
Tritordeum as an Innovative Alternative to Wheat: A Comparative Digestion Study on Bread

 , , ,
, , ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
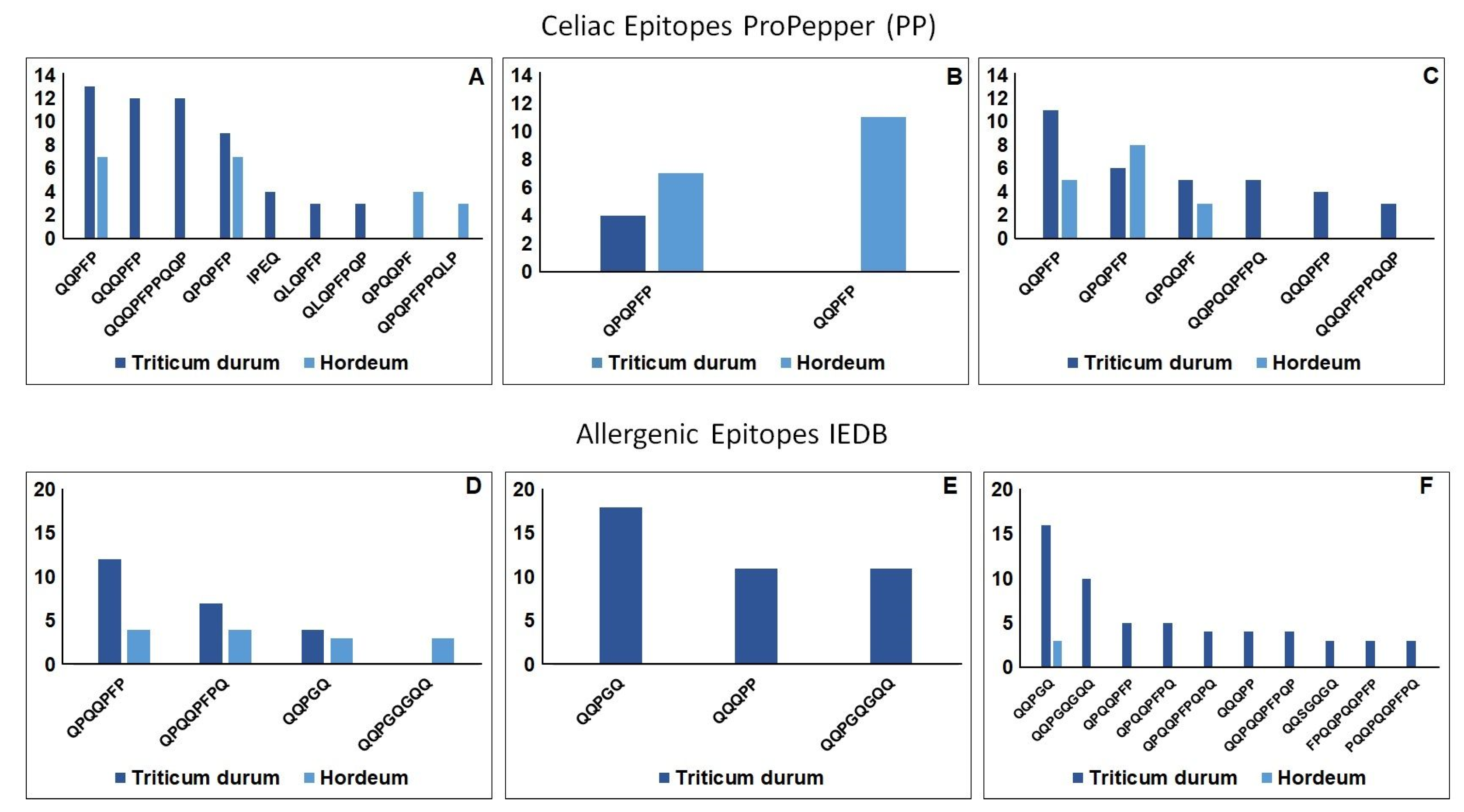
2. Results and Discussion
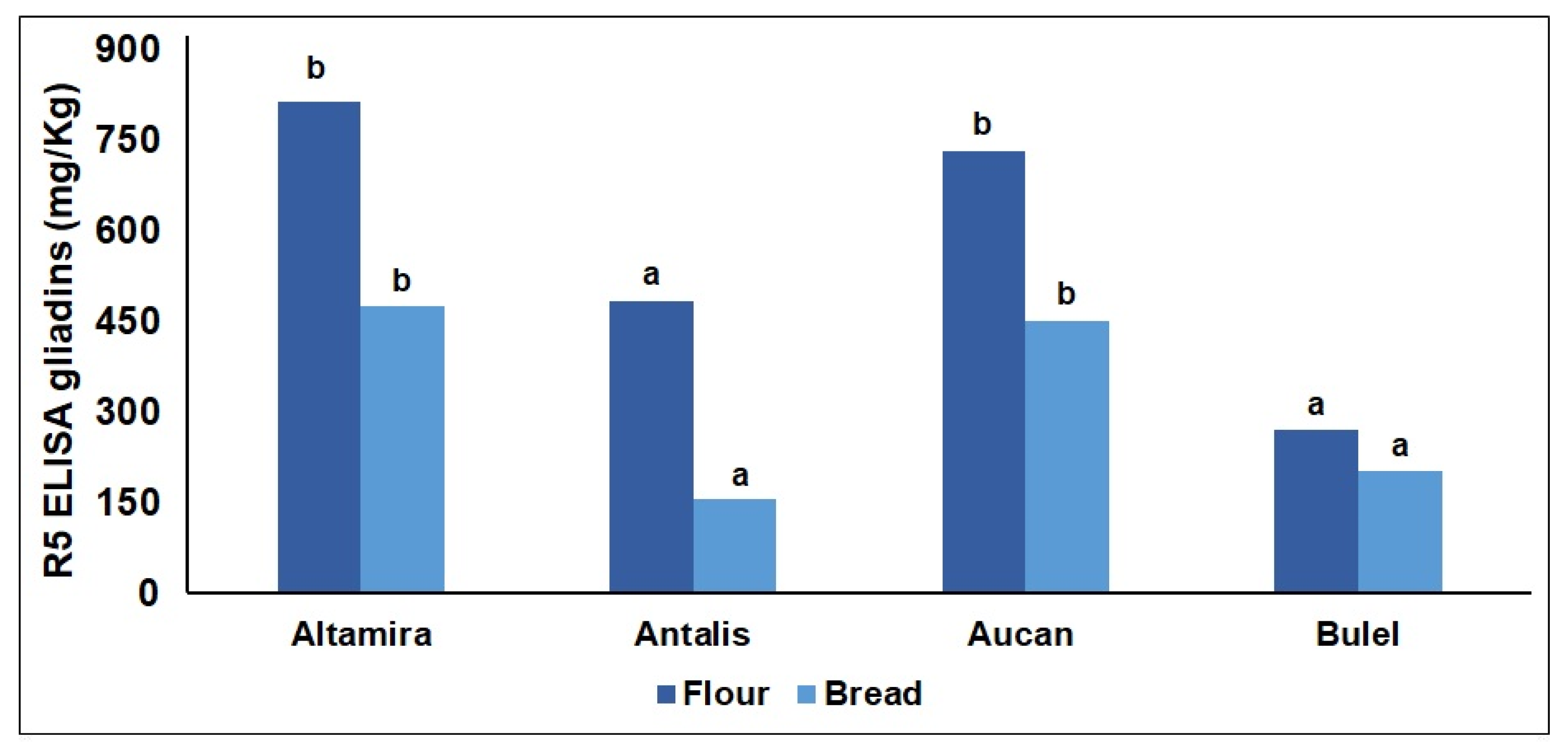
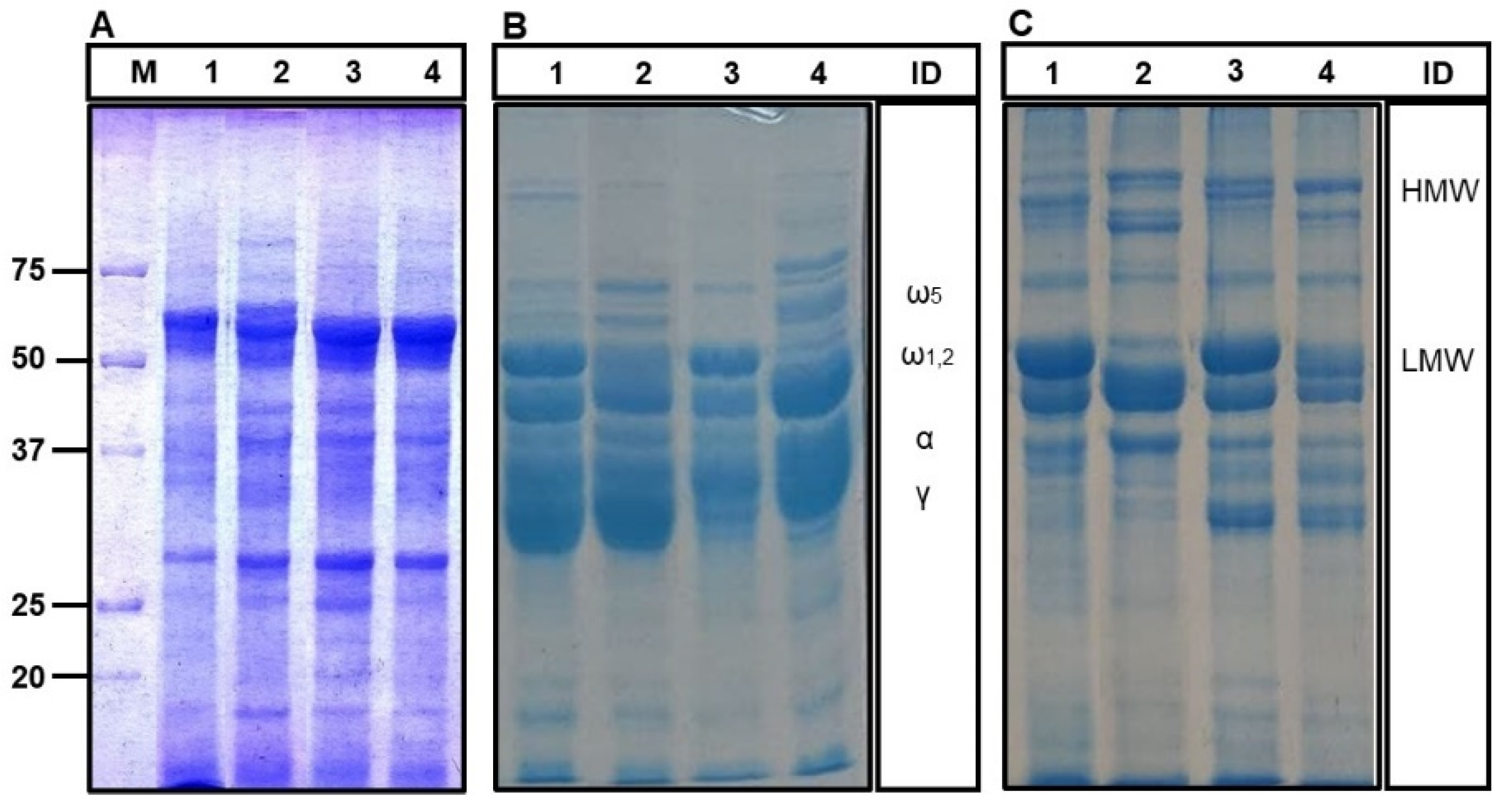
2.1. Flour Protein Characterization
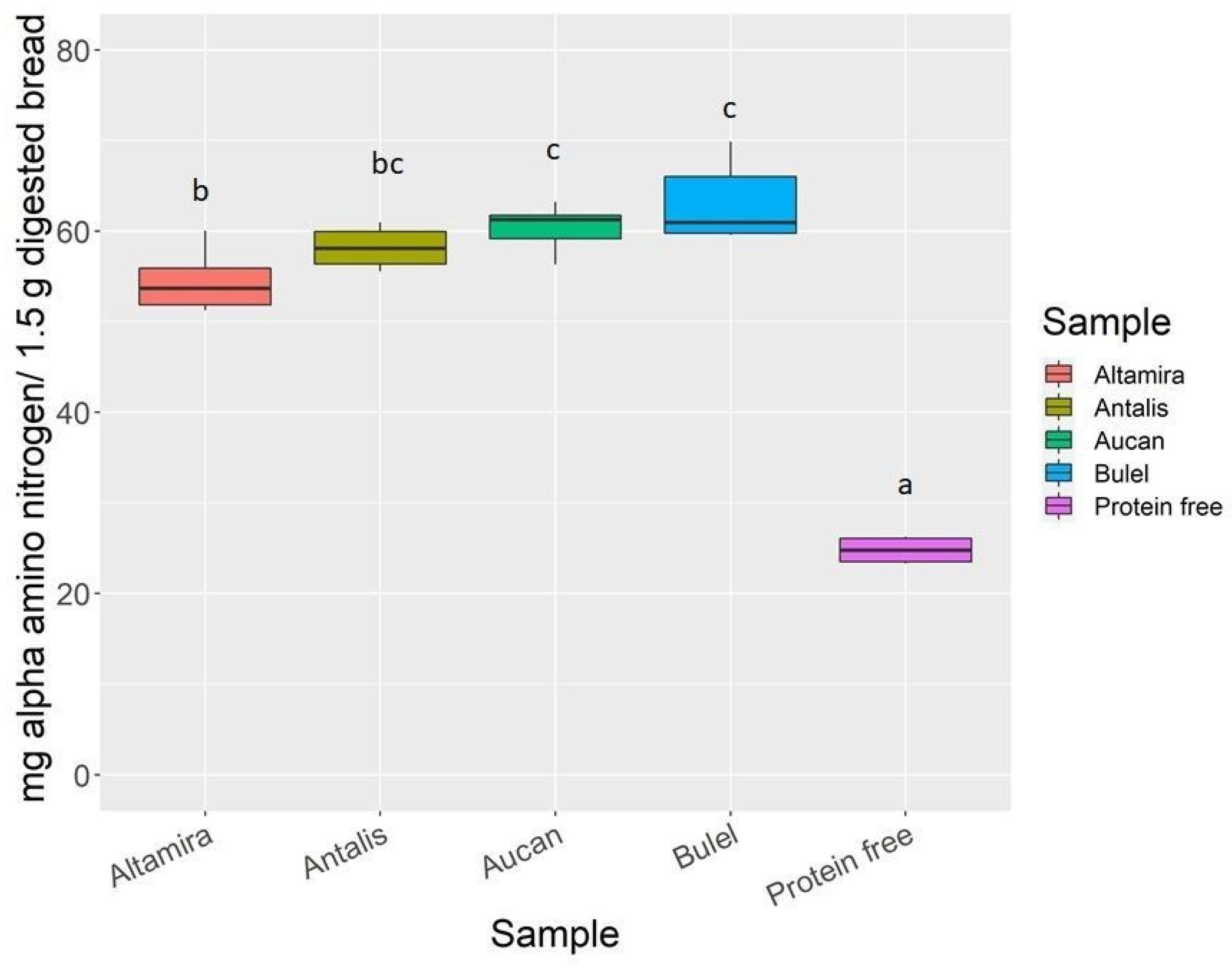
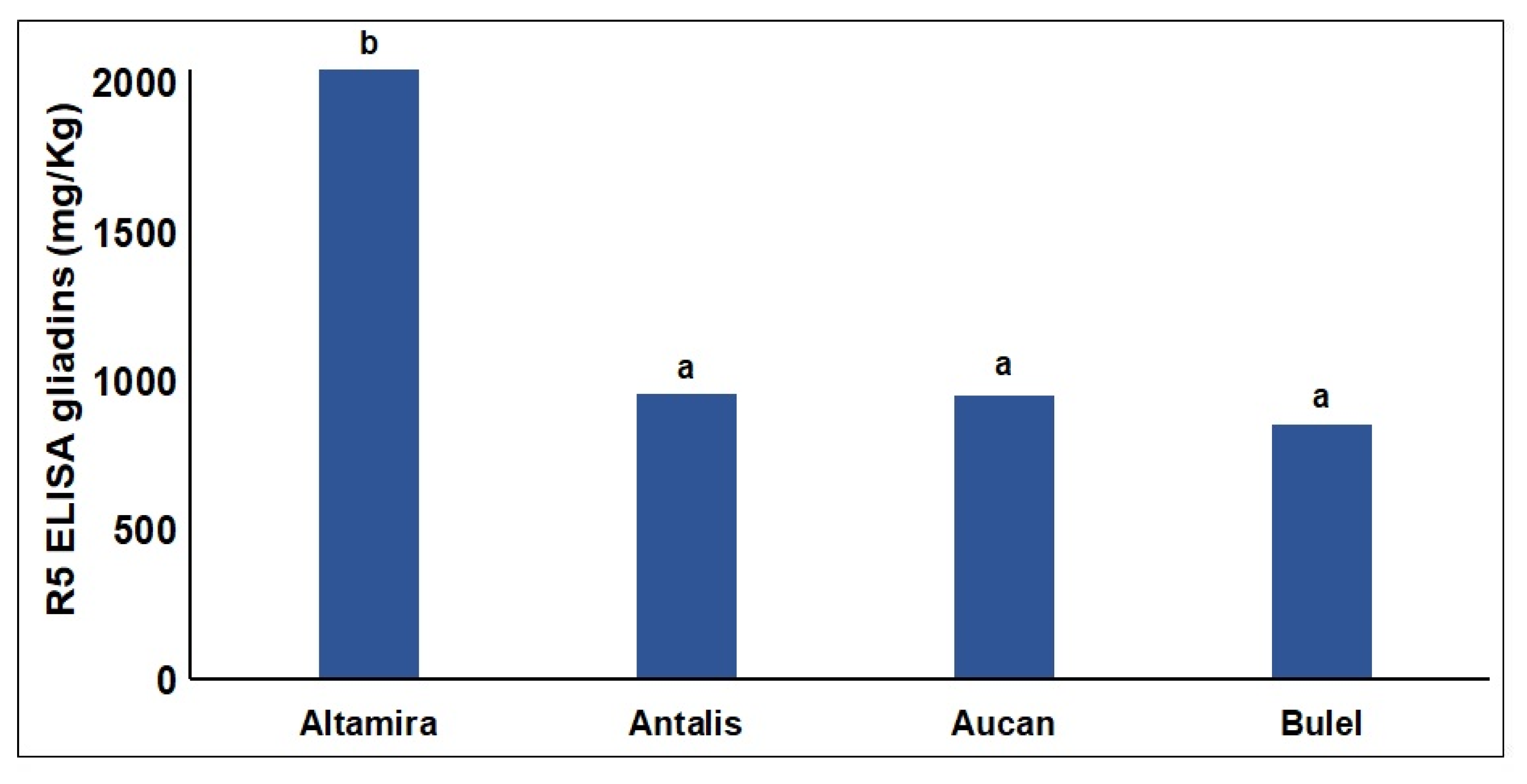
2.2. Digestomics
2.2.1. Quantitative Analysis of the End Products of Digestion
2.2.2. Qualitative Evaluation of the Peptides Resistant to Digestion
3. Materials and Methods
3.1. Grains and Flours Production
- A soft wheat (hexaploid AABBDD), cv named Altamira (seeds provided by Limagrain Italia S.p.A., Busseto, Italy) classified as ordinary bread-making wheat [31] registered in the Italian varietal list in 2009 (https://www.sian.it/mivmPubb/listeVarieta.do; Sian code: 11239; consulted on the 20 December 2021) and widely cultivated in Italy;
- A durum wheat (tetraploid AABB), cv named Antalis (seeds provided by CGS Sementi S.p.A., Acquasparta, Italy), characterized by medium-high GPC and gluten index; registered in the Italian varietal list in 2014 and widely cultivated in Italy;
- Tritordeum (hexaploid AABBHchHch), cv named Bulel (seeds provided by Arcadia S.p.A., Pamplona, Spain), which was registered in the CPVO (Community Plant Variety Office) List in 2015;
- Tritordeum (hexaploid AABBHchHch), cv named Aucan (seeds provided by Arcadia S.p.A., Pamplona, Spain), which was registered in the CPVO List in 2013.
3.2. Materials
3.3. Quantification of Protein in Flour and Semolina
3.4. Gliadin Quantification with R5 Commercial ELISA
3.5. Osborne Fractionation
3.6. 1-Dimensional Electrophoresis (SDS-PAGE)
3.7. Preparation of the Model Breads
3.8. Static Oral-Gastric-Duodenal Digestion of Model Breads
3.9. Preparation of Samples for the Alpha Amino Nitrogen Determination
3.10. Free Glucose Quantitative Determination
3.11. Preparation of Peptides for Mass Spectrometry Analysis
3.12. Liquid Chromatography-Tandem Mass Spectrometry (LC/MSMS) Analysis
3.13. In Silico Analysis of Peptides Resistant to Digestion
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| AAI | alpha amylase inhibitors |
| ACN | acetonitrile |
| AGC | automatic gain control |
| AmBic | ammonium bicarbonate |
| ANOVA | analysis of variance |
| cv | cultivar |
| DTT | 1,4-Dithio-D-threitol |
| ELISA | enzyme-linked immunosorbent assay |
| EDTA | ethylenediaminetetraacetic acid |
| FA | formic acid |
| FDR | false discovery rate |
| GPC | grain protein content |
| GS | growth stage |
| Hch | Hordeum chilense |
| HMW-GS | High-molecular-weight glutenin subunit |
| IEDB | Immune Epitope Database |
| IT | injection time |
| LC-MS/MS | liquid chromatograph-mass spectrometer/mass spectrometry |
| LMW-GS | Low-molecular-weight glutenin subunit |
| RSR | reducing sugar release |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SGF | simulated gastric fluid |
| SIF | simulated intestinal fluid |
| SSF | simulated salivary fluid |
| TAME | p-toluenesulfonyl-L-arginine methyl ester |
| TCA | trichloroacetic acid |
| TFA | trifluoroacetic acid |
| TPC | total protein content |
| Tris-HCl | tris(hydroxymethyl) aminomethane hydrochloride |
References
- Poole, N.; Donovan, J.; Erenstein, O. Viewpoint: Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy 2021, 100, 101976. [Google Scholar] [CrossRef] [PubMed]
- Örgeç, M.; Doğan, C.N.; Ağıl, F.; Şahin, G.; Zencirci, N. Introduction (Chapter 1). In Book Wheat Landraces, 1st ed.; Zencirci, N., Baloch, F.S., Habyarimana, E., Chung, G., Eds.; Springer Nature: Cham, Switzerland, 2021; pp. 1–12. [Google Scholar]
- Obembe, O.S.; Hendricks, N.P.; Tack, J. Decreased wheat production in the USA from climate change driven by yield losses rather than crop abandonment. PLoS ONE 2021, 16, e0252067. [Google Scholar] [CrossRef] [PubMed]
- Daloz, A.S.; Rydsaa, J.H.; Hodnebrog, Ø.; Sillmann, J.; van Oort, B.; Mohr, C.W.; Agrawal, M.; Emberson, L.; Stordal, F.; Zhang, T. Direct and indirect impacts of climate change on wheat yield in the Indo-Gangetic plain in India. J. Agric. Food Res. 2021, 4, 100132. [Google Scholar] [CrossRef]
- Bento, V.A.; Ribeiro, A.F.S.; Russo, A.; Gouveia, C.M.; Cardoso, R.M.; Soares, P.M.M. The impact of climate change in wheat and barley yields in the Iberian Peninsula. Sci. Rep. 2021, 11, 15484. [Google Scholar] [CrossRef] [PubMed]
- Sagnelli, D.; Chessa, S.; Mandalari, G.; Di Martino, M.; Sorndech, W.; Mamone, G.; Vincze, E.; Buillon, G.; Nielsen, D.S.; Wiese, M.; et al. Low glycaemic index foods from wild barley and amylose-only barley lines. J. Funct. Foods 2018, 40, 408–416. [Google Scholar] [CrossRef]
- Martín, A.; Cabrera, A.; Hernández, P.; Ramírez, M.C.; Rubiales, D.; Ballesteros, J. Prospect for the use of Hordeum chilense in durum wheat breeding. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Royo, C., Nachit, M., Di Fonzo, N., Araus, J.L., Eds.; CIHEAM (Options Méditerranéennes: Série A. Séminaires Méditerranéens; n. 40); Seminar on Durum Wheat Improvement in the Mediterranean Region: New Challenges, 12–14 April 2000; Instituto Agronómico Mediterráneo de Zaragoza: Foggia, Spain, 2002; pp. 111–115. [Google Scholar]
- Vaquero, L.; Comino, I.; Vivas, S.; Rodríguez-Martín, L.; Giménez, M.J.; Pastor, J.; Sousa, C.; Barro, F. Tritordeum: A novel cereal for food processing with good acceptability and significant reduction in gluten immunogenic peptides in comparison with wheat. J. Sci. Food Agric. 2018, 98, 2201–2209. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Ballesteros, J.; Sillero, J.A.; Martin, L.M. Tritordeum: A new crop of potential importance in the food industry. Hereditas 1992, 116, 193–197. [Google Scholar] [CrossRef]
- Martín, A.; Alvarez, J.B.; Martín, L.M.; Barro, F.; Ballesteros, J. The development of tritordeum: A novel cereal for food processing. J. Cereal Sci. 1999, 30, 85–95. [Google Scholar] [CrossRef]
- Sánchez-León, S.; Haro, C.; Villatoro, M.; Vaquero, L.; Comino, I.; González-Amigo, A.B.; Vivas, S.; Pastor, J.; Sousa, C.; Landa, B.B.; et al. Tritordeum breads are well tolerated with preference over gluten-free breads in non-celiac wheat-sensitive patients and its consumption induce changes in gut bacteria. J. Sci. Food Agric. 2021, 101, 3508–3517. [Google Scholar] [CrossRef]
- Kakabouki, I.; Beslemes, D.F.; Tigka, E.L.; Folina, A.; Karydogianni, S.; Zisi, C.; Papastylianou, P. Performance of six genotypes of tritordeum compare to bread wheat under east mediterranean condition. Sustainability 2020, 12, 9700. [Google Scholar] [CrossRef]
- Visioli, G.; Lauro, M.; Vamerali, T.; Dal Cortivo, C.; Panozzo, A.; Folloni, S.; Piazza, C.; Ranieri, R. A comparative study of organic and conventional management on the rhizosphere microbiome, growth and grain quality traits of tritordeum. Agronomy 2020, 10, 1717. [Google Scholar] [CrossRef]
- Giordano, D.; Reyneri, A.; Locatelli, M.; Coïsson, J.D.; Blandino, M. Distribution of bioactive compounds in pearled fractions of tritordeum. Food Chem. 2019, 301, 125228. [Google Scholar] [CrossRef]
- Mendis, M.; Simsek, S. Arabinoxylans and human health. Food Hydrocoll. 2014, 42, 239–243. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A critical review on the impacts of β-glucans on gut microbiota and human health. J. Nutr. Biochem. 2018, 61, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Kahlenberg, F.; Sanchez, D.; Lachmann, I.; Tuckova, L.; Tlaskalova, H.; Méndez, E.; Mothes, T. Monoclonal antibody R5 for detection of putatively coeliac-toxic gliadin peptides. Eur. Food Res. Technol. 2006, 222, 78–82. [Google Scholar] [CrossRef]
- Immer, U.; Haas-Lauterbach, S. Gluten Detection. In Molecular Biological and Immunological Techniques and Applications for Food Chemists; Popping, B., Diaz-Amigo, C., Hoenicke, K., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 359–376. ISBN 9780470637685. [Google Scholar]
- Landolfi, V.; D’Auria, G.; Nicolai, M.A.; Nitride, C.; Blandino, M.; Ferranti, P. The effect of nitrogen fertilization on the expression of protein in wheat and tritordeum varieties using a proteomic approach. Food Res. Int. 2021, 148, 110617. [Google Scholar] [CrossRef]
- Li, Y.; Fu, J.; Shen, Q.; Yang, D. High-Molecular-Weight Glutenin Subunits: Genetics, Structures, and Relation to End Use Qualities. Int. J. Mol. Sci. 2020, 22, 184. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Campos, L.A.C.; Martín, A.; Sillero, J.A.; Martín, L.M. Genetic analysis of prolamins synthesised by the H(ch) genome and their effects on gluten strength in hexaploid tritordeum. Euphytica 1999, 107, 177–184. [Google Scholar] [CrossRef]
- Tercero, J.A.; Bernardo, A.; Jouve, N. Encoding genes for endosperm proteins in Hordeum chilense. Theor. Appl. Genet. 1991, 81, 127–132. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Martín, A.; Martín, L.M. Variation in the high-molecular-weight glutenin subunits coded at the Glu-Hch1 locus in Hordeum chilense. Theor. Appl. Genet. 2001, 102, 134–137. [Google Scholar] [CrossRef]
- Mikulíková, D.; Benková, M.; Kraic, J. The Potential of Common Cereals to form Retrograded Resistant Starch. Czech. J. Genet. Plant. Breed. 2018, 42, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Daly, M.; Bromilow, S.N.; Nitride, C.; Shewry, P.R.; Gethings, L.A.; Mills, E.N.C. Mapping Coeliac Toxic Motifs in the Prolamin Seed Storage Proteins of Barley, Rye, and Oats Using a Curated Sequence Database. Front. Nutr. 2020, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.Q.T.; Wang, S.; Kumar, A.; Yap, L.J.; Luu, T.T.; Lescar, J.; Tam, J.P. Discovery and characterization of pseudocyclic cystine-knot α-amylase inhibitors with high resistance to heat and proteolytic degradation. FEBS J. 2014, 281, 4351–4366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo, G.; Snchez-Monge, R.; Garca-Casado, G.; Armentia, A.; Gomez, L.; Barber, D. The Cerealα-Amylase/Trypsin Inhibitor Family Associated with Bakers’ asthma and Food Allergy. In Plant Food Allergens; Blackwell Publishing Ltd.: Oxford, UK, 2003; pp. 70–86. [Google Scholar]
- Koehler, P.; Wieser, H.; Konitzer, K. Celiac Disease—A Complex Disorder (Chapter 1). In Celiac Disease and Gluten; Koehler, P., Wieser, H., Konitzer, K., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: London, UK, 2014; pp. 1–96. ISBN 9780124202207. [Google Scholar]
- Armentia, A.; Sanchez-Monge, R.; Gomez, L.; Baber, D.; Salcedo, G. In vivo allergenic activities of eleven purified members of a major allergen family from wheat and barley flour. Clin. Exp. Allergy 1993, 23, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Foca, G.; Ulrici, A.; Corbellini, M.; Pagani, M.A.; Lucisano, M.; Franchini, G.C.; Tassi, L. Reproducibility of the Italian ISQ method for quality classification of bread wheats: An evaluation by expert assessors. J. Sci. Food Agric. 2007, 87, 839–846. [Google Scholar] [CrossRef]
- Blandino, M.; Vaccino, P.; Reyneri, A. Late-Season Nitrogen Increases Improver Common and Durum Wheat Quality. Agron. J. 2015, 107, 680–690. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food – an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Abrams, D.; Metcalf, D.; Hojjatie, M. Determination of Kjeldahl Nitrogen in Fertilizers by AOAC Official MethodSM 978.02: Effect of Copper Sulfate as a Catalyst. J. AOAC Int. 2014, 97, 764–767. [Google Scholar] [CrossRef]
- Burton, P.; Lightowler, H.J. The impact of freezing and toasting on the glycaemic response of white bread. Eur. J. Clin. Nutr. 2008, 62, 594–599. [Google Scholar] [CrossRef]
- Bromilow, S.; Gethings, L.A.; Buckley, M.; Bromley, M.; Shewry, P.R.; Langridge, J.I.; Clare Mills, E.N. A curated gluten protein sequence database to support development of proteomics methods for determination of gluten in gluten-free foods. J. Proteom. 2017, 163, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Martens, L.; Vandekerckhove, J.; Gevaert, K. DBToolkit: Processing protein databases for peptide-centric proteomics. Bioinformatics 2005, 21, 3584–3585. [Google Scholar] [CrossRef] [PubMed]
- Juhász, A.; Haraszi, R.; Maulis, C. ProPepper: A curated database for identification and analysis of peptide and immune-responsive epitope composition of cereal grain protein families. Database 2015, 2015, bav100. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.D.; Stephens, R.M. Sequence logos: A new way to display consensus sequences. Nucleic Acids Res. 1990, 18, 6097–6100. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cv | Ashes (%) | GPC (%) | TPC (%) |
|---|---|---|---|---|
| Soft wheat | Altamira | 1.89 ± 0.04 b | 11.17 ± 0.23 a | 8.65 ± 0.01 a |
| Durum wheat | Antalis | 1.82 ± 0.02 a | 11.66 ± 0.35 b | 9.19 ± 0.21 ab |
| Tritordeum | Aucan | 1.96 ± 0.02 c | 13.43 ± 0.15 c | 10.63 ± 0.33 c |
| Tritordeum | Bulel | 1.86 ± 0.04 ab | 11.97 ± 0.07 b | 9.84 ± 0.3 bc |
| Accession | Species | −10LgP | Coverage (%) | Peptides | Description | |
|---|---|---|---|---|---|---|
| PROTEINS IDENTIFIED IN BOTH TRITORDEUM DIGESTS | Q9XGF0 | TRITD | 74.72 | 20 | 12 | LMW-GS |
| A0A446W0B5 | TRITD | 72.20 | 14 | 7 | AAI | |
| K4N1X7 | TRITD | 74.67 | 10 | 8 | HMW-GS | |
| A0A446W0A1 | TRITD | 76.67 | 12 | 9 | AAI | |
| H8Y0D1 | TRITD | 68.82 | 15 | 9 | Alpha prolamin | |
| A0A446W0B4 | TRITD | 63.66 | 12 | 4 | UNP | |
| A0A446W085 | TRITD | 71.15 | 11 | 7 | AAI | |
| A0A446TL77 | TRITD | 39.57 | 5 | 2 | rRNA N-glycosidase | |
| A0A446W0C7 | TRITD | 51.01 | 9 | 3 | AAI | |
| A0A446V2J2 | TRITD | 42.65 | 4 | 2 | AAI | |
| A0A446V2Q9 | TRITD | 45.34 | 8 | 3 | AAI | |
| Q6EEY5 | HORCH | 40.78 | 8 | 3 | Gamma 3 hordein | |
| B0L965 | HORCH | 31.52 | 2 | 1 | D-hordein | |
| A0A446YMF0/M0WF36 | TRITD/HORVV | 21.54 | 4 | 1 | UNP | |
| A0A287EEX5 | TRITD | 40.07 | 6 | 3 | UNP | |
| PROTEIN IDENTIFIED ONLY IN TRITORDEUM CV BULEL | A0A446JGR8 | TRITD | 63.12 | 8 | 5 | AAI |
| A0A0E4G9A4 | TRITD | 48.57 | 6 | 5 | HMW-GS | |
| H8Y0M9 | HORBR | 37.19 | 12 | 3 | Gamma prolamin | |
| A0A7H1K1W3 | TRITD | 31.27 | 7 | 2 | AAI | |
| A0A446IHD3 | TRITD | 20.67 | 6 | 1 | AAI | |
| A0A446IHC0 | TRITD | 31.50 | 4 | 1 | AAI | |
| PROTEINS IDENTIFIED IN TRITORDEUM CV AUCAN | A0A2L1K3K6 | TRITD | 77.43 | 12 | 11 | HMW-GS |
| Q41603 | TRITD | 44.31 | 9 | 3 | LMW-GS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nitride, C.; D’Auria, G.; Dente, A.; Landolfi, V.; Picariello, G.; Mamone, G.; Blandino, M.; Romano, R.; Ferranti, P. Tritordeum as an Innovative Alternative to Wheat: A Comparative Digestion Study on Bread. Molecules 2022, 27, 1308. https://doi.org/10.3390/molecules27041308
Nitride C, D’Auria G, Dente A, Landolfi V, Picariello G, Mamone G, Blandino M, Romano R, Ferranti P. Tritordeum as an Innovative Alternative to Wheat: A Comparative Digestion Study on Bread. Molecules. 2022; 27(4):1308. https://doi.org/10.3390/molecules27041308
Chicago/Turabian StyleNitride, Chiara, Giovanni D’Auria, Andrea Dente, Viola Landolfi, Gianluca Picariello, Gianfranco Mamone, Massimo Blandino, Raffaele Romano, and Pasquale Ferranti. 2022. "Tritordeum as an Innovative Alternative to Wheat: A Comparative Digestion Study on Bread" Molecules 27, no. 4: 1308. https://doi.org/10.3390/molecules27041308
APA StyleNitride, C., D’Auria, G., Dente, A., Landolfi, V., Picariello, G., Mamone, G., Blandino, M., Romano, R., & Ferranti, P. (2022). Tritordeum as an Innovative Alternative to Wheat: A Comparative Digestion Study on Bread. Molecules, 27(4), 1308. https://doi.org/10.3390/molecules27041308