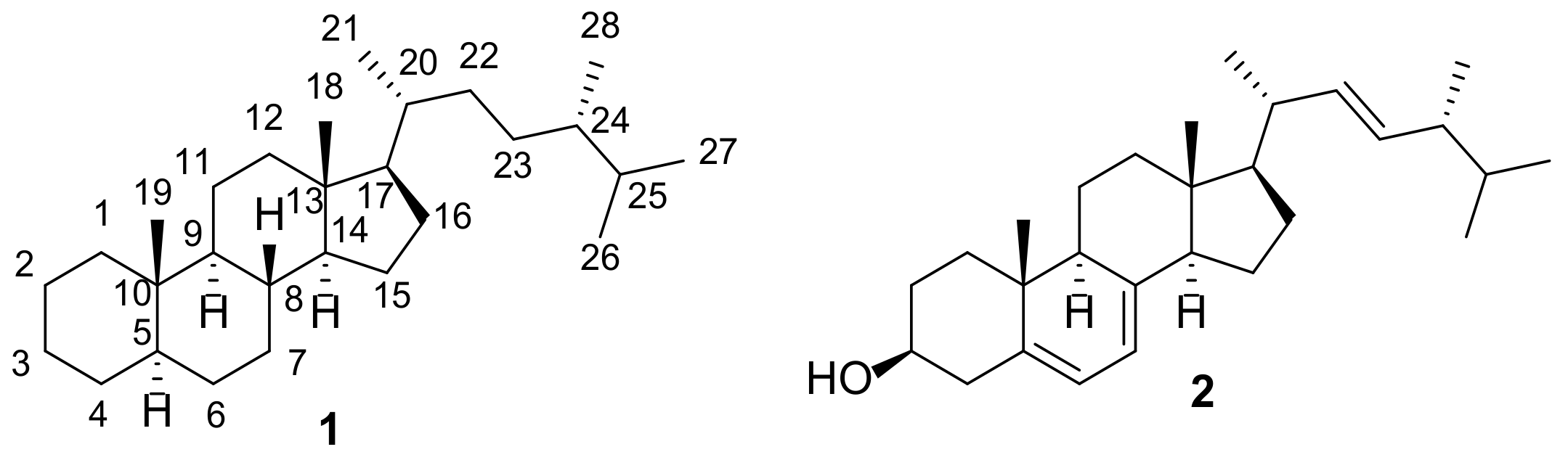
3. Endoperoxides
Compounds containing a peroxide group are quite widespread among various natural substances, and steroids are not an exception [
27]. Two 5α,8α-endoperoxides, ergosterol peroxide (EP,
21a) and 9,11-dehydroergosterol peroxide (DHEP,
22a) (
Figure 5), are the most typical representatives of fungal steroids. Publications up to 2016 on the biological activity of EP (
5a) have been thoroughly reviewed by Merdivan and Lindequist [
24], and only the more recent literature regarding this compound will be discussed here.
Biological studies of endoperoxides
21a and
22a have been aimed primarily at assessing their cytotoxic potential. Both compounds revealed quite high level of cytotoxicity in a wide range of cancer cells (
Table 1). It should be noted that measurements of cell toxicity often vary significantly from laboratory to laboratory. Thus, for EP and cell line MCF-7 the values of IC
50 varied from IC
50 1.18 μM [
122] to 151 μM [
123].
Attempts have been made to understand the cytotoxicity mechanism for
21a, and some authors have concluded that more than one mechanism is at work. Obviously, the peroxide bridge plays a crucial role, bearing in mind that ergosterol is not cytotoxic. It was assumed that induction of apoptosis is the main cause of cytotoxicity [
24]. Homolytic cleavage of the peroxide moiety in a reducing medium leads to the formation of reactive oxygen species (ROS), which are powerful internal stimuli for apoptosis. This has been confirmed, in particular, in experiments with MCF-7 cells [
124]. Their treatment with
21a at concentrations of 40–80 μg/mL led to an increase in the production of ROS in a dose-dependent manner and to the induction of apoptosis. The inhibitory properties of
21a against A549 lung cancer cells were mediated by mitochondria-dependent apoptosis and autophagy [
125]. EP also reduced LPS/ATP-induced proliferation and migration of A549 cells. A synergistic effect was observed when using EP with kinase inhibitor Sorafenib.
Based on ID
50 values for the MCF-7 cell line (1.18 μM) compared to the MDA-MB-231 cell line (12.82 μM), EP (
21a) was hypothesized to target estrogen receptors [
122]. Its possible role as an ERα antagonist was suggested by Kim et al. based on the suppression of the increase in the viability of MCF-7 cells caused by 17β-estradiol [
126].
Ergosterol peroxide (
21a) and 9,11-dehydroergosterol peroxide (
22a) were often isolated from the same fungal material, and on the whole both compounds exhibit similar biological properties. DHEP (
22a) was slightly more cytotoxic than EP (
21a) on the Hep 3B cell viability (IC
50 16.7 and 19.4 μg/mL, respectively) [
127]. In experiments with BV-2 microglia cells, compound
22a did not damage cell viability, although EP was cytotoxic to these cells [
128]. Kobori et al. showed that
22a selectively inhibits the growth of HT29 human colon adenocarcinoma cells without affecting normal human WI38 fibroblasts [
129]. The inhibition was attributed to the induction of expression of an inhibitor of cyclin-dependent kinase 1A, thus causing cell cycle arrest and apoptosis. The rather strong cytotoxic effect of
22a (IC
50 8.58 μM) on HeLa human cervical carcinoma cells was associated with the regulated expression of stathmin 1, a protein that is critical for the regulation of the cell cytoskeleton [
130]. The mechanisms of
22a cytotoxicity in A375 melanoma cells have been shown to be caspase-dependent and mediated via the mitochondrial pathway and include targeting of the induced differentiation protein of myeloid leukemia cells Mcl-1, release of cytochrome c, and activation of caspase-9 and -3 [
131].
In experiments with a large number of cell lines EP possessed cytotoxic activity at the level of 1 μM and was more active in comparison with DHEP [
132]. On the other hand, in the aromatase inhibitory assay 9(11)-double-bond enhances the inhibitory activity (IC
50 > 100 μM vs. 32.6 μM for EP and DHEP, respectively) [
59].
EP was thought to be one of the main compounds responsible for the antiproliferative effect of an ethanolic extract of the native New Zealand mushroom
Hericium novae-zealandiae [
133]. Two possible mechanisms of the observed effect have been proposed: apoptosis based on upregulation of CASP3, CASP8, CASP9, and anti-inflammation, as follows from downregulation of IL6 and upregulation of IL24.
Studying the cytotoxic effects on renal cell carcinoma cells, Zhang et al. found that EP treatment suppressed cell growth, colonization, migration and invasion, arrested the cell cycle, and triggered apoptosis [
134]. This also means that several mechanisms can act for the same effect.
A similar situation with multiple pathways was observed in experiments with ovarian cancer cells [
135]. Their treatment with
21a inhibited nuclear β-catenin, thus decreasing the expression levels of cyclin D1 and c-Myc. Meanwhile, the level of protein tyrosine phosphatase SHP2 was increased in the treated cells, while the activity of Src kinase was suppressed. Thus, the antitumor effect of
21a on ovarian cancer cells is due to both the β-catenin and STAT3 signaling pathways.
Significant inhibition of the formation of experimental lung metastases in vivo was found for EP (
21a) [
136]. The effect was attributed to inhibition of the NF-κB and STAT3 inflammatory pathways in 4T1 breast cancer cells.
EP was more effective than cisplatin in a mouse tumor model, inhibiting CT26 cell growth and improving the survival of tumor mice with no obvious side effects [
137]. The growth of tumor cells of the gastrointestinal tract was suppressed due to the induction of apoptosis by the stress of the endoplasmic reticulum and mitochondria-dependent pathway.
Table 1.
Cytotoxicity of fungal endoperoxides on different cell lines.
Table 1.
Cytotoxicity of fungal endoperoxides on different cell lines.
| Compound | Cell Line | Origin * | Effect [Ref.] |
|---|
| 21a | 4T1 | Mouse breast cancer | IC50 9.06 μM [138] |
| A549 | Lung carcinoma | IC50 17.04 μM [138], IC50 17.2 μM [84], IC50 > 20 μM [139], IC50 23 μM [125], IC50 57 μM [140] |
| B 16 | Murine melanoma | IC50 78.77 μM [141] |
| B16F10 | Murine melanoma | IC50 55.8 μM [142] |
| BGC823 | Gastric cancer | IC50 35.23 μg/mL [137] |
| Eca-109 | Esophageal carcinoma | IC50 23.17 μg/mL [137] |
| DU145 | Prostate cancer | IC50 21 μg/mL [133] |
| HCT116 | Colorectal carcinoma | IC50 80.72 μM [142] |
| HeLa | Cervical carcinoma | IC50 13.6 μM [84], IC50 > 20 μM [139], IC50 31 μM [125], IC50 > 50 μM [143], IC50 > 50 μM [138] |
| Hep 3B | Hepatocellular carcinoma | IC50 35.2 μg/mL [144] |
| HepG2 | Liver carcinoma | IC50 13.19 μM [138], IC50 > 20 μM [139], IC50 23.15 μM [145], IC50 23.5 μM [146], IC50 34 μM [147], IC50 46.9 μM [144], IC50 113 μM [123] |
| HL-60 | Promyelocytic leukemia | IC50 39.4 μM [143] |
| HT-29 | Colon adenocarcinoma | IC50 25.47 μM [137], IC50 > 50 μM [138] |
| J5 | Hepatocellular carcinoma | IC50 33 μM [125] |
| L1210 | Mouse lymphotic leukemia | IC50 36.40 μM [138] |
| LNCap | Prostate cancer | IC50 15 μg/mL [133], IC50 35.53 μg/mL [141] |
| LS180 | Colon adenocarcinoma | IC50 17.3 μg/mL [148] |
| MDA-MB-231 | Breast carcinoma | IC50 12.82 μM [122], EC50 18 μM [149], IC50 24.75 μM [146], IC50 44.6 μM [147] |
| MCF-7 | Breast cancer | IC50 1.18 μM [122], IC50 9.01 μM [138], IC50 26 μM [140], IC50 26.06 μM [145,146], IC50 29 μM [125], IC50 38.2 μM [143], IC50 40 μM [124], IC50 98.12 μM [142], IC50 > 100 μM [126,144], IC50 151 μM [123] |
| MGC-803 | Gastric carcinoma | IC50 15.2 μM [84] |
| NCI 60 panel | | significant activity against most tumor cell lines tested [132] |
| PC3 | Prostate cancer | IC50 42 μg/mL [133] |
| PC-3M | Prostatic carcinoma | IC50 23.15 μM [123] |
| RCC | Renal carcinoma | IC50 30 μM [134] |
| SK-Hep1 | Liver cancer | IC50 19.25 μM [145], IC50 19.71 μM [146] |
| SUM-149 | Breast cancer | EC50 9 μM [149], EC50 20 μM [150] |
| T-47D | Breast cancer | EC50 19 μM [149] |
| 21b | A549 | Lung carcinoma | IC50 14.21 μM [151] |
| HCT-15 | Colon adenocarcinoma | IC50 17.49 μM [151] |
| SK-MEL-2 | Skin melanoma | IC50 9.01 μM [151] |
| SK-OV-3 | Ovary malignant ascites | IC50 15.11 μM [151] |
| U87 | Glioblastoma | 20.1% inhibition at 100 μM [152] |
| 21c | HepG2 | Liver carcinoma | IC50 12.34 (n = 1), 9.46 (n = 2), 6.74 (n = 3) μM [145] |
| MCF-7 | Breast cancer | IC50 14.80 (n = 1), 13.70 (n = 2), 7.45 (n = 3) μM [145] |
| SK-Hep1 | Liver cancer | IC50 10.43 (n = 1), 11.70 (n = 2), 5.92 (n = 3) μM [145] |
| 21d | HepG2 | Liver carcinoma | 6.60 μM [145] |
| MCF-7 | Breast cancer | 10.62 μM [145] |
| SK-Hep1 | Liver cancer | 8.10 μM [145] |
| 21e | MDA-MB-231 | Breast carcinoma | EC50 7 μM [149] |
| SUM-149 | Breast cancer | EC50 2 μM [149] |
| T-47D | Breast cancer | EC50 16 μM [149] |
| 21f | HCT-116 | Colon carcinoma | IC50 0.21 μM [153] |
| 21g | SUM-149 | Breast cancer | EC50 12 μM [150] |
| 21h | MDA-MB-231 | Breast carcinoma | EC50 10 μM [149] |
| SUM-149 | Breast cancer | EC50 4 μM [149] |
| T-47D | Breast cancer | EC50 > 10 μM [149] |
| 21i | HepG2 | Liver carcinoma | IC50 0.85 μM [146] |
| MCF-7 | Breast cancer | IC50 3.26 μM [146] |
| MDA-MB-231 | Breast carcinoma | IC50 4.12 μM [146] |
| SK-Hep1 | Liver cancer | IC50 1.75 μM [146] |
| 21j | HepG2 | Liver carcinoma | IC50 2.83 μM [146] |
| MCF-7 | Breast cancer | IC50 4.62 μM [146] |
| MDA-MB-231 | Breast carcinoma | IC50 3.99 μM [146] |
| SK-Hep1 | Liver cancer | IC50 0.92 μM [146] |
| 22a | 4T1 | Mouse breast cancer | IC50 9.31 μM [138] |
| A375 | Malignant melanoma | IC50 9.46 μg/mL [131] |
| A549 | Lung carcinoma | IC50 9.7 μM [84], IC50 10.77 μM [138], IC50 49 μM [125], IC50 63 μM [140], IC50 103.74 μM [154], IC50 121.9 μM [155], No cytotoxicity [156] |
| Calu-6 | Lung carcinoma | IC50 71.2 μM [155] |
| Colo201 | Colorectal adenocarcinoma | IC50 13.02 μg/mL [131] |
| H1264 | Lung carcinoma | IC50 92.3 μM [155] |
| H1299 | Lung carcinoma | IC50 50.6 μM [155] |
| HeLa | Cervical carcinoma | IC50 7.6 μM [84], IC50 8.58 μM [130], IC50 35.82 μM [138], IC50 37 μM [125] |
| Hep 3B | Hepatocellular carcinoma | IC50 16.7 μg/mL [127] |
| HepG2 | Liver carcinoma | IC50 10.93 μM [138], IC50 44.5 μM [147], IC50 64.95 μM[154] |
| HGC27 | Gastric carcinoma | IC50 26.47 μM [16] |
| HT-29 | Colon adenocarcinoma | IC50 30.76 μM [138] |
| J5 | Hepatocellular carcinoma | IC50 36 μM [125] |
| L1210 | Mouse lymphotic leukemia | IC50 29.31 μM [138] |
| MCF-7 | Breast cancer | IC50 3.3 μM [140], IC50 8.40 μM [138], IC50 16.89 μg/mL [131], IC50 34 μM [125], IC50 67.89 μg/mL [131], IC50 > 100 μM [126] |
| MDA-MB-231 | Breast carcinoma | IC50 72.68 μM [154], IC50 99 μM [16], IC50 328 μM [147] |
| MGC-803 | Gastric carcinoma | IC50 7.8 μM [84] |
| Panc-28 | Pancreatic adenocarcinoma | No cytotoxicity [156] |
| SW620 | Colorectal adenocarcinoma | IC50 32.87 μg/mL [131] |
| 22b | A549 | Lung carcinoma | No cytotoxicity [156] |
| A549 | Lung carcinoma | IC50 15.42 μM [151] |
| HCT-15 | Colon adenocarcinoma | IC50 19.32 μM [151] |
| Panc-28 | Pancreatic adenocarcinoma | No cytotoxicity [156] |
| SK-MEL-2 | Skin melanoma | IC50 12.96 μM [151] |
| SK-OV-3 | Ovary malignant ascites | IC50 18.26 μM [151] |
| 27 | A549 | Lung carcinoma | IC50 5.26 μg/mL [12] |
| MCF-7 | Breast cancer | IC50 5.15 μg/mL [12] |
| 28 | A549 | Lung carcinoma | IC50 0.26 μg/mL [157] |
| HSC-3 | Oral squamous cell carcinoma | IC50 1.72 μg/mL [157] |
| HSC-4 | Oral squamous cell carcinoma | IC50 1.94 μg/mL [157] |
| MKN45 | Stomach adenocarcinoma | IC50 0.34 μg/mL [157] |
Compound
21a can be used as a radiosensitizer in the treatment of cervical cancer to reduce the toxic effects that occur after ionizing radiation therapy. Loss of viability of the cervical cell lines HeLa and CaSki was observed with increasing dose of
21a [
158].
Biological effects of EP (
21a) and its Δ
9,11-counterpart
22a are not limited to their cytotoxic and anticancer properties. A detailed study on the bioactivity of the components of the truffle
Reddellomyces parvulosporus revealed a number of EP activities, including anti-tyrosinase, anti-urease, anti-α-glucosidase, and anti-α-amylase ones [
159]. Tyrosinase is an enzyme involved in the biosynthesis of melanin in humans, and its inhibitors are of interest for preventing excessive melanin production, as being active ingredients of skin whitening agents. Tyrosinase inhibitory activity (IC
50: 202.37 μg/mL) of EP was also detected by Bai et al. [
160].
Ng et al. reported the antidiabetic effect of
21a that was due to the upregulation of glucose absorption and modulation of the PI3K/Akt, MAPK, and GLUT-4 signaling pathways [
161].
EP was tested for its antileishmania activity against
Leishmania donovani promastigotes and showed good activity with IC
50 values of 9.43 μM [
162]. The EP trypanocidal activity has been associated with its interaction with CYP51 [
163]. The key structural moiety responsible for this is the peroxide bridge, which mediates interaction with the CYP51 heme binding site. At a later stage, this can cause the appearance of free radicals through homolytic cleavage at the O-O site, the pharmacophore responsible for the biological activity of
21a.
Zhou et al. studied the immunoregulatory effect on inflammation caused by influenza A virus in human alveolar epithelial cells A549. EP (
21a) was found to have anti-inflammatory effects and prevent virus-induced apoptosis by attenuating retinoic acid-inducible gene I signaling in infected cells [
164].
Oral administration of EP to piglets infected with porcine delta-coronavirus resulted in a reduction in diarrhea, relief of intestinal damage, and a decrease in viral load in feces and tissues [
165]. Wang et al. demonstrated that ergosterol peroxide prevents infection by suppressing porcine delta-coronavirus-induced autophagy via the p38 signaling pathway [
166,
167].
DHEP (
22a) was found to exhibit strong anti-inflammatory effect in lipopolysaccharide-stimulated RAW264.7 cells [
168,
169,
170]. It suppressed the production of NO even at 12.5 μM and pro-inflammatory cytokines interleukin 6 at 25 μM [
168].
With age, mesenchymal stem cells in bone marrow tend to differentiate more into adipocytes than into osteocytes. Compounds
21a and
22a have been shown to inhibit the differentiation of mesenchymal stem cells toward adipocytes, which may be useful for the treatment of postmenopausal osteoporosis [
171].
In experiments with 3T3-L1 mouse embryonic fibroblast cells, it was shown that EP inhibits triglyceride synthesis and reduces the accumulation of lipid droplets by suppressing adipogenesis [
172].
The endoperoxides
21a and
22a were tested for their antibacterial activity [
173,
174,
175,
176,
177]. The presence of a 9,11-double bond contributed to the increase in activity [
173,
177]. Thus, Δ
9,11-derivative
22a was more effective against
M. tuberculosis H37Rv in comparison with
21a (MIC 16 μg/mL and 64 μg/mL, respectively) [
173]. Antitubercular activity of the fungus
Gliocladium sp. MR41., was tested on M. tuberculosis. It was found to be due to EP (
21a) with MIC 0.78 μg/mL [
178].
Kim et al. isolated glucosides
21b and
22b from the Korean wild fungus
Xerula furfuracea and tested their effects on adipogenesis and osteogenesis in a mouse mesenchymal stem cell line [
10]. Both compounds were found to inhibit the differentiation of stem cells into adipocytes, which is of interest in the treatment of syndromes associated with menopause.
Significant antifungal and cytotoxic activities were reported for EP decanoate (
21f) [
153]. In disk diffusion test against
Candida albicans culture, its MIC value was found to be 8.3 μg/disc that was comparable to clotrimazole (MIC 5.1 μg/disc). Compound
21f showed also very good cytotoxicity against the HCT-116 cell line with IC
50 value of 0.21 μM compared to doxorubicin (IC
50 0.06 μM).
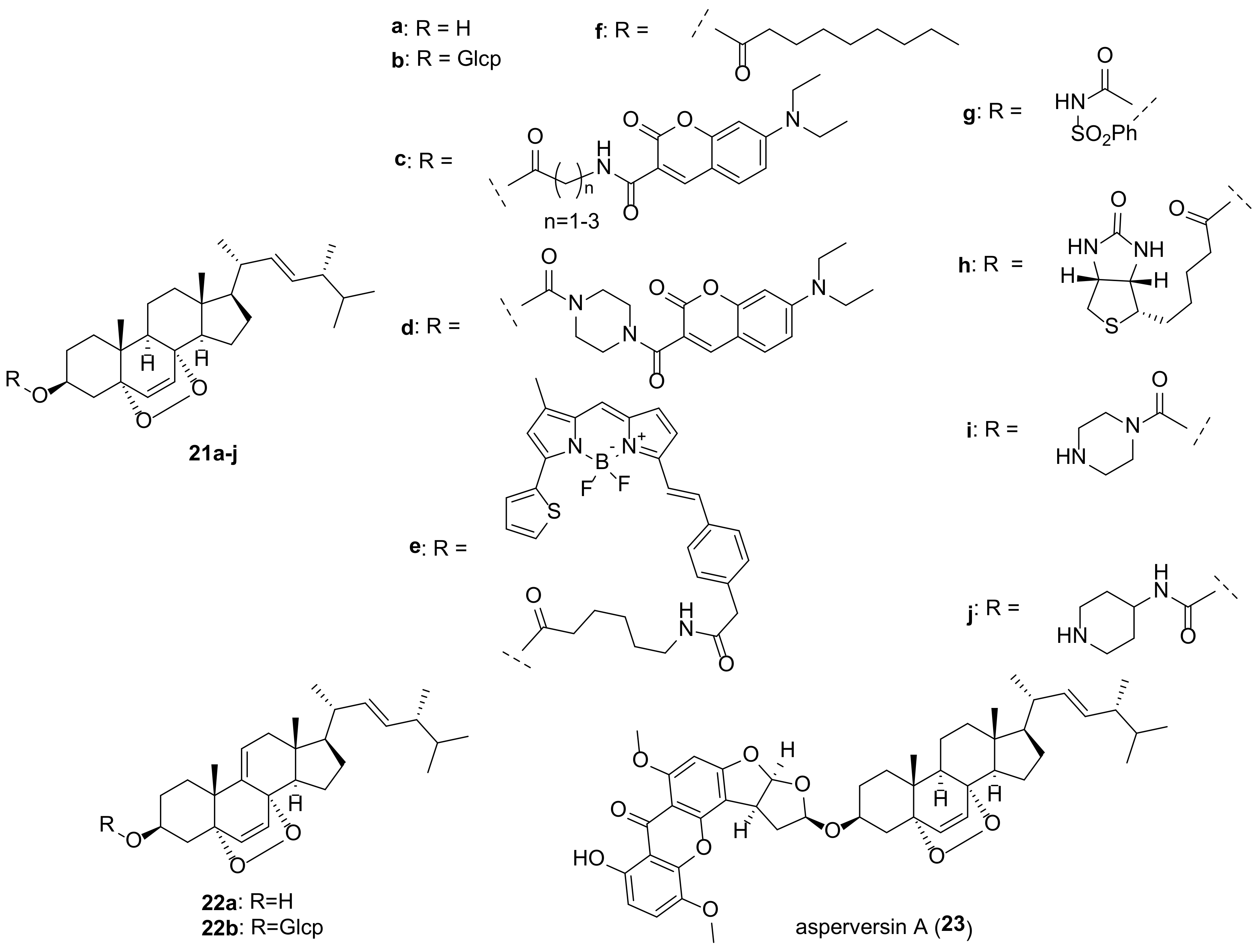
In an attempt to improve antitumor activity, a number of derivatives of endoperoxide
21a have been studied. Ergosterol peroxide sulfonamide
21g was found to be more effective in reducing cancer cell viability than the parental endoperoxide
21a [
150]. Significantly, its toxicity to normal human BJ fibroblasts was minimal, indicating that
21g targets cancer cells.
A series of EP analogs containing BODIPY or a biotin moiety was obtained by Rivas et al. as probes for cellular localization studies [
149]. They demonstrated that EP is distributed across the cytosol with significant accumulation in the endoplasmic reticulum. In addition, the resulting compounds were tested for antiproliferative activity in breast cancer cell models. The most active ones were analogs
21e and
21h (
Table 1).
Several adducts of EP with 7-
N,
N-diethylaminocoumarins have been obtained by Bu et al. [
145]. Analogues
21c and
21d exhibited increased cytotoxicity compared to
21a, which was explained by their localization mainly in mitochondria, as followed from fluorescence imaging. In addition, the piperazine derivative
21d suppressed the formation, invasion, and migration of cell colonies, induced arrest of HepG2 cells in the G2/M phase, and increased the level of intracellular reactive oxygen species.
A number of EP 3-carbamate derivatives were obtained by Hu et al. [
146]. They exhibited antiproliferative activity, which was 6–28 times stronger than that of the initial endoperoxide
21a (
Table 1). The most active compounds
21i and
21j contain piperazinyl and piperidinyl fragments.
A steroid-xanthone heterodimer, asperversin A (
23), was isolated from the culture of
Aspergillus versicolor, an endophytic fungus isolated from the marine brown alga
Sargassum thunbergii [
179]. Compound
23 was tested for biological activities against some bacterial and fungal strains with no noticeable effect.
Further structural modifications of steroidal molecule with retention of the 5α,8α-endoperoxide scaffold included changes in the carbon skeleton of the side chain [
180,
181]. Thus, 7-dehydrocholesterol peroxide, its acetate and hemisuccinate showed improved anticancer activity and selectivity over the corresponding derivatives of EP [
180].
In comparison with the compounds
21a and
22a, 5α,9α-endoperoxides have been studied much less due to their lower availability. Compounds
24 and
25 (
Figure 6) were isolated from the edible mushroom
Grifola gargal and evaluated in the osteoclast-forming assay [
182]. They inhibited osteoclast formation, which may be of interest for the prevention of osteoporosis. Endoperoxide
26, isolated from the fruiting bodies of
Stropharia rugosoannulata, protected neuronal cells by attenuating endoplasmic reticulum stress caused by thapsigargin, an inhibitor of the Ca
2+-ATPase [
81]. A significant cytotoxicity (
Table 1) against A549 and MCF-7 cells was noted for the endoperoxide
27, isolated from the fruiting body of a medicinal macro fungus
Ganoderma lingzhi [
12]. Agarol (
28) was isolated as a tumoricidal substance from the mushroom
Agaricus blazei [
157]. Its cytotoxicity was evaluated against four cancer lines (
Table 1). Agarol (
28) was shown to induce apoptosis by increasing generation of ROS and release of apoptosis-inducing factor from the mitochondria to the cytosol.
4. Epoxides
The majority of compounds of this group are 5α,6α epoxides (
Figure 7). Almost all of them contain a hydroxy- or keto group at C-7, Δ
8(9)-, or Δ
8(14)-double bond, and some 5α,6α-epoxides have a functionalized ring D. Other epoxides (4,5-, 5β,6β-, 8,9-, 8,14-, and 9,11-derivatives) are much less common in fungi (
Figure 8). Compounds
29–
59 were tested in various assays, including AChE inhibitory, cytotoxic, α-glucosidase inhibition, NO production inhibition, etc., (
Table 2).
Bae et al. noted that the presence of an epoxy group in the tetracyclic skeleton of ergosterol derivatives increases their cytotoxic properties [
183]. Isolation of a series of 5α,6α-epoxides from the macrofungus
Omphalia lapidescens allowed to establish some structure activity relationship correlations [
15]. The greatest cytotoxicity against a human gastric cancer cell line, HGC-27, was noted for the compound
30a and
31a containing an α-oriented hydroxyl group at C-7 and Δ
8(9)- or Δ
8(14)-double bond (
Table 2). The transition to 7-ketones
33 and
36 led to a decrease in activity, and of both compounds, the derivative
33 without a double bond in the BC cycles was less active. The diepoxide
52 showed the least activity, which indicates the importance of the double bond for cytotoxic activity.
Epoxides
41,
43a, and
43b, isolated from the culture of Basidiomycete
Polyporus ellisii, were evaluated for cytotoxicity against five human cancer cell lines [
184]. The first two compounds were practically inactive, while epoxide
41 exhibited strong activity against all tested cell lines with IC
50 in the range from 1.5 to 3.9 μM (
Table 2).
Ferreira et al. performed virtual screening experiments on low-molecular weight fungal constituents as potential MDM2 inhibitors [
185]. The latter is an important negative regulator of the p53 tumor suppressor, and its inhibitors have significant anti-tumor activity. From the compounds studied, epoxide
29 returned one of the best docking scores.
Epoxide
31b was found to exhibit potent inhibitory activity on the expression of mRNA of proprotein convertase subtilisin/kexin type 9 (PCSK9) [
186]. The latter affects the low density lipoprotein receptor on the surface of liver cells, resulting in high level of low density lipoprotein cholesterol (LDL-C). PCSK9 inhibitors have been proposed as novel LDL-C-lowering agents for the treatment of hyperlipidemia. Compound
31b showed activity with IC
50 values of 8.23 μM, which was comparable with berberine (IC
50 8.04 μM) used as a positive control.
A number of epoxides were tested for their anti-inflammatory activity. As a rule, inhibition of TNF-α and NO production in LPS-stimulated RAW264.7 macrophage cells was used to evaluate anti-inflammatory effects. Epoxide
30c showed superior inhibitory activity on NO production with IC
50 value of 3.24 μM [
103]. In the same experiment, the positive control L-NMMA, nitric oxide synthase inhibitor, revealed IC
50 value of 49.86 μM. TNF-α secretion decreased after treatment of macrophage cells with epoxide
49, which at 10 μM exhibited activity with inhibition value of 37.5% [
187]. This was comparable to the positive control (52.5% at 1 μM) exerted by celecoxib, the cyclooxygenase-specific inhibitor.
Table 2.
Sources and biological activity of fungal epoxides.
Table 2.
Sources and biological activity of fungal epoxides.
| Compound | Fungal Source [Ref.] | Assays (Activity) [Ref.] |
|---|
| 29 | Hericium erinaceus [187,188], Chaetomium sp. M453 [189], Colletotrichum sp. YMF432 [190], Cordyceps sinensis [191], Stereum insigne CGMCC5.57 [79] | AChE inhibitory assay (IC50 67.8 μM) [190], nematicidal and antibacterial assays (no activity) [79] |
| 30a | Amauroderma subresinosum [83], Ganoderma lucidum [147], G. resinaceum [103], Grifola frondosa [154], Omphalia lapidescens [15], Simplicillium sp. YZ-11 [192], Stropharia rugosoannulata [193], Pleurotus eryngii [6] | α-glucosidase inhibition assay (IC50 > 100 μM) [154], cytotoxic assay (HGC-27, IC50 11.69 μM) [15], (MCF-7, IC50 24.3 μM; NCI-H460, IC50 19.8 μM; SF-268, IC50 15.5 μM) [194], (A549, IC50 35.99 μM; HepG2, IC50 25.81 μM; MDA-MB-231, IC50 29.73 μM) [154], (HepG2, IC50 22.1 μM; MDA-MB-231, IC50 20.3 μM) [147], lettuce hypocotyl growth assay (65–80% inhibition) [193], NO production inhibition assay (IC50 12.4 μM) [6], (IC50 19.77 μM) [103] |
| 30b | Ganoderma resinaceum [103], Stropharia rugosoannulata [81] | anti-fungal assay (MIC 250 μM) [81], NO production inhibition assay (IC50 17.23 μM) [103], osteoclast-forming assay [81] |
| 30c | Amauroderma amoiensis [82], Ganoderma resinaceum [103] | NO production inhibition assay (IC50 3.24 μM) [103] |
| 31a | Cortinarius glaucopus [195], Ganoderma lucidum [147], G. resinaceum [103], G. sinense [196], Grifola frondosa [154], Hygrophorus russula [183], Leptographium qinlingensis [197], Omphalia lapidescens [15], Simplicillium sp. YZ-11 [192], Stropharia rugosoannulata [193], Phellinus linteus [198], Pleurotus eryngii [6], Termitomyces microcarpus [132] | α-glucosidase inhibition assay (IC50 > 100 μM) [154], cytotoxic assay (HGC-27, IC50 18.97 μM) [15], (MCF-7, IC50 > 50 μM; NCI-H460, IC50 > 50 μM); SF-268, IC50 > 50 μM)-194], (A549, IC50 69.11 μM; HepG2, IC50 38.87 μM; MDA-MB-231, IC50 46.76 μM) [154], (A549, IC50 15.3 μg/mL; XF498, IC50 15.1 μg/mL) [183], (HepG2, IC50 50.6 μM; MDA-MB-231, IC50 46.7 μM) [147], HNE inhibitory assay (IC50 28.2 μM) [198], lettuce hypocotyl growth assay (61–78% inhibition) [193], NCI 60 panel [132], NO production inhibition assay (IC50 > 30 μM) [6], (IC50 23.34 μM) [103], (IC50 > 40 μM) [196] |
| 31b | Ganoderma resinaceum [103], Hericium erinaceus [187,188], Sparassis crispa [186,199], Phellinus linteus [198], Pleurotus eryngii [6] | cytotoxic assay (MCF-7, IC50 > 50 μM) [194], (NCI-H460, IC50 > 50 μM) [194], (SF-268, IC50 > 50 μM) [194], NO production inhibition assay (IC50 14.3 μM) [6], (IC50 17.23 μM) [103], PCSK9 mRNA expression (inhibition, IC50 8.23 μM) [186] |
| 31c | Hericium erinaceum [200] | PPAR transactivation assay (EC50 8.2 μM) [200] |
| 31d | Hericium erinaceum [200] | PPAR transactivation assay (EC50 6.4 μM) [200] |
| 32 | Pleurotus eryngii [59] | aromatase inhibitory assay (IC50 17.3 μM) [59] |
| 33 | Hericium erinaceum [187], Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 29.34 μM) [15], HNE inhibitory assay (IC50 75.1 μM) [198], TNF-α secretion assay (inhibition value of 37.5% at 10 μM) [187] |
| 34 | Grifola gargal [182] | osteoclast-forming assay [182] |
| 35 | Amauroderma subresinosum [83] | AChE inhibitory assay (20.9% at 100 μM) [83] |
| 36 | Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 23.41 μM) [15] |
| 37a | Pleurotus eryngii [201] | cytotoxic assay (RAW264.7, IC50 > 30 μM) [201] |
| 37b | Stropharia rugosoannulata [81] | osteoclast-forming assay [81] |
| 38 | Grifola gargal [182] | cytotoxic assay (HepG2, IC50 200.9 μM; MDA-MB-231, IC50 189.4 μM) [147], osteoclast-forming assay [182] |
| 39 | Amauroderma subresinosum [83], Polyporus ellisii [184] | cytotoxic assay (HL-60, IC50 32.1 μM; SMMC-7721, A549, MCF-7, SW480, IC50 > 40 μM) [184] |
| 40 | Pleurotus eryngii [201] | cytotoxic assay (RAW264.7, IC50 > 30 μM) [201], NO production inhibition assay (IC50 13.2 μM) [201] |
| 41 | Polyporus ellisii [184] | cytotoxic assay (HL-60, IC50 1.5 μM; SMMC-7721, IC50 3.9 μM; A549, IC50 2.7 μM; MCF-7, IC50 3.1 μM; SW480, IC50 2.9 μM) [184] |
| 42 | Phomopsis sp. [202] | α-glucosidase inhibition assay (IC50 > 100 μM) [202] |
| 43a | Polyporus ellisii [184], Phomopsis sp. [202] | antibacterial assay (MIC 28.2 μM against Micrococcus tenuis) [202], cytotoxic assay (HL-60, IC50 32.1 μM; SMMC-7721, A549, MCF-7, SW480, IC50 > 40 μM) [184] |
| 43b | Ganoderma resinaceum [103], Polyporus ellisii [184], Phomopsis sp. [202] | cytotoxic assay (HL-60, IC50 18.8 μM; SMMC-7721, A549, MCF-7, SW480, IC50 > 40 μM) [184] |
| 44 | Grifola gargal [182] | osteoclast-forming assay [182] |
| 45 | Pleurotus eryngii [6] | NO production inhibition assay (IC50 > 30 μM) [6] |
| 46 | Ganoderma lucidum [147] | cytotoxic assay (HepG2, IC50 138.3 μM; MDA-MB-231, IC50 176.1 μM) [147] |
| 47 | Amauroderma amoiensis [82] | AChE inhibitory assay (14.63% inhibition at 100 μM) [82] |
| 48 | Trametes versicolor [168] | (NO inhibitory activity at 12.5 μM, IL-6 inhibitory effect at 25 μM) [168] |
| 49 | Hericium erinaceus [187,188] | TNF-αsecretion assay (37.5% inhibition at 10 μM) [187] |
| 50 | Hericium erinaceus [187,188], Phellinus linteus [198], Stropharia rugosoannulata [193] | HNE inhibitory assay (IC50 35.2 μM) [198], inhibition of lettuce hypocotyl growth (no activity) [193] |
| 51 | Ganoderma lucidum [147], Hericium erinaceum [187] | NO production inhibition assay (moderate activity) [187] |
| 52 | Aspergillus awamori [203], Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 58.43 μM) [15], (A549, IC50 64 μM) [203] |
| 53 | Hericium erinaceum [187], Pleurotus eryngii [6] | NO production inhibition assay (IC50 > 30 μM) [6] |
| 54 | Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 15.37 μM) [15] |
| 55 | Pleurotus eryngii [201] | cytotoxic assay (RAW264.7, IC50 > 30 μM) [201] |
| 56 | Talaromyces stipitatus [204] | cytotoxic assay (Hep3B, IC50 4.75 μM; HepG2, IC50 8.85 μM; Huh-7, IC50 13.78 μM) [204] |
| 57 | Aspergillus penicillioides [205], Ganoderma lingzhi [12] | antibacterial assay (MIC 32 μg/mL against Vibrio anguillarum) [205], cytotoxic assay (A549, IC50 8.57 μM; MCF-7, IC50 6.09 μM) [12] |
| 58 | Chaetomium sp. [189] | AChE inhibitory assay (20–60% inhibition at 50 μg/mL) [189] |
| 59 | Colletotrichum sp. [206] | AChE inhibitory assay (18.2% inhibition at 100 μg/mL) [206] |
Human neutrophil elastase (HNE) is a serine protease that can degrade extracellular matrix proteins such as collagen, fibronectin, etc. Inhibition of this enzyme can prevent the loss of skin elasticity, thereby preventing skin aging. Yoo et al. reported the HNE-inhibitory properties of
Phellinus linteus mycelium components [
198]. All three tested epoxides
31a,
34, and
50 showed significant activity with ID
50 ranging from 28.2 to 75.1 μM.
Epoxides
30a,
31a, and
33 were isolated after anaerobic incubation of ergosterol peroxide (EP,
21a) with rat intestinal flora [
207]. Two of them (
30a and
33) were found to be more active against human colorectal cancer cells than the original EP. This means that EP’s strong anti-tumor properties may be (at least in part) due to its metabolic products.
A number of ergostane-type sterol fatty acid esters, including epoxides
31c and
31d, were isolated from the mushroom
Hericium erinaceum and evaluated for their PPAR transactivational effects using a luciferase reporter system [
200]. Oleyl and linoleyl esters
31c and
31d proved to be the most potent activators of the transcriptional activity of PPARs with EC
50 values down to 6.4 μM.
5. Polyols
It should be kept in mind that the structures of ergostane-type steroids with hydroxyl and/or carbonyl group(s) given below do not fully reflect their diversity in fungal sources. A large number of compounds have been isolated before 2010; for a number of compounds isolated later, no data on biological activity are given, and for this reason they are not included in this review.
Many fungal ergostanes of this class are 5α-alcohols containing (an)other hydroxy (or a functionalized hydroxy) group(s) at C-6, C-9, and/or C-14 (
Figure 9). 5α,6α Epoxides are their evident biosynthetic precursors. As a rule, rings A and B are trans-fused for most ergostanes of this group, with the exception of 5β-alcohols
77, 78, 84 (
Figure 10). It should be noted that fomentarol B (
84) has a cis-junction of ring B and C, which is rare among the ergostane type steroids [
208].
Cerevisterol (
60) is probably the best studied among 5α,6β-dihydroxy derivatives, as it is widespread in the fungal kingdom (
Table 3). It should be noted that data on its cytotoxicity are inconsistent and sometimes contradictory. Thus, cerevisterol (
60) showed significant activity with IC
50 values of 1.1–1.9 μM against the BT-549, KB, SK-MEL, and SKOV-3 cancer cell lines [
209]. On the other hand, it was practically inactive toward A549, HeLa, HepG2, and MCF-7 cells [
210]. This inconsistence may be partly due to the diverse cell lines used by different authors. But a large difference was also observed in experiments with the same cell lines (e.g., reported IC
50 values for HepG2 varied from 14.5 μM [
211] to 174.6 μM [
147]).
The results of studies of antimicrobial activity also vary quite a lot. Thus, in the course of searching for biologically active constituents of wood decaying mushrooms,
Trametes gibbosa and
Trametes elegans, Agyare et al. isolated cerevisterol (
60) as a compound responsible for their antimicrobial activity [
212]. It inhibited the growth of a number of bacteria with MICs ranging from 25 to 50 µg/mL (ciprofloxacin MICs were between 0.31 and 3.50 µg/mL). The sub-inhibitory concentration of
60 (3 µg/mL) modified the activity of commonly used antibiotics (either potentiating or reducing). Similar results with respect to antimicrobial activity of
60 were obtained by Zhou et al. [
213]. On the other hand, no antimicrobial activity for cerevisterol (
60) was reported in works [
214,
215].
To access the anti-inflammatory activity of cerevisterol (
60), Lee et al. measured the levels of NO and PGE
2 and the production of cytokines TNF-α, IL-1, and IL-6 in LPS-stimulated macrophages [
216]. It was shown that
60 suppressed the LPS-induced production of NO and PGE2 and decreased the expression of pro-inflammatory cytokines.
Table 3.
Sources and biological activity of fungal alcohols.
Table 3.
Sources and biological activity of fungal alcohols.
| Compound | Fungal Source [Ref.] | Assays (Activity) [Ref.] |
|---|
| 60 | Aspergillus fumigatus [213], A. versicolor [179], Cladosporium sp. [217], Clitocybe nebularis [214], Eurotium rubrum [80], Fomes fomentarius [208], Fusarium chlamydosporum [209,218], F. equiseti [219], F. solani [216], Ganoderma sinense [196,220], Glomerella sp. [215], Gomphus clavatus [221], Hericium erinaceum [222,223], Hypholoma lateritium [224], Lentinus polychrous [225], Leptographium qinlingensis [197], Leucocalocybe mongolica [210], Meripilus giganteus [91], Morchella esculenta [226], Omphalia lapidescens [15], Penicillium brasilianum [227], Pleurotus eryngii [6], P. tuber-regium [228], Polyporus umbellatus [77,211], Termitomyces microcarpus [132], Trametes gibbosa and T. elegans [212], Tricholoma populinum [229], Xylaria nigripes [105] | AChE inhibitory assay (0.4% inhibition at 100 μg/mL) [80], antibacterial assay (no activity against Streptococcus agalactiae, Staphylococcus epidermidis, Moraxella catarrhalis, Haemophilus influenzae, and Proteus mirabilis) [214], (S. typhi, S. aureus and A. niger, MICs 25 μg/mL each, E. faecalis, MIC 50 μg/mL) [212], (Bacillus subtilis and Escherichia coli, MICs 64 μg/mL each; Staphylococcus aureus, MIC 32 μg/mL) [213], cytotoxic assay (A549, IC50 94.75 μM; HeLa, IC50 74.13 μM; HepG2, IC50 46.58 μM; MCF-7, IC50 63.76 μM) [210], (T47D, 50.2% inhibition at 30 μM) [229], (BT-549, 1.4 μM; KB, 1.90 μM; SK-MEL, 1.70 μM; SKOV-3, 1.1 μM) [209], (Caco-2, IC50 37.56 μM; MCF-7, IC50 32.4 μM; MDA-MB-231, IC50 41.5 μM) [219], (HGC-27, IC50 37.71 μM) [15], (MCF-7, IC50 37.2 μM; PC-3, IC50 80 μM) [221], (HepG2, IC50 14.5 μM) [211], (HepG2, IC50 174.6 μM; MDA-MB-231, IC50 148.8 μM) [147], (SW1990, IC50 32.81 μM; Vero, IC50 > 100 μM) [220], NF-κB inhibitory assay (IC50 5.1 μM) [226], HIV-inhibitory assay (IC50 9.3 μM) [230], HNE inhibitory assay (IC50 77.5 μM) [198], DPPH free radical-scavenging assay (IC50 11.38 μM) [222], GIRK channel inhibitory assay (13% inhibition at 10 μM) [224], lipoxygenase inhibitory assay (IC50 5.46 μM) [218], NO production inhibition assay (IC50 > 40 μM) [196], (IC50 > 30 μM) [6], ORAC assay (antioxidant activity 1.94 mmol TE/g) [91], PTP1B inhibitory activity assay (IC50 7.5 μg/mL) [77], toxicity to Pinus armandi seedlings assay (lethal rate 95% at 30 μg/mL) [197], trap activity assay (reduction to 28.1% from 332% in control cells) [223] |
| 61a | Aspergillus penicillioides [205], A. ustus [231], Aspergillus versicolor [179], Eurotium rubrum [80], Ganoderma lucidum [232], G. sinense [233], Hericium erinaceum [223], Omphalia lapidescens [15], Penicillium brasilianum [227], Pleurotus eryngii [6], Tricholoma populinum [229], Xylaria nigripes [105] | AChE inhibitory assay (2.7% inhibition at 100 μg/mL) [80], cytotoxic assay (T47D, 23.7% inhibition at 30 μM; MDA-MB-231, 54.7% inhibition at 30 μM) [229], (U2OS, IC50 6.0 μM) [105], (HGC-27, IC50 4.17 μM) [15], [15], (HL-60, IC50 22.4 μM; LLC, IC50 55.3 μM; MCF-7, IC50 > 100 μM) [232], HIV-inhibitory assay (IC50 3.8 μM) [230], HNE inhibitory assay (IC50 14.6 μM) [198], neuroprotective activity assay (20.9% increase in cell viability against Aβ25-35-induced injury in SH-SY5Y neuroblastoma cells at the concentration 10 μM) [105], NO production inhibition assay (IC50 20.4 μM) [6], (108.2% inhibitory rate at 10 μM) [230], trap activity assay (reduction to 74.8% from 332% in control cells) [223] |
| 61b | Fomes fomentarius [208], Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 25.50 μM) [15] |
| 61c | Eurotium rubrum [80], Hericium erinaceum [223] | AChE inhibitory assay (17.9% inhibition at 100 μg/mL) [80], trap activity assay (reduction to 81.8% from 332% in control cells) [223] |
| 61d | Fusarium chlamydosporum [218] | lipoxygenase inhibitory assay (IC50 3.06 μM) [218] |
| 61e | Hericium erinaceum [223] | ORAC assay (antioxidant activity 8.01 mmol TE/g at 10 μM) [223] |
| 62a | Eurotium rubrum [80], Fomes fomentarius [208], Hericium erinaceum [223], Hygrophorus russula [183], Omphalia lapidescens [15] | AChE inhibitory assay (2.4% inhibition at 100 μg/mL) [80], cytotoxic assay (HGC-27, IC50 > 100 μM) [15], (HepG2, IC50 196.9 μM; MDA-MB-231, IC50 114.2 μM) [147], (A549, >30 μg/mL; XF498, >30 μg/mL) [183], trap activity assay (reduction to 138.9% from 332% in control cells) [223] |
| 62b | Hericium erinaceum [200] | PPAR transactivation assay (EC50 18.7 μM) [200] |
| 62c | Hericium erinaceum [200] | PPAR transactivation assay (EC50 20.6 μM) [200] |
| 63a | Ganoderma lucidum [147], Pleurotus eryngii [6] | cytotoxic assay (HepG2, IC50 62.5 μM; MDA-MB-231, IC50 56.3 μM) [147], NO production inhibition assay (IC50 29.8 μM) [6] |
| 63b | Ganoderma sinense [220] | cytotoxic assay (SW1990, IC50 5.05 μM; Vero, IC50 22.59 μM) [220] |
| 64 | Fomes fomentarius [208], Ganoderma lucidum [147], Hericium erinaceum [187] | cytotoxic assay (HepG2, IC50 156.4 μM; MDA-MB-231, IC50 168.9 μM) [147], TNF-α secretion assay (33.7% inhibition at 10 μg/mL) [187] |
| 65 | Clitocybe nebularis [214], Fomes fomentarius [208], Hericium erinaceum [223], Hygrophorus russula [183], Leptographium qinlingensis [197], Naematoloma fasciculare [151], Stropharia rugosoannulata [81], Tricholoma populinum [229] | antibacterial assay (no activity against Streptococcus agalactiae, Staphylococcus epidermidis, Haemophilus influenzae, and Proteus mirabilis, marginal activity against Moraxella catarrhalis) [214], anti-fungal assay (MIC 500 μM) [81], cytotoxic assay (MCF-7, MDA-MB-231, T47D, no activity) [229], (HepG2, IC50 129.7 μM; MDA-MB-231, IC50 148.2 μM) [147], (A549, 17.1 μg/mL; XF498, 16.5 μg/mL) [183], (A549, 10.83 μM; HCT-15, 13.2 μM; SK-MEL-2, 10.39 μM; SK-OV-3, 12.16 μM;) [151] |
| 66 | Ganoderma lucidum [147] | cytotoxic assay (HepG2, IC50 286.4 μM; MDA-MB-231, IC50 216.5 μM) [147] |
| 67a | Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 12.71 μM) [15], (HepG2, IC50 184.6 μM; MDA-MB-231, IC50 224.2 μM) [147] |
| 67b | Hericium erinaceum [200] | PPAR transactivation assay (EC50 22.3 μM) [200] |
| 68a | Omphalia lapidescens [15] | cytotoxic assay (HGC-27, IC50 26.74 μM) [15] |
| 68b | Fomes fomentarius [208] | cytotoxic assay (A549, IC50 29.8 μM; MCF-7, IC50 26.1 μM; NUGC-3, IC50 24.1 μM) [208] |
| 69 | Pleurotus eryngii [6] | NO production inhibition assay (IC50 > 30 μM) [6] |
| 70 | Hericium erinaceus [187,188] | TNF-α secretion assay (25% inhibition at 10 μg/mL) [187] |
| 71 | Penicillium granulatum [234] | cytotoxic assay (no activity) [234] |
| 72 | Hericium erinaceum [187] | TNF-α secretion assay (36.7% inhibition at 10 μg/mL) [187] |
| 73 | Coprinus setulosus [101], Ganoderma lipsiense [235], G. resinaceum [103], Xylaria nigripes [105] | antigiardial assay (93.6% inhibition against Giardia duodenalis throphozoites) [235], NO production inhibition assay (IC50 27.6 μM) [105], (IC50 22.76 μM) [103], tyrosinase inhibitory assay (IC50 6.9 μM) [236] |
| 74 | Eurotium rubrum [80] | AChE inhibitory assay (23.1% inhibition at 100 μg/mL) [80] |
| 75 | Ganoderma resinaceum [103] | NO production inhibition assay (IC50 22.76 μM) [103] |
| 76 | Penicillium granulatum [234] | cytotoxic assay (no activity) [234] |
| 77 | Omphalia lapidescens [16] | cytotoxic assay (GES-1, IC50 > 50 μM; HGC-27, IC50 12.28 μM; MDA-MB-231, IC50 11.33 μM) [16] |
| 78 | Omphalia lapidescens [16], Pleurotus eryngii [6] | cytotoxic assay (GES-1, IC50 28.0 μM; HGC-27, IC50 > 50 μM; MDA-MB-231, IC50 24.85 μM) [16], NO production inhibition assay (IC50 > 30 μM) [6] |
| 79 | Ganoderma duripora [237], Ganoderma lucidum [232,238], Phellinus linteus [198] | cytotoxic assay (HL-60, IC50 12.7 μM; LLC, IC50 45.2 μM; MCF-7, IC50 > 100 μM) [232], (A549, MCF-7, PC-3, IC50 > 50 μM) [238], HNE inhibitory assay (IC50 > 100 μM) [198] |
| 80 | Lasiodiplodia pseudotheobromae [11] | AChE inhibitory assay (no activity) [11], α-glucosidase inhibition assay (no activity) [11] |
| 81 | Penicillium granulatum [234] | cytotoxic assay (A549, IC50 5.5 μM) [234] |
| 82 | Penicillium granulatum [234] | cytotoxic assay (A549, BEL-7402, SHG-44, IC50 > 20 μM; ECA-109, IC50 9.2 μM; HepG2, IC50 7.0 μM) [234] |
| 83 | Omphalia lapidescens [16] | cytotoxic assay (GES-1, HGC-27, MDA-MB-231, IC50 > 50 μM) [16] |
| 84 | Fomes fomentarius [208], Omphalia lapidescens [16] | cytotoxic assay (MDA-MB-231, IC50 140.86 μM) [16], NO production inhibition assay (98.77% inhibitory activity at 50 μM) [208] |
| 85 | Penicillium chrysogenum [239], Penicillium granulatum [240] | anti-fungal assay (8 mm diameter at 20 μg/disk) [239], cytotoxic assay (HeLa, IC50 15 μg/mL; NCI-H460, IC50 40 μg/mL; SW1990, IC50 31 μg/mL) [239], (HepG2, IC50 8.2 μM) [240] |
| 86 | Penicillium granulatum [234] | cytotoxic assay (no activity) [234] |
| 87 | Penicillium granulatum [234] | cytotoxic assay (A549, IC50 8.0 μM; BEL-7402, IC50 8.5 μM; ECA-109, IC50 8.3 μM; HepG2, IC50 6.7 μM; SHG-44, IC50 4.8 μM) [234] |
| 88 | Penicillium granulatum [234] | cytotoxic assay (no activity) [234] |
Yoo et al. studied the HNE-inhibitory potency of ergostanes isolated from the mycelium of
Phellinus linteus [
198]. Methyl ether
61a revealed the highest activity among all tested compounds with an IC
50 14.6 μM, which was comparable with the positive control (epigallocatechin gallate, IC
50 12.5 μM). The corresponding alcohol
60 was five times less active than
61a.
Kim et al. studied the inhibitory activity of steroids isolated from
Hericium erinaceum against tartrate-resistant acid phosphatase (TRAP) [
223]. The latter has become a promising target for the development of new therapeutics for the treatment of osteoporosis and other bone-related diseases. Compounds
60, 61a, 61c, 62a at a concentration of 10 μM reduced TRAP activity in osteoclasts differentiated from RAW 264.7 cells, from 322% in control cells to 28–139% in treated cells.
Compared to 5α,6-diols, other fungal polyols (
Figure 10) have been relatively less studied. As mentioned above, many ergostane steroids are found in both mushrooms and plants. In particular, this applies to triol
73 found in various fungal species [
101,
103,
105,
235]. Among sixty-three compounds isolated from bamboo
Sinocalamus affinis and studied as inhibitors of estrogen biosynthesis, triol
73 showed the highest activity with an IC
50 value of 0.5 μM [
241]. It reduced the level of expression of aromatase mRNA in granulosa-like cells of human ovaries without affecting the catalytic activity of aromatase. This discovery makes the steroid
73 an interesting lead compound in the development of new agents for the treatment of estrogen-dependent cancers.
Studying the cytotoxicity of compounds isolated from the fruiting bodies of a medicinal mushroom
Ganoderma lucidum, Min et al. selected the 2β,3α,9α-triol
79 for a more detailed evaluation [
232]. Treatment with
79 in a dose-dependent manner inhibited the growth of HL-60 human premyelocytic leukemia cells with the IC
50 value of 12.7 μg/mL. The effect was attributed to the induction of the apoptotic process, including activation of DNA fragmentation and caspase-3 activity.
7. Ketones
Most compounds of this group of ergostane-type steroids contain keto functions at C-3 and C-6, as well as a number of double bonds (
Figure 14). Ergone (
124) is probably the best studied among them [
256]. It is found in many fungal sources (
Table 5), usually with a content of less than 10 μg/g of mushroom fruit bodies.
Polyporus umbellatus, in comparison with other mushrooms, contains the highest amount of this compound, which, under optimized conditions, can reach 86.9 μg/g [
257]. For practical purposes, ergone (
124) can be easily obtained through a three-step chemical synthesis from ergosterol [
258]. Ergone has been reported to possess various activities (
Table 5), including cytotoxic, anti-bacterial [
205], anti-inflammatory [
228,
259], anti-malarial [
249], diuretic [
260] abilities, and protective effects of early renal injury [
261,
262].
Attempts were made to study the mechanism of its action. A strong anticancer effect of
124 to HepG2 cells was associated with the induction of G2/M cell cycle arrest and apoptosis in a caspase-dependent manner [
270].
Wang et al. studied the effect of ergone (
124) on lipopolysaccharide-induced acute lung injury [
272]. Pretreatment of mice with
124 was found to reduce neutrophil recruitment, regulate the release of inflammatory cytokines, reduce pulmonary edema, and correct pulmonary insufficiency. The observed effects were associated with inhibition of the NLRP3 signaling pathway.
Ergone (
124) was found to inhibit signaling pathways STAT3 and Src in head and neck cancer-initiating cells [
263] that results in the reduction of their stemness properties and tumorigenicity and is of interest for the treatment of head and neck squamous cell carcinoma.
The variety of pharmacological activities prompted scientists to study pharmacokinetic properties of ergone. Fan et al. investigated the interactions between ergone and human serum albumin [
273]. The latter is a carrier protein for many endogenous and exogenous molecules in blood and greatly affects the pharmacokinetics of drugs. Fluorescence spectroscopy revealed the binding of ergone to albumin, in which hydrogen bonds and hydrophobic interactions play a dominant role.
The following pharmacokinetic parameters were measured after a single oral administration (20 mg/kg) of ergone to rats: the area under the plasma concentration versus time curve from time 0 h to indefinite time (AUC
0–∞) was 19.6 μg h mL
−1, peak plasma concentration (C
max) was 1.5 μg/mL, the elimination half-life (t
1/2) was 5.90 h, and time to C
max (T
max) was 3.8 h [
266].
To improve the therapeutic effect of ergone, several drug delivery systems has been proposed [
274,
275]. The folate receptor is known to be overexpressed in a wide variety of cancers, which is the basis for the development of tumor-targeted drug delivery systems. One of them uses the most abundant protein in plasma, albumin. Folate-modified ergone bovine serum albumin nanoparticles showed increased cellular uptake, targeting ability and cytotoxicity toward KB cells [
274]. An in vivo experiment showed a higher antitumor effect and less toxicity of ergone nanoparticles compared to free ergone. Another delivery system was based on the encapsulation of ergone in PEGylated liposomes [
275]. Pharmacokinetic studies have shown that encapsulation provides a longer residence time of ergone in the blood, which leads to a more effective in vivo antitumor effect.
9. Ergostanes with a Rearranged Tetracyclic Skeleton
Due to their intriguing structural complexity and promising biological activities, ergostanes with a rearranged tetracyclic carbon skeleton have become very attractive targets for chemists and biologists. A recent review [
23] has covered this area quite thoroughly, but for consistency and completeness some results will be briefly discussed here.
Most ergostanes with a modified skeleton are highly functionalized compounds bearing three and more functional groups. A certain exception are aromatic 1(10→6)abeo-ergostane-type steroids
157–
160 (
Figure 17). Two of them,
157 and
158, exhibited significant cytotoxicity toward murine colorectal CT26 and human leukemia K562 cancer cell lines (
Table 7). Citreoanthrasteroid B (
158) was also tested for the neuroprotective effects on PC12 cells injured by glutamate (15 mM) [
288]. Compound
158 showed potential neuroprotective activities by inhibiting the death of injured PC12 cells with EC
50 value of 24.2 μM.
Another 1(10→6)abeo-steroid, aspersteroid A (
161), was isolated from the culture extract of
Aspergillus ustus [
280]. It exhibited moderate cytotoxicity on four cancer cell lines, antimicrobial activity against Gram-negative and Gram-positive bacteria and immunosuppressive activities against the proliferation of T and B lymphocyte cells in vitro (
Table 7).
Three anthrasteroid glycosides, malsterosides A-C (
162a–
c), were isolated from the fungus
Malbranchea filamentosa [
289]. The sugar moiety in the side chain of all glycosides was found to be D-mannose and the glycoside
162c contained N-acetyl-D-glucosamine at the C-3 position. Cytotoxicity studies were performed with the A549 and Hela cancer cell lines. A moderate cytotoxicity in both lines was noted for malsteroside A (
162a).
Two 1(10→6)-abeo-14,15-secosteroids, asperfloketals A (
163) and B (
164), were found in the sponge-associated fungus
Aspergillus f locculosus 16D-1 [
290]. They exhibited no cytotoxicity against three tested cancer cell lines. Promising results were obtained in anti-inflammatory assays. Compounds
163 and
164 displayed stronger activity in the CuSO
4-induced transgenic fluorescent zebrafish than ibuprofen used as a positive control.
A-nor-B-homo steroid
165 (
Figure 18) containing a 10(5→4)-abeo-ergostane fragment was isolated from culture of basidiomycete
Polyporus ellisii [
184] and from the mangrove-derived fungus
Phomopsis sp. MGF222 [
202]. Compound
165 exhibited inhibitory activities against four out of five human cancer cell lines tested except A549 [
184] (
Table 7). It was also tested for the antibacterial activities against seven pathogenic bacteria and for the inhibitory activities against α-glucosidase, but no effect was observed [
202].
Another A-nor steroid
166 was isolated from the fungus of
Lasiodiplodia pseudotheobromae [
11]. A distinguished structural feature of this compound is an additional δ-lactone ring between C-3 and C-9.
Two nearly identical steroids
167 and
168 featured a bicyclo[3.3.1]nonane motif were discovered in the fungi
Phomopsis sp. TJ507A [
7] and
Stereum hirsutum [
17]. The only difference in their structures is the presence of a methoxy group in phomopsterone A (
167) instead of an ethoxy one in steresterone A (
168). Compound
167 was tested for NO inhibitory activity. Steresterone A (
168) was evaluated for the cytotoxicity against five human cancer cell lines. Both compounds showed no activity in the respective tests.
Three C25 steroids, neocyclocitrinols E-G (
169–
171) were isolated from endophytic fungus
Chaetomium sp. M453 [
189]. All compounds were tested for AChE inhibitory activities and cytotoxicity, however, no effect was found.
Cheng et al. isolated from
Ganoderma theaecolum ganotheaecolin A (
173), having a naphtho[1,8-ef]azulene ring system steroid [
291]. At a concentration of 10 μM, it showed activity to promote neurite growth in PC12 cells, comparable to that of nerve growth factor used as control.
A new steroid sarocladione (
174) bearing a 5,10:8,9-diseco moiety was isolated from the deep-sea-derived fungus
Sarocladium kiliense [
292]. The initially proposed configuration at C-3 and C-7 proved to be incorrect and was revised to 3
S,7
R through the chemical synthesis [
293]. Cytotoxic studies of compound
174 revealed no apparent cellular toxicities.
Lin et al. isolated from the sponge-derived fungus
Aspergillus flocculosus 16D-1 two 11(9→10)-
abeo-5,10-secosteroids, aspersecosteroids A (
175) and B (
176) [
278], a characteristic structural feature of which was the presence of a dioxatetraheterocyclic ring system. Both compounds were non-cytotoxic at the concentrations up to 40 μM and showed a strong inhibitory effect on the production of TNF-α and IL-6.
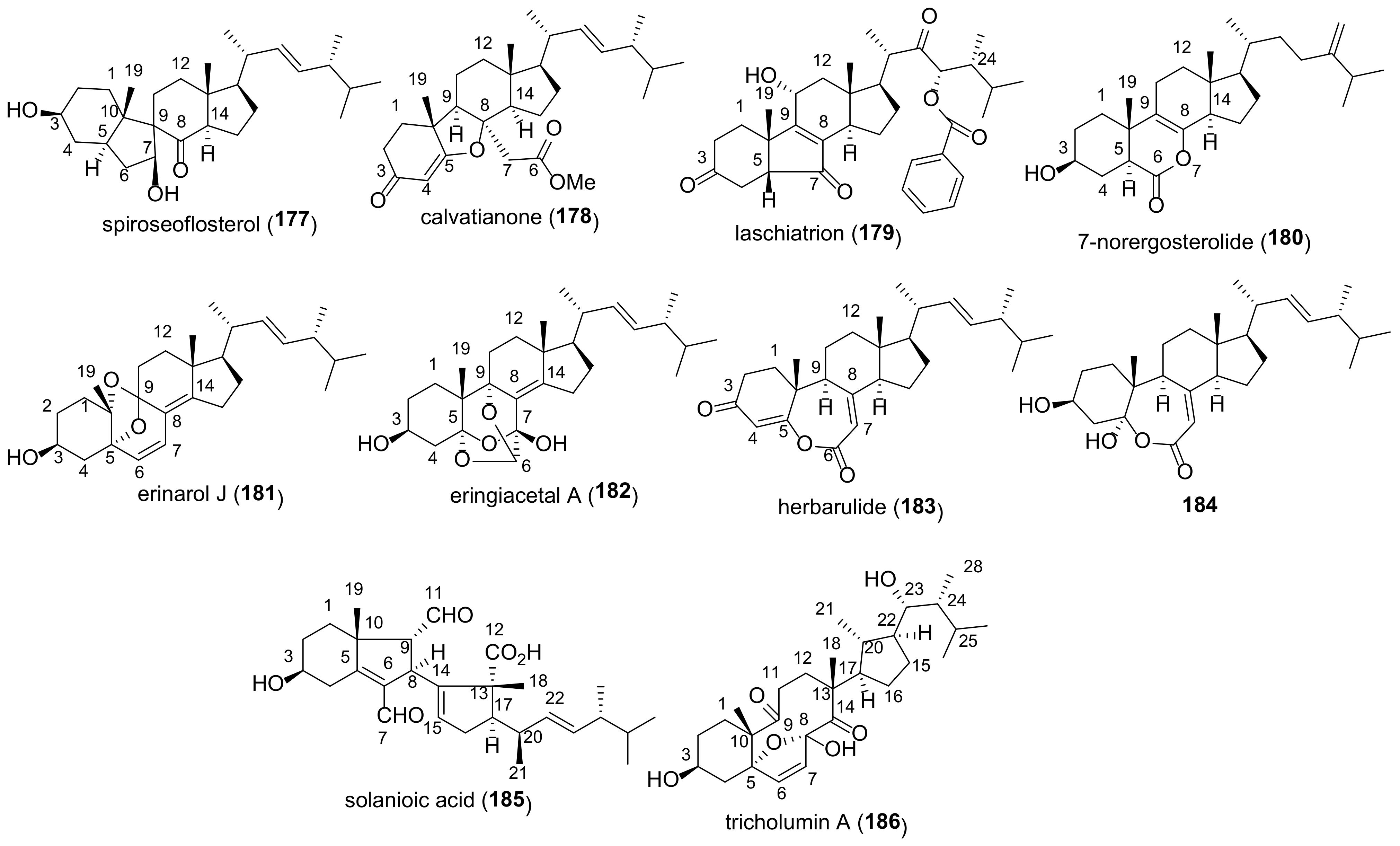
Spiroseoflosterol (
177) (
Figure 19), having a unique spiro[4.5]decan-6-one moiety, was isolated from the fruiting bodies of
Butyriboletus roseoflavus [
294]. It showed a strong cytotoxic effect on HepG2 cell line (IC
50 9.1 μM), which was comparable to that of sorafenib (IC
50 5.5 μM) used as a positive control. Moreover, spiroseoflosterol (
177) was active against sorafenib-resistant Huh7/S cells with an IC
50 value of 6.2 μM, that makes it a promising candidate for antihepatoma drug development.
Calvatianone (
178), featuring a contracted tetrahydrofuran B-ring, was found in a rare mushroom
Calvatia nipponica [
126]. It showed a weak cytotoxicity against MCF-7 with IC
50 > 100 μM (
Table 7).
Another compound with a five-membered B ring, laschiatrion (
179), was isolated from fermentations of
Favolaschia sp. [
281,
295]. It was not active in antibacterial and cytotoxic assays, but exhibited antifungal activity in the agar diffusion test [
281].
7-Nor-ergosterolide (
180), featuring a pentalactone B-ring system, was found in the culture extract of an endophytic fungus
Aspergillus ochraceus EN-31 [
296] and a halotolerant fungus
Aspergillus flocculosus PT05-1 [
13]. Compound
180 showed pronounced cytotoxic and antibacterial properties.
A characteristic structural feature of erinarol J (
181), isolated from the dried fruiting bodies of
Hericium erinaceum, is the presence of 6,8-dioxabicyclo[3.2.1]oct-2-ene moiety [
187]. Biotests have shown potent anti-inflammatory activity of
181 due to the inhibition of TNF-α secretion and NO production.
The first natural 5,6-secosteroid, eringiacetal A (
182), was isolated from the fruiting bodies of mushroom
Pleurotus eryngii [
250]. Biological assays showed its modest cytotoxicity and ability to inhibit NO production.
Herbarulide (
183) was first isolated from the endophytic fungus
Pleospora herbarum as a compound having a campestane side chain [
297]. Later the same structure was assigned to one of the constituents of the Taiwanese fungus
Antrodia camphorate [
298]. The correct structure of herbarulide (
183) was proposed by Chen and Liu who isolated it from the fungus
Stereum hirsutum [
17]. The assignment was based rather on the assumption that the C-24 stereocenter of the starting ergosterol will remain unchanged during the transformations in the cyclic part. Finally, the correct structure of
183 was confirmed by its chemical synthesis [
299]. Compound
184, structurally very close to herbarulide (
183), was isolated from the fruiting bodies of
Ganoderma resinaceum [
103].
Solanioic acid (
185) is a degraded and rearranged steroid isolated from laboratory cultures of the fungus
Rhizoctonia solani [
300]. An important feature of its biological activity is antibacterial effect against methicillin-resistant
Staphylococcus aureus. The latter is a cause of infection that is difficult to treat due to resistance to many antibiotics.
Tricholumin A (
186) was isolated from the alga-endophytic fungus
Trichoderma asperellum [
301]. The only structural element of the parent ergosterol that remained after a number of metabolic stages of its biosynthesis is cycle A. The rest of the molecule, including a fragment of the side chain, has undergone deep transformations. Inhibitory properties of
186 against harmful microalgae and weak antibacterial activity against five aquatic pathogens were found.
Dankasterone A (
187) (
Figure 20) was first isolated from a fungal strain of
Gymnascella dankaliensis derived from the sponge
Halichondria japonica [
302]. The initial erroneous assignment of stereochemistry at C-24 was corrected from
S to
R in a follow-up work by these authors [
303]. Subsequently, compound
187 was repeatedly isolated from fungal sources as one of the ergostane constituents (
Table 7). The only structural difference between
187 and dankasterone B (
188) is the saturated ring A. From the endophytic fungus
Phomopsis sp. TJ507A was also isolated phomopsterone B (
190) differing from
187 by the presence of a methyl group at C-23 [
7]. Dankasterone A (
187) showed promising anticancer activities with IC
50 down to 2.3 μM on a range of cancer cell lines (
Table 7). Structure activity relationship studies of dankasterones A and B showed that the Δ
4-double bond is essential for high cytotoxicity against the cancer cell lines tested. Carbonyl groups in dankasterone B (
188) were other structural elements important for the high biological activity, because products of its NaBH
4 reduction were not cytotoxic [
17]. Phomopsterone B (
190) was tested for inflammatory activity and showed promising results in iNOS inhibitory and NO production inhibition assays [
7].
At first glance, the carbon skeleton of periconiastone A (
189) [
304] looks completely different from that of dankasterone B (
188). In fact, compound
189 is available from
188 in one step via the intramolecular aldol reaction [
305], which is also evidently realized in the course of its biosynthesis. So far, periconiastone A (
189) has been tested for anti-inflammatory and antibacterial activities. Positive results were obtained in an antibacterial assay against Gram-positive bacteria [
304].
An 8,14-seco-steroid, childinasterone A (
191), was isolated from fruiting bodies of the ascomycete
Daldinia childiae [
306]. It showed no activity in cytotoxic studies and exhibited strong inhibition of NO production (IC
50 value of 21.2 μM versus 41.5 μM for L-NMMA used as a positive control).
9,11-Secosteroids are quite common in sea sponges [
22], but rather rare in fungal sources. The first such an ergostane
192 was isolated from king trumpet mushroom
Pleurotus eryngii [
6]. Compound
192 exhibited NO inhibitory activity similar to that of L-NMMA and revealed no cytotoxicity. Another 9,11-secoergostane (
193), found in the fruiting bodies of
Pleurotus eryngii, displayed similar profile of biological activity [
6].
Three steroids with a rearranged ring B, eringiacetal B (
194), matsutakone (
195), and pleurocin B (
196), were isolated from the fruiting bodies of
Pleurotus eryngii by Tanaka et al. [
248]. All three compounds revealed inhibitory activity on production of NO which was stronger than that of L-NMMA. The 13,14-seco-13,14-epoxysteroid, eringiacetal B (
194), was most active with an IC
50 of 13.0 μM compared to 23.9 μM for the L-NMMA positive control.
An 8(14→15)-abeo-steroid, asperflotone (
197), was obtained from the solid culture of
Aspergillus flocculosus 16D-1 [
277]. Its characteristic structural feature is a rearranged bicyclo[4.2.1]non-2-ene ring system. Compound
197 was tested on three cancer cell lines with no cytotoxic effects. In immune-suppressive activity assay, asperflotone (
197) exhibited inhibitory effects on IL-6 secretion.
The 15(14→22)abeo-steroid framework is common for ergostanes
198–
203 (
Figure 21), collectively referred to as strophasterols. It took some effort to establish the correct structures of these structurally related compounds. Strophasterols A–D (
198–
201) were first isolated from the mushroom
Stropharia rugosoannulata [
307]. The structure of strophasterin A (
198) was established by X-ray crystallographic analysis. Comparison of the NMR data made it possible to assign the structure of
199 as the C-22 isomer of strophasterol A that was later confirmed by X-ray analysis [
193]. Structure of strophasterol C (
200) was proposed based on NOE correlations by Aung et al., who isolated it from the basidiomycete
Cortinarius glaucopus together with glaucoposterol A (
203) [
195]. Additional evidence for the structure of
200 was obtained by its chemical synthesis [
308]. Two more steroids with a strophastane skeleton, strophasterol E (
202) and strophasterol F (
203), were isolated from the fruiting bodies of
Pleurotus eryngii [
201]. Their structures were determined by X-ray analysis of the corresponding tris-p-bromobenzoate derivatives. Structural elucidation of strophasterol D (
201) was done by comparing it with a synthetically prepared sample [
309]. This work also showed that glaucoposterol A and strophasterol F are the same compound (
203).
So far, the biological activity of strophasterols has been studied only marginally. Strophasterol A (
198) showed a dose-dependent inhibitory effect on the toxicity of thapsigargin. The latter is known to disrupt the balance of the Ca
2+ concentration in the endoplasmic reticulum that is especially harmful to neuronal cells. Under the action of strophasterol A (
198), an increase in cell viability by 10.3% compared with the control was noted [
307]. Strophasterols E and F were tested for anti-inflammatory activity, but showed no promising results [
201].
A 15(14→11)-abeo-ergostane, penicillitone (
204), was isolated from the culture of the fungus
Penicillium purpurogenum SC0070 [
254]. It was evaluated for cytotoxicity against three cancer lines and showed good potency with IC
50 ranging from 4.44 to 5.98 μM. In addition, compound
204 was active in the inflammatory assay on the production of TNF-α and IL-6. At the concentration of 5 μM it reduced their secretion by 70.7% and 96.6%, respectively. For comparison, inhibition rates of the positive control dexamethasone at 100 μM were 87.3% and 96.7%, respectively. This makes promising further in-depth study of penicillitone (
204) as an anti-inflammatory or antitumor agent.
Table 7.
Sources and biological activity of fungal steroids with a rearranged tetracyclic carbon skeleton.
Table 7.
Sources and biological activity of fungal steroids with a rearranged tetracyclic carbon skeleton.
| Compound | Fungal Source [Ref.] | Assays (Activity) [Ref.] |
|---|
| 157 | Antrodia camphorata [310], Aspergillus ustus [231], Gibberella zeae [311] | cytotoxic assay (CT26, IC50 15.3 μM; K562, IC50 19.9 μM) [310] |
| 158 | Antrodia camphorata [310], Penicillium citreo-viride [312], Phyllosticta capitalensis [288] | cytotoxic assay (CT26, IC50 18.2 μM; K562, IC50 12.5 μM) [310], neuroprotective activity assay (EC50 24.2 μM) [288] |
| 159a | Aspergillus ustus [231] | |
| 159b | Aspergillus ustus [231] | |
| 160 | Penicillium citreo-viride [312] | |
| 161 | Aspergillus ustus [280] | antimicrobial assay (Candida albicans, MIC50 17.24 μg/mL; Escherichia coli, MIC50 17.24 μg/mL; Staphylococcus aureus, MIC50 15.51 μg/mL) [280], cytotoxic assay (A549, IC50 40.32 μM; Hela, IC50 26.09 μM; HT-29, IC50 43.58 μM; MCF-7, IC50 32.03 μM) [280], immunosuppressive assay (ConA-induced T-cell proliferation, IC50 23.61 μM; LPS-induced B-cell proliferation, IC50 23.61 μM) [280] |
| 162a | Malbranchea filamentosa [289] | cytotoxic assay (A549, IC50 38.6 μM; Hela, IC50 28.1 μM) [289] |
| 162b | Malbranchea filamentosa [289] | cytotoxic assay (A549, Hela, no activity) [289] |
| 162c | Malbranchea filamentosa [289] | cytotoxic assay (Hela, IC50 76.9 μM) [289] |
| 163 | Aspergillus flocculosus [290] | anti-inflammatory assay [290] |
| 164 | Aspergillus flocculosus [290] | anti-inflammatory assay [290] |
| 165 | Phomopsis sp. [202], Polyporus ellisii [184] | α-glucosidase inhibition assay (IC50 > 100 μM) [202], cytotoxic assay (A549, IC50 > 40 μM; HL-60, IC50 17.1 μM; MCF-7, IC50 23.3 μM; SMMC-7721, IC50 21.3 μM; SW480, IC50 16.3 μM) [184] |
| 166 | Lasiodiplodia pseudotheobromae [11] | |
| 167 | Phomopsis sp. [7] | NO production inhibition assay (IC50 > 25 μM) [7] |
| 168 | Stereum hirsutum [17] | cytotoxic assay (A549, HL-60, MCF-7, SMMC-7721, SW480, IC50 > 40 μM) [17] |
| 169 | Chaetomium sp. [189] | cytotoxic assay (A549, HL-60, MCF-7, SMMC-7721, SW480, IC50 > 40 μM) [189] |
| 170 | Chaetomium sp. [189] | cytotoxic assay (A549, HL-60, MCF-7, SMMC-7721, SW480, IC50 > 40 μM) [189] |
| 171 | Chaetomium sp. [189] | cytotoxic assay (A549, HL-60, MCF-7, SMMC-7721, SW480, IC50 > 40 μM) [189] |
| 172 | Xylaria sp. [313] | |
| 173 | Ganoderma theaecolum [291] | neurite outgrowth-promoting assay in PC12 cells (stimulated cell differentiation with a maximum effect at 10 μM) [291] |
| 174 | Sarocladium kiliense [292] | cytotoxic assay (Bel-7402, ECA-109, HeLa, PANC-1, SHG-44, no activity) [292] |
| 175 | Aspergillus flocculosus [278] | cytotoxic assay (A549, HepG2, THP-1, IC50 > 80 μM) [278], IL-6 immune-suppressive activity assay (IC50 21 μM), TNF-α secretion assay (IC50 28 μM) [278] |
| 176 | Aspergillus flocculosus [278] | cytotoxic assay (A549, HepG2, THP-1, IC50 > 80 μM) [278], IL-6 immune-suppressive activity assay (IC50 26 μM), TNF-α secretion assay (IC50 31 μM) [278] |
| 177 | Butyriboletus roseoflavus [294] | cytotoxic assay (HepG2, IC50 9.1 μM; Huh7/S, IC50 6.2 μM; L02, IC50 22.8 μM) [294] |
| 178 | Calvatia nipponica [126] | cytotoxic assay (MCF-7, IC50 > 100 μM) [126] |
| 179 | Favolaschia calocera [281], Favolaschia sp. [295] | antifungal assay (activity against Candida albicans, Cryptococcus neoformans, etc. at concentrations of 10–50 μg/mL) [295] |
| 180 | Aspergillus flocculosus [13], Aspergillus ochraceus [296] | antibacterial assay (MIC 1.9 μg/mL against Candida albicans, 7.5 μg/mL against Pseudomonas aeruginosa and Enterobacter aerogenes) [13], cytotoxic assay (BEL-7402, IC50 17.7 μM; HL-60, IC50 12.4 μM) [13], (NCI-H460, IC50 5.0 μg/mL; SMMC-7721, IC50 7.0 μg/mL; SW1990, IC50 28.0 μg/mL) [296] |
| 181 | Hericium erinaceum [187] | NO production inhibition assay (38.4% inhibition at 10 μg/mL) [187], TNF-α secretion assay (43.3% inhibition at 10 μg/mL) [187] |
| 182 | Pleurotus eryngii [250] | cytotoxic assay (RAW264.7, IC50 25.6 μM) [250], NO production inhibition assay (IC50 19.9 μM) [250] |
| 183 | Antrodia camphorate [298], Gymnoascus reessii [249], Stereum hirsutum [17] | cytotoxic assay (A549, HL-60, MCF-7, SMMC-7721, SW480, IC50 > 40 μM) [17], (KB, MCF-7, IC50 > 50 μM; NCI-H187, IC50 22.6 μM; Vero, IC50 43.8 μM) [249] |
| 184 | Ganoderma resinaceum [103] | NO production inhibition assay (56.37% inhibition at 50 μM) [103] |
| 185 | Rhizoctonia solani [300] | antibacterial assay (MIC 1 μg/mL against the Gram-positive bacteria Bacillus subtilis, Staphylococcus aureus, and MRSA; MIC 16 μg/mL against the yeast Candida albicans; MIC 64 μg/mL against the Gram-negative bacteria Escherichia coli and Pseudomonas aeruginosa) [300] |
| 186 | Trichoderma asperellum [301] | antibacterial assay (against V. harveyi, V. splendidus, and P. citrea with inhibitory zones of 10, 7.5, and 8.0 mm, respectively, at 50 μg/disk) [301], antifungal assay (MIC 12 μg/mL against Glomerella cingulate) [301] |
| 187 | Antrodia camphorate [310], Arthrinium sp. [314], Aspergillus penicillioides [205], Colletotrichum sp. [206], Conocybe siliginea [315], Gymnascella dankaliensis [303], Neosartorya fennelliae, N. tsunodae [316], Pestalotiopsis sp. [139], Phomopsis sp. [7], Pleosporales sp. [317], Stereum hirsutum [17], Talaromyces purpurogenus [318], Talaromyces sp. [255] | cytotoxic assay (P388, ED50 2.2 μg/mL) [303], (A549, IC50 4.4 μM; HL-60, IC50 2.3 μM; MCF-7, IC50 2.7 μM; SMMC-7721, IC50 3.3 μM; SW480, IC50 3.5 μM) [17], (K562, IC50 > 20 μM; ST26, IC50 6.7 μM) [310], (A549, IC50 21.3 μM; HL-60, IC50 7.9 μM; MCF-7, IC50 23.8 μM; SMMC-7721, IC50 > 40 μM; SW480, IC50 14.2 μM) [318], iNOS inhibitory assay (IC50 6.58 μM) [7], NO production inhibition assay (IC50 13.04 μM) [7] |
| 188 | Antrodia camphorate [310], Calvatia nipponica [126], Gymnascella dankaliensis [303], Stereum hirsutum [17] | cytotoxic assay (P388, ED50 2.8 μg/mL) [303], (MCF-7, IC50 > 100 μM) [126], (A549, IC50 16.6 μM; HL-60, IC50 15.6 μM; MCF-7, IC50 17.2 μM; SMMC-7721, IC50 16.3 μM; SW480, IC50 17.3 μM) [17], (K562, IC50 23.1 μM; ST26, IC50 8.4 μM) [310] |
| 189 | Periconia sp. [304] | antibacterial assay (MIC 4 μg/mL against Staphylococcus aureus, MIC 32 μg/mL against Enterococcus faecalis; MIC > 100 μg/mL against all four Gram-negative bacteria tested) [304], NO production inhibition assay (IC50 > 40 μM) [304] |
| 190 | Phomopsis sp. [7] | iNOS inhibitory assay (IC50 1.49 μM) [7], NO production inhibition assay (IC50 4.65 μM) [7] |
| 191 | Daldinia childiae [306] | cytotoxic assay (MCF-7, SMMC-7721, SW480, IC50 > 40 μM) [306], NO production inhibition assay (IC50 21.2 μM) [306] |
| 192 | Pleurotus eryngii [6] | NO production inhibition assay (IC50 10.3 μM) [6] |
| 193 | Pleurotus eryngii [201] | NO production inhibition assay (NO produced 57.8% at 30 μM) [201] |
| 194 | Pleurotus eryngii [248] | NO production inhibition assay (IC50 13.0 μM) [248] |
| 195 | Tricholoma matsutake [319], Pleurotus eryngii [248] | AChE inhibitory assay (62.8% inhibition at 50 μg/mL) [319], NO production inhibition assay (IC50 25 μM) [248] |
| 196 | Pleurotus eryngii [248] | NO production inhibition assay (IC50 23.6 μM) [248] |
| 197 | Aspergillus flocculosus [277] | cytotoxic assay (A549, HepG2, THP-1, IC50 > 80 μM) [277], IL-6 immune-suppressive activity assay (IC50 22 μM) [277] |
| 198 | Stropharia rugosoannulata [307] | |
| 199 | Stropharia rugosoannulata [307] | |
| 200 | Stropharia rugosoannulata [307] | |
| 201 | Cortinarius glaucopus [195], Stropharia rugosoannulata [307] | |
| 202 | Pleurotus eryngii [201] | cytotoxic assay (RAW 264.7, IC50 > 30 μM) [201] |
| 203 | Pleurotus eryngii [201] | cytotoxic assay (RAW 264.7, IC50 > 30 μM) [201] |
| 204 | Penicillium purpurogenum [254] | cytotoxic assay (A549, IC50 5.57 μM; HepG2, IC50 4.44 μM; MCF-7, IC50 5.98 μM) [254], IL-6 immune-suppressive activity assay (96.7% inhibition at 5 μg/mL) [254], NO production inhibition assay (70.7% inhibition at 5 μg/mL) [254] |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}