
Understanding the Biosynthesis of Paxisterol in Lichen-Derived Penicillium aurantiacobrunneum for Production of Fluorinated Derivatives



Abstract
:1. Introduction
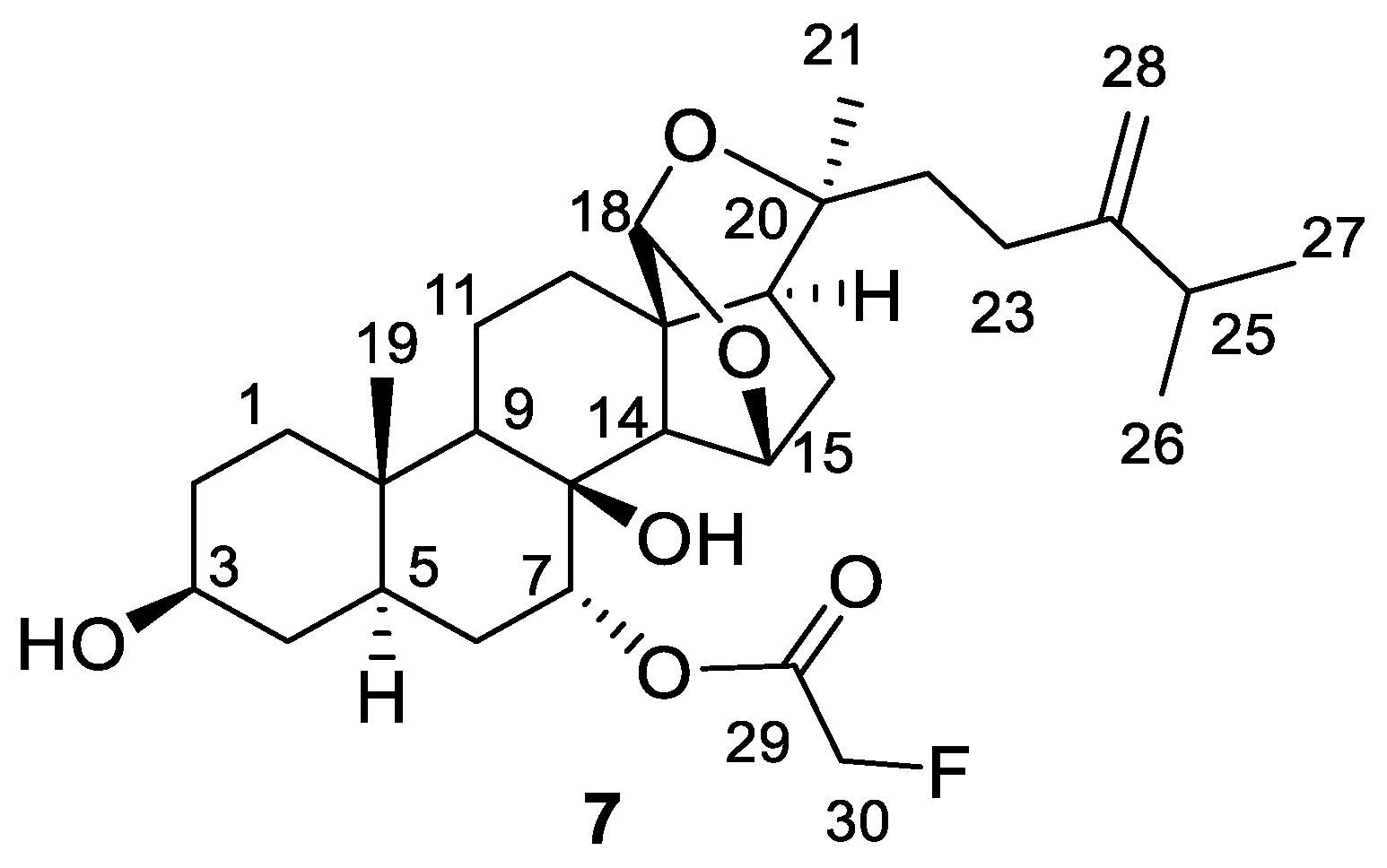
2. Results
3. Materials and Methods
3.1. Fungal Source
3.2. Feeding Assays
3.2.1. 13C-Labelled Glucose
3.2.2. 13C-Labelled Sodium Acetate
3.3. Extraction and Isolation of Compound 7
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Tan, C.Y.; Wang, F.; Anaya-Eugenio, G.D.; Gallucci, J.C.; Goughenour, K.D.; Rappleye, C.A.; Spjut, R.W.; Carcache de Blanco, E.J.; Kinghorn, A.D.; Rakotondraibe, L.H. α-Pyrone and sterol constituents of Penicillium aurantiacobrunneum, a fungal associate of the lichen Niebla homalea. J. Nat. Prod. 2019, 82, 2529–2536. [Google Scholar] [CrossRef] [PubMed]
- Anaya-Eugenio, G.D.; Tan, C.Y.; Rakotondraibe, L.H.; Carcache de Blanco, E.C. Tumor suppressor p53 independent apoptosis in HT-29 cells by auransterol from Penicillium aurantiacobrunneum. Biomed. Pharmacother. 2020, 127, 110124. [Google Scholar] [CrossRef] [PubMed]
- Nakano, H.; Hara, M.; Yamashita, Y.; Ando, K.; Shuto, K. Paxisterol, a new analgesic sterol without anti-inflammation activity from Penicillium. J. Antibiot. 1988, 41, 409–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huestis, M.P.; Terrett, J.A. Simple strategy towards amide bioisosteres. Nat. Chem. 2022, 14, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Singh, R.K. Morpholine as ubiquitous pharmacophore in medicinal chemistry: Deep insight into the structure-activity relationship (SAR). Bioorg. Chem. 2020, 96, 103578. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, K.; Agbaba, D. Pharmacophore development and SAR studies of imidazoline receptor ligands. Mini-Rev. Med. Chem. 2012, 12, 1542–1555. [Google Scholar] [CrossRef] [PubMed]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef] [PubMed]
- Canham, P.L.; Vining, L.C.; McInnes, A.G.; Walter, J.A.; Wright, J.L.C. Use of carbon-13 in biosynthetic studies. Incorporation of 13C-labeled acetate into chartreusin by Streptomyces chartreusis. Can. J. Chem. 1977, 55, 2450–2457. [Google Scholar] [CrossRef]
- Seo, S.; Uomori, A.; Yoshimura, Y.; Takeda, K.; Seto, H.; Ebizuka, Y.; Noguchi, H.; Sankawa, U. Biosynthesis of sitosterol, cycloartenol, and 24-methylenecycloartanol in tissue cultures of higher plants and of ergosterol in yeast from [1,2-13C2]- and [2-13C2H3]-acetate and [5-13C2H2]MVA. J. Chem. Soc. Perkin Trans. 1988, 1, 2407–2414. [Google Scholar] [CrossRef]
- Nabeta, K.; Ichihara, A.; Sakamura, S. Biosynthesis of epoxydone and related compounds by Phyllosticta species. Agric. Biol. Chem. 1975, 39, 409–413. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | (20R)-7α-Fluoroacetoxy-8-hydroxypaxisterol (7 a) | (20R)-7,8-Dihydroxypaxisterol (3 b) | ||
|---|---|---|---|---|
| 1H | 13C | 1H | 13C | |
| δH (m, J in Hz) | δC, Type | δH (m, J in Hz) | δC, Type | |
| 1 | 1.02 (m), 1.77 (m) | 37.6, CH2 | 1.02 (m), 1.77 (m) | 30.2, CH2 |
| 2 | 1.48 (m), 1.75 (m) | 30.2, CH2 | 1.49 (m), 1.75 (m) | 30.2, CH2 |
| 3 | 3.54 (m) | 70.5, CH | 3.58 (ddd, 15.8, 10.9, 4.7) | 70.6, CH2 |
| 4 | 1.34 (m), 1.45 (m) | 36.5, CH2 | 1.37 (m), 1.48 (dd, 12, 5.5) | 36.7, CH2 |
| 5 | 1.57 (m) | 37.5, CH | 1.68 (m) | 36.2, CH |
| 6 | 1.34 (m), 2.09 (m) | 29.3, CH2 | 1.26 (m), 2.06 (m) | 32.1, CH2 |
| 7 | 5.15 (brt, 2.5) | 74.7, CH | 3.74 (t, 2.6) | 71.4, CH |
| 8 | 72.8, C | 73.9, C | ||
| 9 | 1.07 (m) | 49.2, CH | 1.11 (m) | 48.1, CH |
| 10 | 35.5, C | 35.4, C | ||
| 11 | 1.64 (m), 1.80 (m) | 18.5, CH2 | 1.63 (m), 1.80 (m) | 18.4, CH2 |
| 12 | 1.71 (m), 2.24 (m) | 28.2, CH2 | 1.71 (m), 2.27 (m) | 28.2, CH2 |
| 13 | 57.5, C | 57.2, C | ||
| 14 | 1.82 (m) | 55.2, CH | 2.23 (brs) | 55.0, CH |
| 15 | 4.30 (brs) | 74.6, CH | 4.34 (brs) | 74.3, CH |
| 16 | 1.70 (m), 1.92 (d, 13.1) | 34.8, CH2 | 1.83 (m), 1.95 (m) | 34.8, CH2 |
| 17 | 2.14 (d, 9.7) | 49.2, CH | 2.15 (d, 9.5) | 49.1, CH |
| 18 | 5.51 (s) | 107.3, CH | 5.54 (s) | 107.3, CH |
| 19 | 1.00 (a) | 11.7, CH3 | 0.98 (s) | 11.5, CH3 |
| 20 | 85.2, C | 85.0, C | ||
| 21 | 1.33 (s) | 26.4, CH3 | 1.37 (s) | 26.2, CH3 |
| 22 a,b | 1.75 (m) | 39.8, CH2 | 1.79 (m) | 39.6, CH2 |
| 23 | 1.95 (m), 2.02 (m) | 29.5, CH2 | 2.01 (m) | 29.1, CH2 |
| 24 | 155.8, C | 155.7, C | ||
| 25 | 2.26 (sept, 6.8) | 33.9, CH | 2.28 (sept, 6.8) | 33.8, CH |
| 26 | 1.03 (6.8) | 21.1, CH3 | 1.06 (s) | 20.9, CH3 |
| 27 | 1.03 (6.8) | 21.1, CH3 | 1.06 (s) | 20.9, CH3 |
| 28 | 4.71 (brs), 4.76 (brs) | 105.8, CH2 | 4.73 (brs), 4.79 (brs) | 105.6, CH2 |
| CH2F | 4.94 (dd, 46.8, 17.6), 4.97 (dd, 46.8, 17.6) | 77.8 (178.2 Hz), CH2 | ||
| O-C=O | 168.2, C | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamano, Y.; Rakotondraibe, H.L. Understanding the Biosynthesis of Paxisterol in Lichen-Derived Penicillium aurantiacobrunneum for Production of Fluorinated Derivatives. Molecules 2022, 27, 1641. https://doi.org/10.3390/molecules27051641
Yamano Y, Rakotondraibe HL. Understanding the Biosynthesis of Paxisterol in Lichen-Derived Penicillium aurantiacobrunneum for Production of Fluorinated Derivatives. Molecules. 2022; 27(5):1641. https://doi.org/10.3390/molecules27051641
Chicago/Turabian StyleYamano, Yoshi, and Harinantenaina L. Rakotondraibe. 2022. "Understanding the Biosynthesis of Paxisterol in Lichen-Derived Penicillium aurantiacobrunneum for Production of Fluorinated Derivatives" Molecules 27, no. 5: 1641. https://doi.org/10.3390/molecules27051641
APA StyleYamano, Y., & Rakotondraibe, H. L. (2022). Understanding the Biosynthesis of Paxisterol in Lichen-Derived Penicillium aurantiacobrunneum for Production of Fluorinated Derivatives. Molecules, 27(5), 1641. https://doi.org/10.3390/molecules27051641