
Separation, Purification, Structural Characterization, and Anticancer Activity of a Novel Exopolysaccharide from Mucor sp.

 and
and
Abstract
:1. Introduction
2. Results and Discussion
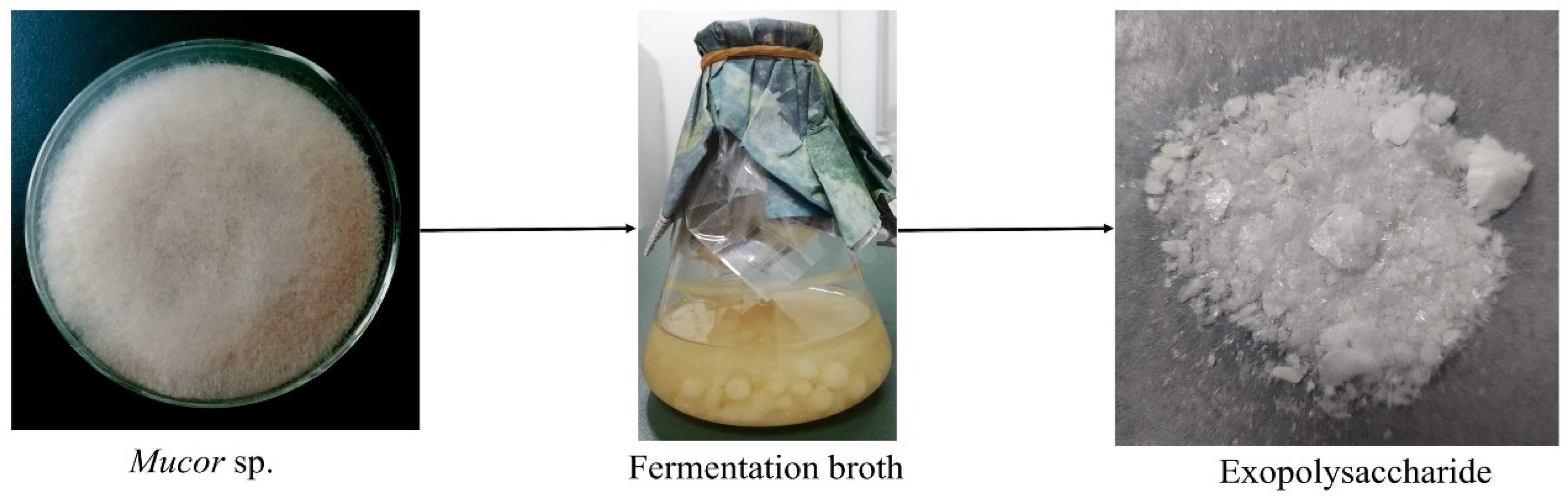
2.1. Isolation and Purification of EPS
2.2. Homogeneity and Molecular Weight
2.3. FT-IR Spectrum
2.4. Monosaccharide Composition
2.5. Methylation Analysis
2.6. NMR Spectroscopy
2.7. Anticancer Activity of MSEPS
2.7.1. Effect of MSEPS on SGC-7901 Cell Inhibition
2.7.2. Effect of MSEPS on Morphological Changes of SGC-7901 Cells
2.7.3. Effect of MSEPS on the Apoptotic Induction of SGC-7901 Cells
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Isolation and Purification of EPS from Mucor sp.
3.3. Characterization
3.3.1. Homogeneity and Molecular Weight of MSEPS
3.3.2. Fourier-Transform Infrared (FT-IR) Spectroscopy Analysis
3.3.3. Monosaccharide Composition Analysis
3.3.4. Methylation Analysis
3.3.5. Nuclear Magnetic Resonance Spectroscopy (NMR) Analysis
3.4. In Vitro Anticancer Activities
3.4.1. Cell Lines and Culture
3.4.2. Measurement of Cell Viability
3.4.3. Morphologic Observations
3.4.4. Hoechst 33,258 Staining
3.4.5. Apoptosis Measurement
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bai, F.R.; Yao, S.; Cai, C.S.; Zhang, T.C.; Wang, Y.; Liu, W.; Ma, B.W.; Rong, C.Y.; Cheng, C. Mucor rongii sp. nov., a New Cold-Tolerant Species from China. Curr. Microbiol. 2021, 78, 2464–2469. [Google Scholar] [CrossRef] [PubMed]
- Taj-Aldeen, S.J.; Almaslamani, M.; Theelen, B.; Boekhout, T. Phylogenetic analysis reveals two genotypes of the emerging fungus Mucor indicus, an opportunistic human pathogen in immunocompromised patients. Emerg. Microbes Infect. 2017, 6, e63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitzman, I.; Dellalatta, P.; Housey, G.; Rebatta, G. Mucor ramosissimus Samutsevitsch isolated from a thigh lesion. J. Clin. Microbiol. 1993, 31, 2523–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Zhang, B.B.; Wu, Z.H. Studies on Mucor racemosus Fermentation to Manufacture Gamma-linolenic Acid Functional Food Douchi. Food Sci. Technol. Res. 2010, 16, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Q.; Hu, Q.; Li, L.T.; Saito, M.; Yin, L.J. Production of Sufu, a Traditional Chinese Fermented Soybean Food, by Fermentation with Mucor flavus at Low Temperature. Food Sci. Technol. Res. 2009, 15, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Han, B.Z.; Kuijpers, A.F.A.; Thanh, N.V.; Nout, M.J.R. Mucoraceous moulds involved in the commercial fermentation of Sufu Pehtze. Antonie Van Leeuwenhoek 2004, 85, 253–257. [Google Scholar] [CrossRef]
- Lima, D.X.; Barreto, R.W.; Lee, H.B.; Cordeiro, T.R.L.; de Souza, C.A.F.; de Oliveira, R.J.V.; Santiago, A.L.C.M.D. Novel Mucoralean Fungus From a Repugnant Substrate: Mucor merdophylus sp. nov., Isolated From Dog Excrement. Curr. Microbiol. 2020, 77, 2642–2649. [Google Scholar] [CrossRef]
- Morin-Sardin, S.; Nodet, P.; Coton, E.; Jany, J.L. Mucor: A Janus-faced fungal genus with human health impact and industrial applications. Fungal Biol. Rev. 2017, 31, 12–32. [Google Scholar] [CrossRef]
- Carvalho, A.K.F.; Faria, E.L.P.; Rivaldi, J.D.; Andrade, G.S.S.; de Oliveira, P.C.; de Castro, H.F. Performance of whole-cells lipase derived from Mucor circinelloides as a catalyst in the ethanolysis of non-edible vegetable oils under batch and continuous run conditions. Ind. Crops Prod. 2015, 67, 287–294. [Google Scholar] [CrossRef]
- de Souza, E.L.; Sales, C.V.; de Oliveira, C.E.V.; Lopes, L.A.A.; da Conceicao, M.L.; Berger, L.R.R.; Stamford, T.C.M. Efficacy of a coating composed of chitosan from Mucor circinelloides and carvacrol to control Aspergillus flavus and the quality of cherry tomato fruits. Front. Microbiol. 2015, 6, 732. [Google Scholar] [CrossRef]
- Huang, J.X.; Zhang, J.; Zhang, X.R.; Zhang, K.; Zhang, X.; He, X.R. Mucor fragilis as a novel source of the key pharmaceutical agents podophyllotoxin and kaempferol. Pharm. Biol. 2014, 52, 1237–1243. [Google Scholar] [CrossRef]
- Lopez-Legarda, X.; Arboleda-Echavarria, C.; Parra-Saldivar, R.; Rostro-Alanis, M.; Alzate, J.F.; Villa-Pulgarin, J.A.; Segura-Sanchez, F. Biotechnological production, characterization and in vitro antitumor activity of polysaccharides from a native strain of Lentinus crinitus. Int. J. Biol. Macromol. 2020, 164, 3133–3144. [Google Scholar] [CrossRef] [PubMed]
- Wan-Mohtar, W.A.Q.I.; Ilham, Z.; Jamaludin, A.A.; Rowan, N. Use of Zebrafish Embryo Assay to Evaluate Toxicity and Safety of Bioreactor-Grown Exopolysaccharides and Endopolysaccharides from European Ganoderma applanatum Mycelium for Future Aquaculture Applications. Int. J. Mol. Sci. 2021, 22, 1675. [Google Scholar] [CrossRef] [PubMed]
- Loncar, J.; Bellich, B.; Parroni, A.; Reverberi, M.; Rizzo, R.; Zjalic, S.; Cescutti, P. Oligosaccharides Derived from Tramesan: Their Structure and Activity on Mycotoxin Inhibition in Aspergillus flavus and Aspergillus carbonarius. Biomolecules 2021, 11, 243. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Lu, Y.; Yu, Z.D.; Xu, L.; Liu, J.; Chen, K.S.; Zhang, P.Y. The inhibitory effect of polysaccharide from Rhizopus nigricans on colitis-associated colorectal cancer. Biomed. Pharmacother. 2019, 112, 108593. [Google Scholar] [CrossRef]
- Xu, L.; Lu, Y.; Cong, Y.Z.; Zhang, P.Y.; Han, J.; Song, G.; Wang, G.D.; Chen, K.S. Polysaccharide produced by Bacillus subtilis using burdock oligofructose as carbon source. Carbohydr. Polym. 2019, 206, 811–819. [Google Scholar] [CrossRef]
- Sun, L.; Yang, Y.B.; Lei, P.; Li, S.; Xu, H.; Wang, R.; Qiu, Y.B.; Zhang, W. Structure characterization, antioxidant and emulsifying capacities of exopolysaccharide derived from Pantoea alhagi NX-11. Carbohydr. Polym. 2021, 261, 117872. [Google Scholar] [CrossRef]
- Li, J.Y.; Yuan, P.F.; Wang, X.H.; Aipire, A.; Li, M.J.; Yang, J.; Tao, H.; Ying, T.L.; Fu, C.S.; Wei, X.X.; et al. Purification, characterization and bioactivities of polysaccharides from Pleurotus ferulae. Food Funct. 2017, 8, 1905–1914. [Google Scholar] [CrossRef]
- Li, L.Q.; Song, A.X.; Wong, W.T.; Wu, J.Y. Isolation and Assessment of a Highly-Active Anti-Inflammatory Exopolysaccharide from Mycelial Fermentation of a Medicinal Fungus Cs-HK1. Int. J. Mol. Sci. 2021, 22, 2450. [Google Scholar]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef]
- Wu, M.B.; Zhang, F.F.; Yu, Z.P.; Lin, J.P.; Yang, L.R. Chemical characterization and in vitro antitumor activity of a single-component polysaccharide from Taxus chinensis var. mairei. Carbohydr. Polym. 2015, 133, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.S.; Zeng, Y.X.; Bi, S.Y.; Zhou, J.W.; Cheng, R.; Li, J.; Jia, A.Q. Characterization and chemical modification of PLN-1, an exopolysaccharide from Phomopsis liquidambari NJUSTb1. Carbohydr. Polym. 2021, 253, 117197. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.G.; Yan, W.; Lv, J.L.; Yao, W.B.; Yu, L.L. Characterization of a Novel Polysaccharide from Tetraploid Gynostemma pentaphyllum Makino. J. Agric. Food Chem. 2013, 61, 4882–4889. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, T.; Wang, H.J.; Cui, Z.; Cheng, F.; Wang, K.P. Structural characterization and in vitro antitumor activity of an acidic polysaccharide from Angelica sinensis (Oliv.) Diels. Carbohydr. Polym. 2016, 147, 401–408. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.M.; Qin, G.Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef]
- Zhang, H.R.; Wang, X.Q.; Li, R.F.; Sun, X.C.; Sun, S.W.; Li, Q.; Xu, C.P. Preparation and Bioactivity of Exopolysaccharide from an Endophytic Fungus Chaetomium sp. of the Medicinal Plant Gynostemma Pentaphylla. Pharmacogn. Mag. 2017, 13, 477–482. [Google Scholar]
- Chikari, F.; Han, J.; Wang, Y.; Ao, W.M. Synergized subcritical-ultrasound-assisted aqueous two-phase extraction, purification, and characterization of Lentinus edodes polysaccharides. Process Biochem. 2020, 95, 297–306. [Google Scholar] [CrossRef]
- Chou, C.H.; Sung, T.J.; Hu, Y.N.; Lu, H.Y.; Yang, L.C.; Cheng, K.C.; Lai, P.S.; Hsieh, C.W. Chemical analysis, moisture-preserving, and antioxidant activities of polysaccharides from Pholiota nameko by fractional precipitation. Int. J. Biol. Macromol. 2019, 131, 1021–1031. [Google Scholar] [CrossRef]
- Sun, Y.Q.; Huo, J.X.; Zhong, S.; Zhu, J.X.; Li, Y.G.; Li, X.J. Chemical structure and anti-inflammatory activity of a branched polysaccharide isolated from Phellinus baumii. Carbohydr. Polym. 2021, 268, 118214. [Google Scholar] [CrossRef]
- Zhai, Z.Y.; Chen, A.; Zhou, H.M.; Zhang, D.Y.; Du, X.H.; Liu, Q.; Wu, X.Y.; Cheng, J.E.; Chen, L.J.; Hu, F.; et al. Structural characterization and functional activity of an exopolysaccharide secreted by Rhodopseudomonas palustris GJ-22. Int. J. Biol. Macromol. 2021, 167, 160–168. [Google Scholar] [CrossRef]
- Yin, J.Y.; Wang, J.Q.; Lin, H.X.; Xie, M.Y.; Nie, S.P. Fractionation, physicochemical properties and structural features of non-arabinoxylan polysaccharide from the seeds of Plantago asiatica L. Food Hydrocoll. 2016, 55, 128–135. [Google Scholar] [CrossRef]
- Tian, J.J.; Wang, X.M.; Zhang, X.L.; Zhang, C.P.; Chen, X.H.; Dong, M.S.; Rui, X.; Zhang, Q.Q.; Fang, Y.; Li, W. Isolation, structural characterization and neuroprotective activity of exopolysaccharide from Paecilomyces cicada TJJ1213. Int. J. Biol. Macromol. 2021, 183, 1034–1046. [Google Scholar] [CrossRef] [PubMed]
- Rozi, P.; Abuduwaili, A.; Mutailifu, P.; Gao, Y.H.; Rakhmanberdieva, R.; Aisa, H.A.; Yili, A. Sequential extraction, characterization and antioxidant activity of polysaccharides from Fritillaria pallidiflora Schrenk. Int. J. Biol. Macromol. 2019, 131, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.W.; Song, J.L.; Liu, Y.; Lu, N.; Wang, Y.B.; Hu, C.J.; He, L.; Wei, H.L.; Lv, G.Y.; Yang, S.Z.; et al. Conformational properties and biological activities of α-D-mannan from Sanghuangporus sanghuang in liquid culture. Int. J. Biol. Macromol. 2020, 164, 3568–3579. [Google Scholar] [CrossRef]
- Pei, J.J.; Wang, Z.B.; Ma, H.L.; Yan, J.K. Structural features and antitumor activity of a novel polysaccharide from alkaline extract of Phellinus linteus mycelia. Carbohydr. Polym. 2015, 115, 472–477. [Google Scholar] [CrossRef]
- Wang, C.Y.; Mao, W.J.; Chen, Z.Q.; Zhu, W.M.; Chen, Y.L.; Zhao, C.Q.; Li, N.; Yan, M.X.; Liu, X.; Guo, T.T. Purification, structural characterization and antioxidant property of an extracellular polysaccharide from Aspergillus terreus. Process Biochem. 2013, 48, 1395–1401. [Google Scholar] [CrossRef]
- Chen, W.B.; Zhu, X.L.; Ma, J.J.; Zhang, M.M.; Wu, H. Structural Elucidation of a Novel Pectin-Polysaccharide from the Petal of Saussurea laniceps and the Mechanism of its Anti-HBV Activity. Carbohydr. Polym. 2019, 223, 115077. [Google Scholar] [CrossRef]
- Xie, X.; Shen, W.; Zhou, Y.R.; Ma, L.M.; Xu, D.Y.; Ding, J.L.; He, L.Y.; Shen, B.Y.; Zhou, C.L. Characterization of a polysaccharide from Eupolyphaga sinensis walker and its effective antitumor activity via lymphocyte activation. Int. J. Biol. Macromol. 2020, 162, 31–42. [Google Scholar] [CrossRef]
- Wang, H.T.; Yang, L.C.; Yu, H.C.; Chen, M.L.; Wang, H.J.; Lu, T.J. Characteristics of fucose-containing polysaccharides from submerged fermentation of Agaricus blazei Murill. J. Food Drug Anal. 2018, 26, 678–687. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.F.; Liu, M.; Fang, Z.X.; Zhang, Q.Q. In Vitro antioxidant effects and cytotoxicity of polysaccharides extracted from Laminaria japonica. Int. J. Biol. Macromol. 2012, 50, 1254–1259. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhang, T.; Jiang, B.; Mu, W.M.; Miao, M. Characterization and antioxidant activity of Ginkgo biloba exocarp polysaccharides. Carbohydr. Polym. 2012, 87, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.L.; Lin, X.; Shen, L.; Wu, F.; Xu, D.S.; Ruan, K.F.; Feng, Y. Mono-PEGylated radix ophiopogonis polysaccharide for the treatment of myocardial ischemia. Eur. J. Pharm. Sci. 2013, 49, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Q.; Zhang, L.M. Chemical structural and chain conformational characterization of some bioactive polysaccharides isolated from natural sources. Carbohydr. Polym. 2009, 76, 349–361. [Google Scholar] [CrossRef]
- Jiang, P.; Zhang, Q.; Zhao, Y.J.; Xiong, J.; Wang, F.; Zhang, T.; Zhang, C.M. Extraction, Purfication, and Biological Activities of Polysaccharides from Branches and Leaves of Taxus cuspidata S. et Z. Molecules 2019, 24, 2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Jeddou, K.; Chaari, F.; Maktouf, S.; Nouri-Ellouz, O.; Helbert, C.B.; Ghorbel, R.E. Strutural, functional, and antioxidant properties of water-soluble polysaccharides from potatoes peels. Food Chem. 2016, 205, 97–105. [Google Scholar] [CrossRef]
- Ji, X.L.; Cheng, Y.Q.; Tian, J.Y.; Zhang, S.Q.; Jing, Y.S.; Shi, M.M. Structural characterization of polysaccharide from jujube (Ziziphus jujuba Mill.) fruit. Chem. Biol. Technol. Agric. 2021, 8, 54. [Google Scholar] [CrossRef]
- Liu, W.B.; Xie, F.; Sun, H.Q.; Meng, M.; Zhu, Z.Y. Anti-tumor effect of polysaccharide from Hirsutella sinensis on human non-small cell lung cancer and nude mice through intrinsic mitochondrial pathway. Int. J. Biol. Macromol. 2017, 99, 258–264. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | Monosaccharide Composition | Relative Molar Ratio |
|---|---|---|
| 5.684 | fucose | 0.139 |
| 11.359 | rhamnose | 0 |
| 11.909 | arabinose | 0.126 |
| 14.900 | galactose | 0.169 |
| 16.967 | glucose | 0.015 |
| 20.209 | xylose | 0 |
| 20.750 | mannose | 0.466 |
| 24.367 | fructose | 0 |
| 27.884 | ribose | 0 |
| 44.942 | galacturonic acid | 0.005 |
| 45.992 | guluronic acid | 0 |
| 48.034 | glucuronic acid | 0.008 |
| 50.817 | mannuronic acid | 0 |
| RT | Methylated Sugar | Mass Fragments (m/z) | Molar Ratios | Type of Linkage |
|---|---|---|---|---|
| 9.223 | 2,3,5-Me3-Araf | 43,71,87,101,117,129,145,161 | 0.119 | Araf-(1→ |
| 11.669 | 2,3,4-Me3-Fucp | 43,59,72,89,101,115,117,131,175 | 0.128 | Fucp-(1→ |
| 16.274 | 2,3,4,6-Me4-Manp | 43,71,87,101,117,129,145,161,205 | 0.025 | Manp-(1→ |
| 17.271 | 2,3,4,6-Me4-Galp | 43,71,87,101,117,129,145,161,205 | 0.018 | Galp-(1→ |
| 20.564 | 3,4,6-Me3-Manp | 43,87,129,161,189 | 0.041 | →2)-Manp-(1→ |
| 20.885 | 2,3,6-Me3-Galp | 43,87,99,101,113,117,129,131,161,173,233 | 0.025 | →4)-Galp-(1→ |
| 22.241 | 2,3,4-Me3-Glcp | 43,87,99,101,117,129,161,189,233 | 0.009 | →6-Glcp-(1→ |
| 22.474 | 2,3,4-Me3-Manp | 43,71,87,99,101,117,129,159,161 | 0.282 | →6)-Manp-(1→ |
| 24.166 | 2,3,4-Me3-Galp | 43,87,99,101,117,129,161,189,233 | 0.052 | →6)-Galp-(1→ |
| 28.430 | 2,4-Me2-Manp | 43,87,117,129,159,189,233 | 0.099 | →3,6)-Manp-(1→ |
| 29.438 | 2,4-Me2-Galp | 43,87,117,129,159,189,233 | 0.066 | →3,6)-Galp-(1→ |
| Glycosyl Residues | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6a/C6 | H6b | ||
|---|---|---|---|---|---|---|---|---|---|
| A |  | →2)-α-d-Manp-(1→ | 5.21 | 4.00 | 3.88 | 3.60 | 3.70 | 3.69 | 3.81 |
| 101.75 | 80.21 | 71.32 | 68.19 | 74.50 | 62.30 | ||||
| B |  | α-l-Fucp-(1→ | 5.16 | 3.52 | nd | nd | nd | 1.14 | |
| 101.96 | 73.23 | nd | nd | nd | 15.31 | ||||
| C |  | α-l-Araf-(1→ | 5.01 | 4.13 | 3.83 | 3.91 | 3.69 | ||
| 108.77 | 82.70 | 77.80 | 85.10 | 62.33 | |||||
| D |  | →3)-α-d-Manp-(1→ | 4.99 | 3.74 | 3.78 | 4.24 | 3.91 | 3.73 | |
| 95.78 | 69.36 | 80.50 | 68.33 | 71.00 | 62.10 | ||||
| E |  | →3,6)-α-d-Manp-(1→ | 4.87 | 4.09 | 3.82 | 3.65 | 3.73 | 3.68 | 3.87 |
| 100.84 | 68.18 | 79.46 | 72.70 | 74.64 | 66.91 | ||||
| F |  | →6)-α-d-Manp-(1→ | 4.82 | 3.91 | 3.75 | 3.75 | 3.68 | 3.92 | 3.89 |
| 100.80 | 71.37 | 71.87 | 67.89 | 74.50 | 66.82 | ||||
| G |  | →4)-β-d-Galp-(1→ | 4.57 | 3.61 | 3.69 | 4.10 | 3.63 | 3.73 | |
| 105.80 | 73.21 | 74.84 | 79.66 | 76.01 | 62.1 | ||||
| H |  | →3,6)-β-d-Galp-(1→ | 4.46 | 3.57 | 3.68 | 4.05 | 3.87 | 3.96 | 3.86 |
| 104.69 | 71.31 | 81.50 | 69.82 | 74.81 | 70.76 | ||||
| I |  | β-d-Galp-(1→ | 4.25 | 3.28 | 3.46 | 3.21 | 3.60 | 3.54 | 3.41 |
| 104.48 | 74.66 | 76.62 | 83.40 | 77.36 | 64.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yuan, P.; Zhang, W.; Liu, C.; Chen, K.; Wang, G.; Shao, T. Separation, Purification, Structural Characterization, and Anticancer Activity of a Novel Exopolysaccharide from Mucor sp. Molecules 2022, 27, 2071. https://doi.org/10.3390/molecules27072071
Wang J, Yuan P, Zhang W, Liu C, Chen K, Wang G, Shao T. Separation, Purification, Structural Characterization, and Anticancer Activity of a Novel Exopolysaccharide from Mucor sp. Molecules. 2022; 27(7):2071. https://doi.org/10.3390/molecules27072071
Chicago/Turabian StyleWang, Jiaojiao, Pingchuan Yuan, Wenzhi Zhang, Chunyan Liu, Kaoshan Chen, Guodong Wang, and Taili Shao. 2022. "Separation, Purification, Structural Characterization, and Anticancer Activity of a Novel Exopolysaccharide from Mucor sp." Molecules 27, no. 7: 2071. https://doi.org/10.3390/molecules27072071