbal, J.; Howari, F.M.; AlRemeithi, F.A.; Kodandaraman, G.; Stopper, H.; Bencurova, E.; Vlachakis, D.; et al. Integrated structural and functional analysis of the protective effects of kinetin against oxidative stress in mammalian cellular systems. Sci. Rep. 2020, 10, 13330. [Google Scholar] [CrossRef]
- Eisa, M.A.; Fathy, M.; Abuo-Rahma, G.; Abdel-Aziz, M.; Nazmy, M.H. Anti-Proliferative and Pro-Apoptotic Activities of Synthesized 3,4,5 Tri-Methoxy Ciprofloxacin Chalcone Hybrid, through p53 Up-Regulation in HepG2 and MCF7 Cell Lines. Asian Pac. J. Cancer Prev. 2021, 22, 3393–3404. [Google Scholar] [CrossRef]
- Fathy, M.; Sun, S.; Zhao, Q.-L.; Abdel-Aziz, M.; Abuo-Rahma, G.E.-D.A.; Awale, S.; Nikaido, T. A New Ciprofloxacin-derivative Inhibits Proliferation and Suppresses the Migration Ability of HeLa Cells. Anticancer Res. 2020, 40, 5025–5033. [Google Scholar] [CrossRef] [PubMed]
- Shytaj, I.L.; Fares, M.; Gallucci, L.; Lucic, B.; Tolba, M.M.; Zimmermann, L.; Adler, J.M.; Xing, N.; Bushe, J.; Gruber, A.D.; et al. The FDA-Approved Drug Cobicistat Synergizes with Remdesivir To Inhibit SARS-CoV-2 Replication In Vitro and Decreases Viral Titers and Disease Progression in Syrian Hamsters. mBio 2022, 13, e0370521. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Saad Eldin, S.M.; Naseem, M.; Dandekar, T.; Othman, E.M. Cytokinins: Wide-Spread Signaling Hormones from Plants to Humans with High Medical Potential. Nutrients 2022, 14, 1495. [Google Scholar] [CrossRef]
- Oba, J.; Okabe, M.; Yoshida, T.; Soko, C.; Fathy, M.; Amano, K.; Kobashi, D.; Wakasugi, M.; Okudera, H. Hyperdry human amniotic membrane application as a wound dressing for a full-thickness skin excision after a third-degree burn injury. Burn. Trauma 2020, 8, tkaa014. [Google Scholar] [CrossRef]
- Okabe, M.; Yoshida, T.; Suzuki, M.; Goto, M.; Omori, M.; Taguchi, M.; Toda, A.; Suzuki, T.; Nakagawa, K.; Hiramoto, F.; et al. Hyperdry Human Amniotic Membrane (HD-AM) is Supporting Aciclovir Included Device of Poly-N-p-Vinylbenzyl-D-Lactonamide (PVLA) Sphere for Treatment of HSV-1 Infected Rabbit Keratitis Model. J. Biotechnol. Biomater. 2017, 7, 251. [Google Scholar] [CrossRef] [Green Version]
- Othman, E.M.; Fathy, M.; Bekhit, A.A.; Abdel-Razik, A.H.; Jamal, A.; Nazzal, Y.; Shams, S.; Dandekar, T.; Naseem, M. Modulatory and Toxicological Perspectives on the Effects of the Small Molecule Kinetin. Molecules 2021, 26, 670. [Google Scholar] [CrossRef]
- Lam, S. Azilsartan: A newly approved angiotensin II receptor blocker. Cardiol Rev. 2011, 19, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Bidard, F.-C.; Pierga, J.-Y.; Vincent-Salomon, A.; Poupon, M.-F. A “class action” against the microenvironment: Do cancer cells cooperate in metastasis? Cancer Metastasis Rev. 2008, 27, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabra, R.T.; Abdellatef, A.A.; Abdel-Sattar, E.; Fathy, M.; Meselhy, M.R.; Hayakawa, Y. Russelioside A, a Pregnane Glycoside from Caralluma tuberculate, Inhibits Cell-Intrinsic NF-κB Activity and Metastatic Ability of Breast Cancer Cells. Biol. Pharm. Bull. 2022, 45, 1564–1571. [Google Scholar] [CrossRef]
- Rodrigues-Ferreira, S.; Abdelkarim, M.; Dillenburg-Pilla, P.; Luissint, A.-C.; di-Tommaso, A.; Deshayes, F.; Pontes, C.L.S.; Molina, A.; Cagnard, N.; Letourneur, F. Angiotensin II facilitates breast cancer cell migration and metastasis. PloS One 2012, 7, e35667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Sarkissyan, M.; Vadgama, J.V. Epithelial-mesenchymal transition and breast cancer. J. Clin. Med. 2016, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldafashi, N.; Darlay, R.; Shukla, R.; McCain, M.V.; Watson, R.; Liu, Y.L.; McStraw, N.; Fathy, M.; Fawzy, M.A.; Zaki, M.Y.W.; et al. A PDCD1 Role in the Genetic Predisposition to NAFLD-HCC? Cancers 2021, 13, 1412. [Google Scholar] [CrossRef]
- Fathy, M.; Awale, S.; Nikaido, T. Phosphorylated Akt Protein at Ser473 Enables HeLa Cells to Tolerate Nutrient-Deprived Conditions. Asian Pac. J. Cancer Prev. 2017, 18, 3255–3260. [Google Scholar] [CrossRef]
- Thiery, J.P.; Lim, C.T. Tumor dissemination: An EMT affair. Cancer Cell 2013, 23, 272–273. [Google Scholar] [CrossRef] [Green Version]
- Bill, R.; Christofori, G. The relevance of EMT in breast cancer metastasis: Correlation or causality? FEBS Lett. 2015, 589, 1577–1587. [Google Scholar] [CrossRef] [Green Version]
- George, A.J.; Thomas, W.G.; Hannan, R.D. The renin–angiotensin system and cancer: Old dog, new tricks. Nat. Rev. Cancer 2010, 10, 745–759. [Google Scholar] [CrossRef]
- Fathy, M.; Khalifa, E.; Fawzy, M.A. Modulation of inducible nitric oxide synthase pathway by eugenol and telmisartan in carbon tetrachloride-induced liver injury in rats. Life Sci. 2019, 216, 207–214. [Google Scholar] [CrossRef]
- Lin, W.-H.; Dai, W.-G.; Xu, X.-D.; Yu, Q.-H.; Zhang, B.; Li, J.; Li, H.-P. Downregulation of DPF3 promotes the proliferation and motility of breast cancer cells through activating JAK2/STAT3 signaling. Biochem. Biophys. Res. Commun. 2019, 514, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.Z.; Patil, P.; Gude, R.P. Role of STAT3 in Cancer Metastasis and Translational Advances. BioMed Res. Int. 2013, 2013, 421821. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Abdel-latif, R.; Abdelgwad, Y.M.; Othman, O.A.; Abdel-Razik, A.-R.H.; Dandekar, T.; Othman, E.M. Nephroprotective potential of eugenol in a rat experimental model of chronic kidney injury; targeting NOX, TGF-β, and Akt signaling. Life Sci. 2022, 308, 120957. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Darwish, M.A.; Abdelhamid, A.M.; Alrashedy, G.M.; Othman, O.A.; Naseem, M.; Dandekar, T.; Othman, E.M. Kinetin Ameliorates Cisplatin-Induced Hepatotoxicity and Lymphotoxicity via Attenuating Oxidative Damage, Cell Apoptosis and Inflammation in Rats. Biomedicines 2022, 10, 1620. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Okabe, M.; Othman, E.M.; Saad Eldien, H.M.; Yoshida, T. Preconditioning of Adipose-Derived Mesenchymal Stem-Like Cells with Eugenol Potentiates Their Migration and Proliferation In Vitro and Therapeutic Abilities in Rat Hepatic Fibrosis. Molecules 2020, 25, 2020. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Okabe, M.; Saad Eldien, H.M.; Yoshida, T. AT-MSCs Antifibrotic Activity is Improved by Eugenol through Modulation of TGF-beta/Smad Signaling Pathway in Rats. Molecules 2020, 25, 348. [Google Scholar] [CrossRef] [Green Version]
- Neutel, J.M.; Cushman, W.C.; Lloyd, E.; Barger, B.; Handley, A. Comparison of long-term safety of fixed-dose combinations azilsartan medoxomil/chlorthalidone vs olmesartan medoxomil/hydrochlorothiazide. J. Clin. Hypertens. 2017, 19, 874–883. [Google Scholar] [CrossRef] [Green Version]
- Hardin, M.D.; Jacobs, T.F. Azilsartan; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Hama Amin, R.R.; Aziz, T.A. Gastroprotective Effect of Azilsartan Through Ameliorating Oxidative Stress, Inflammation, and Restoring Hydroxyproline, and Gastrin Levels in Ethanol-Induced Gastric Ulcer. J. Inflamm. Res. 2022, 15, 2911–2923. [Google Scholar] [CrossRef]
- Martinez, V.R.; Aguirre, M.V.; Todaro, J.S.; Piro, O.E.; Echeverria, G.A.; Ferrer, E.G.; Williams, P.A.M. Azilsartan and its Zn(II) complex. Synthesis, anticancer mechanisms of action and binding to bovine serum albumin. Toxicol Vitr. 2018, 48, 205–220. [Google Scholar] [CrossRef]
- Alaaeldin, R.; Mustafa, M.; Abuo-Rahma, G.E.A.; Fathy, M. In vitro inhibition and molecular docking of a new ciprofloxacin chalcone against SARS-CoV-2 main protease. Fundam. Clin. Pharm. 2022, 36, 160–170. [Google Scholar] [CrossRef]
- Zhang, F.; Yin, G.; Han, X.; Jiang, X.; Bao, Z. Chlorogenic acid inhibits osteosarcoma carcinogenesis via suppressing the STAT3/Snail pathway. J. Cell. Biochem. 2019, 120, 10342–10350. [Google Scholar] [CrossRef]
- Moirangthem, A.; Bondhopadhyay, B.; Mukherjee, M.; Bandyopadhyay, A.; Mukherjee, N.; Konar, K.; Bhattacharya, S.; Basu, A. Simultaneous knockdown of uPA and MMP9 can reduce breast cancer progression by increasing cell-cell adhesion and modulating EMT genes. Sci. Rep. 2016, 6, 21903. [Google Scholar] [CrossRef] [Green Version]
- Alaaeldin, R.; Abuo-Rahma, G.E.-D.A.; Zhao, Q.-L.; Fathy, M. Modulation of apoptosis and epithelial-Mesenchymal transition E-cadherin/TGF-β/Snail/TWIST pathways by a new ciprofloxacin chalcone in breast cancer cells. Anticancer Res. 2021, 41, 2383–2395. [Google Scholar] [CrossRef]
- Alves, C.C.; Carneiro, F.; Hoefler, H.; Becker, K.-F. Role of the epithelial-mesenchymal transition regulator Slug in primary human cancers. Front. Biosci.-Landmark 2009, 14, 3035–3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Bae, S.; An, S.; Park, J.K.; Kim, E.M.; Hwang, S.G.; Kim, W.J.; Um, H.D. Cooperative actions of p21 WAF 1 and p53 induce Slug protein degradation and suppress cell invasion. EMBO Rep. 2014, 15, 1062–1068. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, N.; Fathy, M.; Amgad, S.W. Glycoregulatory Enzymes as Early Diagnostic Markers during Premalignant Stage in Hepatocellular Carcinoma. Am. J. Cancer Prev. 2013, 1, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Noto, Z.; Yoshida, T.; Okabe, M.; Koike, C.; Fathy, M.; Tsuno, H.; Tomihara, K.; Arai, N.; Noguchi, M.; Nikaido, T. CD44 and SSEA-4 positive cells in an oral cancer cell line HSC-4 possess cancer stem-like cell characteristics. Oral Oncol. 2013, 49, 787–795. [Google Scholar] [CrossRef]
- Puisieux, A.; Valsesia-Wittmann, S.; Ansieau, S. A twist for survival and cancer progression. Br. J. Cancer 2006, 94, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Shi, J.; Chai, K.; Ying, X.; Zhou, B.P. The Role of Snail in EMT and Tumorigenesis. Curr. Cancer Drug Targets 2013, 13, 963–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Yang, Z.; Huang, X.; Zhang, Z.; Li, J.; Ju, J.; Zhang, H.; Ma, J. Ilamycin C induces apoptosis and inhibits migration and invasion in triple-negative breast cancer by suppressing IL-6/STAT3 pathway. J. Hematol. Oncol. 2019, 12, 1–14. [Google Scholar] [CrossRef]
- Abd-Elhamid, R.A.; Nazmy, M.H.; Fathy, M. Targeting Apoptosis as a Therapeutic Approach in Cancer. Minia J. Med. Res. 2020, 31, 321–334. [Google Scholar] [CrossRef]
- Eisa, M.A.; Fathy, M.; Nazmy, M.H. Potential COX2 mediated therapeutic effect of ciprofloxacin. Minia J. Med. Res. 2021, 32, 47–57. [Google Scholar] [CrossRef]
- Fathy, M.; Fawzy, M.A.; Hintzsche, H.; Nikaido, T.; Dandekar, T.; Othman, E.M. Eugenol exerts apoptotic effect and modulates the sensitivity of HeLa cells to cisplatin and radiation. Molecules 2019, 24, 3979. [Google Scholar] [CrossRef] [Green Version]
- Wolczyk, D.; Zaremba-Czogalla, M.; Hryniewicz-Jankowska, A.; Tabola, R.; Grabowski, K.; Sikorski, A.F.; Augoff, K. TNF-α promotes breast cancer cell migration and enhances the concentration of membrane-associated proteases in lipid rafts. Cell. Oncol. 2016, 39, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Goel, A.; Prasad, A.K.; Parmar, V.S.; Ghosh, B.; Saini, N. Apoptogenic effect of 7,8-diacetoxy-4-methylcoumarin and 7,8-diacetoxy-4-methylthiocoumarin in human lung adenocarcinoma cell line: Role of NF-kappaB, Akt, ROS and MAP kinase pathway. Chem. Biol. Interact. 2009, 179, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Kolli, S.; Bojic, S.; Ghareeb, A.E.; Kurzawa-Akanbi, M.; Figueiredo, F.C.; Lako, M. The Role of Nerve Growth Factor in Maintaining Proliferative Capacity, Colony-Forming Efficiency, and the Limbal Stem Cell Phenotype. Stem Cells 2019, 37, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Barber, R.D.; Harmer, D.W.; Coleman, R.A.; Clark, B.J. GAPDH as a housekeeping gene: Analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol Genom. 2005, 21, 389–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Greenfield, E.A. Antibodies: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2013. [Google Scholar]
- Davis, J.N.; Kucuk, O.; Sarkar, F.H. Genistein inhibits NF-kB activation in prostate cancer cells. Nutr. Cancer 1999, 35, 167–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| TNFα | Forward: 5′-CGCTTAGGAGGGAGAGCCCA-3′ Reverse: 5′-TGCCATTCTGAAGCCGGGTG-3′ |
| NF-kB | Forward: 5′-ATGTCCGCGTCCCACTAGCA-3′ Reverse: 5′-GCCCCACGCCCTGTTTCTTT-3′ |
| TWIST | Forward: 5′-CGGCCTAGCGAGTGGTTCTT-3′ Reverse: 5′-AGGAAAGAGCGCGGCATAGT-3′ |
| SNAIL | Forward:5′-ATCTGCGGCAAGGCGTTTTCCA-3′ Reverse: 5′-GAGCCCTCAGATTTGACCTGTC-3‘ |
| SLUG | Forward: 5′-GTTTCA TCC AGG ATC GAG CAG-3′ Reverse: 5′-CATCTT CTT CCA GAT GGT GA-3′ |
| bax | Forward: 5′-CCTGTG GAT GAC TGA GTA CC-3′ Reverse: 5′-GAGACA GCC AGG AGA AAT CA-3′ |
| Bcl2 | Forward: 5′-CCCAGAAGACAGTGGACGGG-3′ Reverse: 5′-CGACAGACACATCCGGGGTT-3′ |
| GAPDH | Forward: 5′-CGCTTAGGAGGGAGAGCCCA-3′ Reverse: 5′-TGCCATTCTGAAGCCGGGTG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alaaeldin, R.; Ali, F.E.M.; Bekhit, A.A.; Zhao, Q.-L.; Fathy, M. Inhibition of NF-kB/IL-6/JAK2/STAT3 Pathway and Epithelial-Mesenchymal Transition in Breast Cancer Cells by Azilsartan. Molecules 2022, 27, 7825. https://doi.org/10.3390/molecules27227825
Alaaeldin R, Ali FEM, Bekhit AA, Zhao Q-L, Fathy M. Inhibition of NF-kB/IL-6/JAK2/STAT3 Pathway and Epithelial-Mesenchymal Transition in Breast Cancer Cells by Azilsartan. Molecules. 2022; 27(22):7825. https://doi.org/10.3390/molecules27227825
Chicago/Turabian StyleAlaaeldin, Rania, Fares E. M. Ali, Amany Abdlrehim Bekhit, Qing-Li Zhao, and Moustafa Fathy. 2022. "Inhibition of NF-kB/IL-6/JAK2/STAT3 Pathway and Epithelial-Mesenchymal Transition in Breast Cancer Cells by Azilsartan" Molecules 27, no. 22: 7825. https://doi.org/10.3390/molecules27227825
APA StyleAlaaeldin, R., Ali, F. E. M., Bekhit, A. A., Zhao, Q. -L., & Fathy, M. (2022). Inhibition of NF-kB/IL-6/JAK2/STAT3 Pathway and Epithelial-Mesenchymal Transition in Breast Cancer Cells by Azilsartan. Molecules, 27(22), 7825. https://doi.org/10.3390/molecules27227825