
Characterisation of a Novel Acetyl Xylan Esterase (BaAXE) Screened from the Gut Microbiota of the Common Black Slug (Arion ater)

Abstract
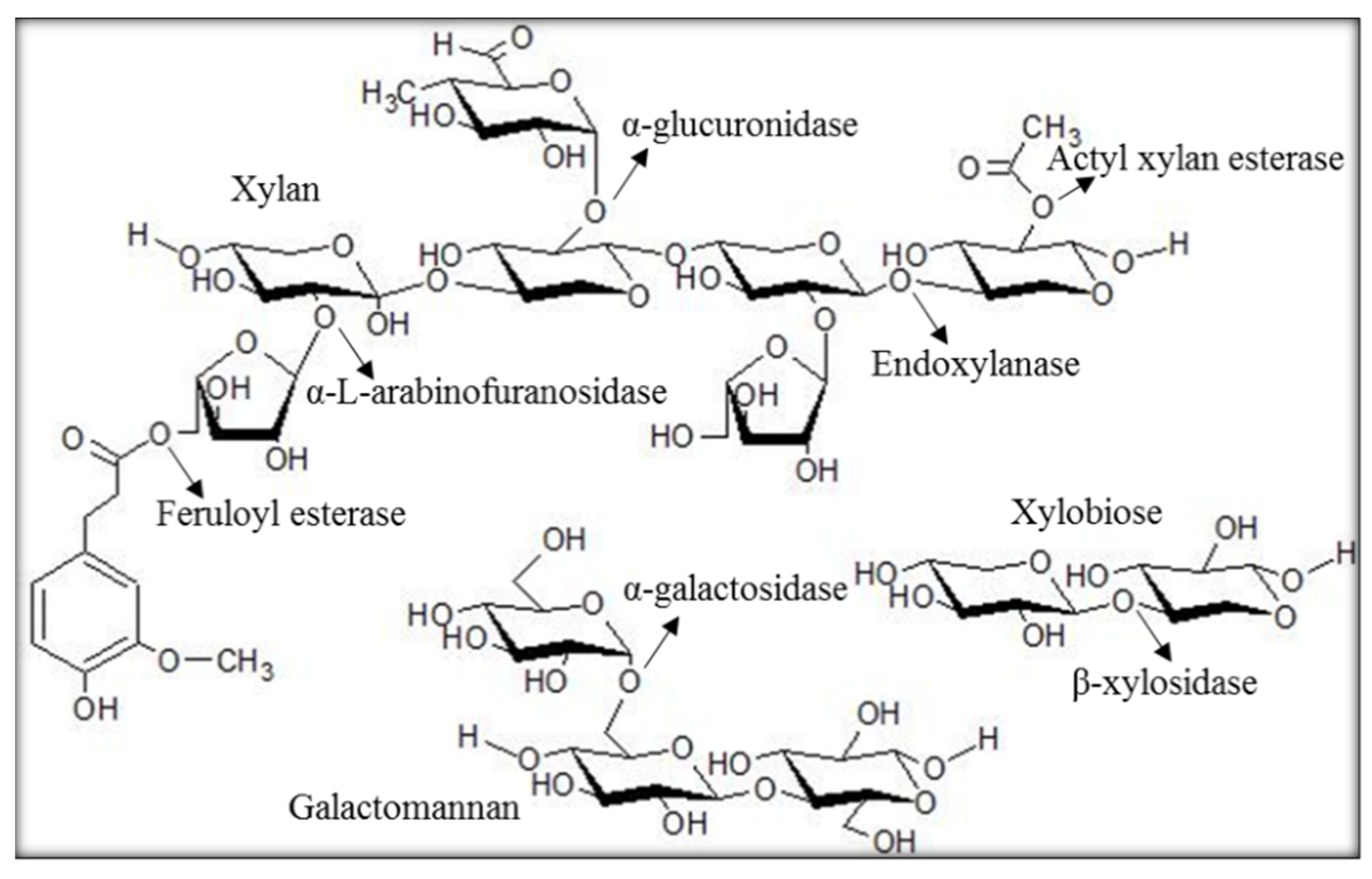
:1. Introduction
2. Results
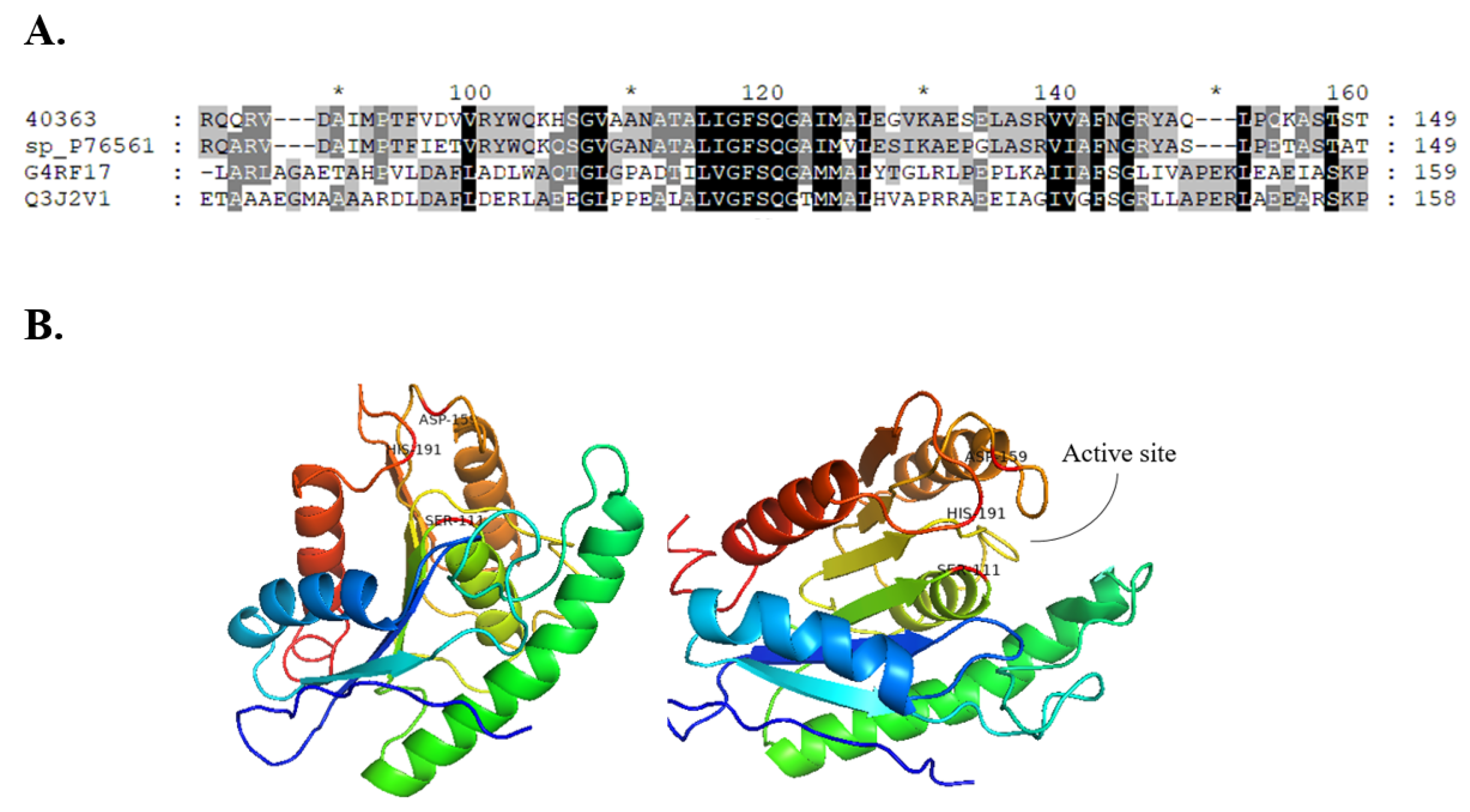
2.1. Bioinformatics and Homology Modelling
2.2. Recombinant Protein and Activity Assay
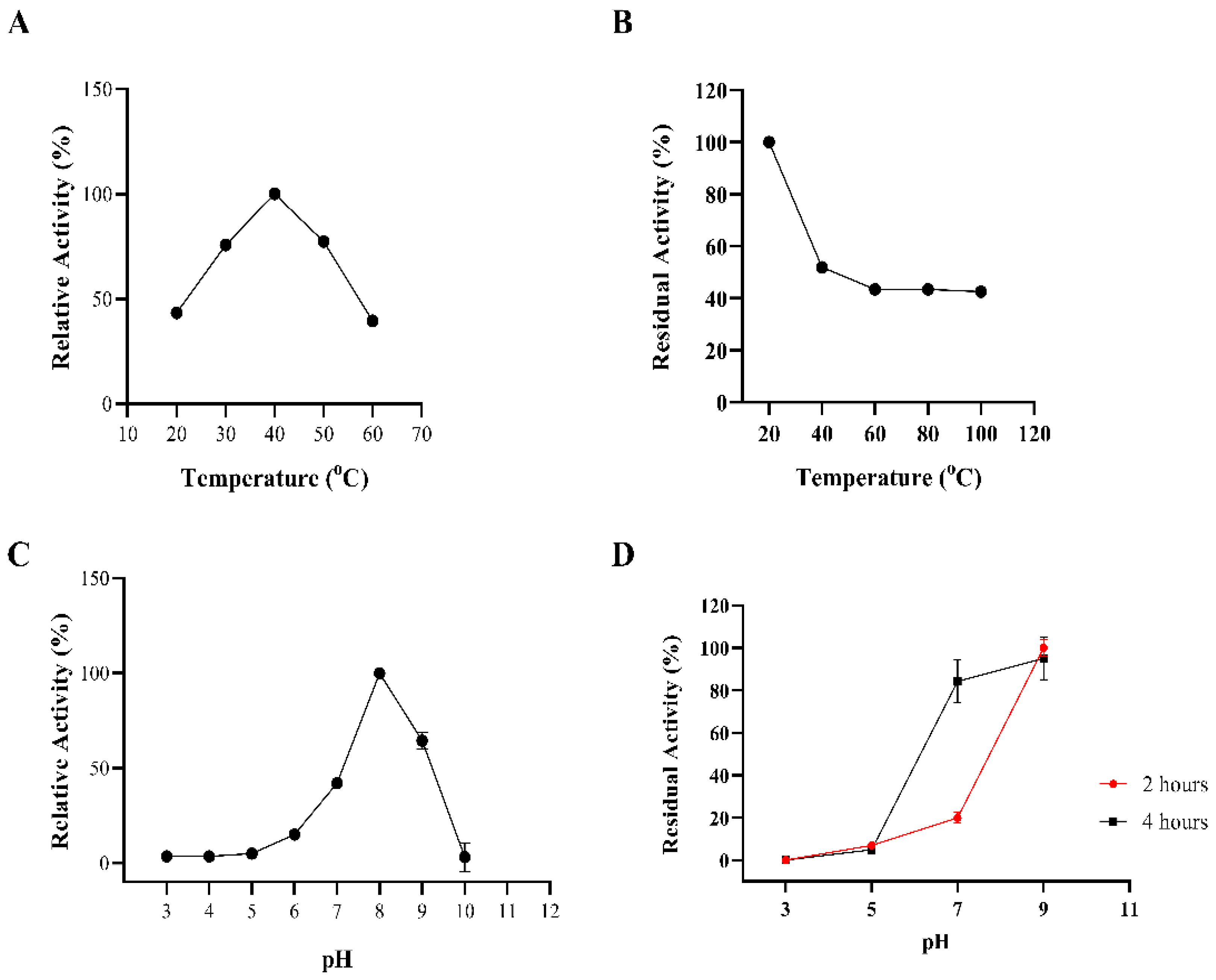
2.3. Biochemical Properties
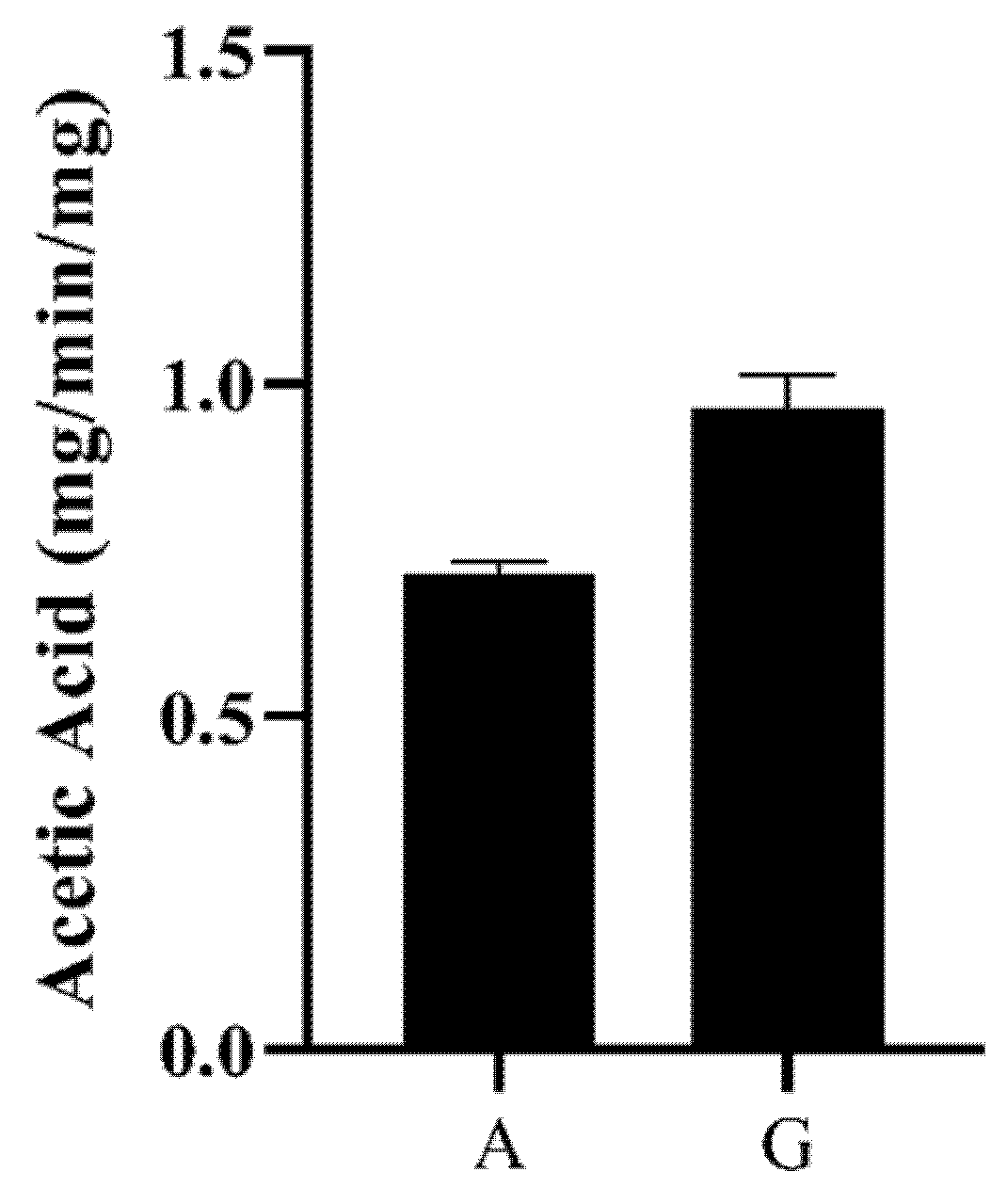
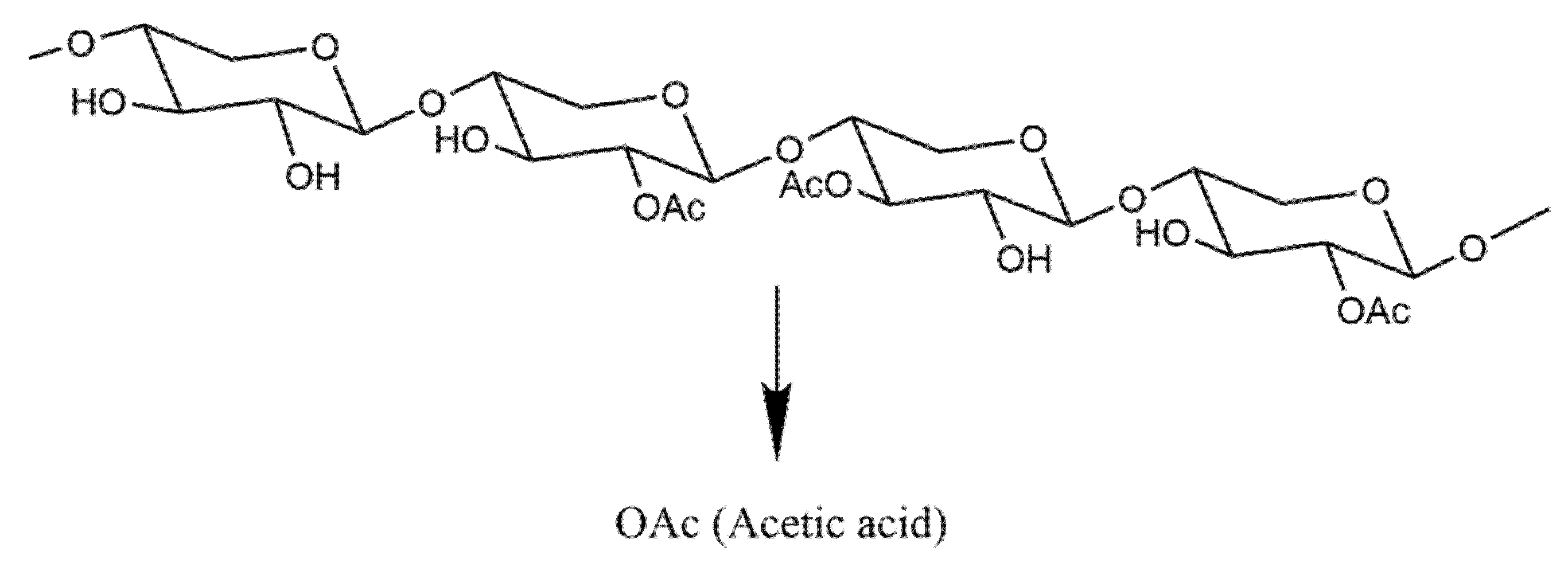
2.4. Functional Characterisation
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Designation | Source Organism | Temp. Opt. | pH Opt. | Temp. Stability | Reference |
|---|---|---|---|---|---|
| axeA | Aspergillus awamori | - | 7 | Enzyme activity decreased at temperatures higher than 40 °C. | [53] |
| axeA | Aspergillus ficuum | 40 °C | 7 | Thermal stability decreased at temperatures above 40 °C. | [54] |
| rAoAXE | Aspergillus oryzae | 45 °C | 6 | Unstable at 40 degrees Celsius, with a half-life of less than 60 min at 40 °C and 10 min at 50 °C | [55] |
| AXE | Bacillus pumilus PS213 | 55 °C | 8 | Stable at 50 °C but rapidly inactivated at a temperature higher than 60 °C, with a half-life of about 1 h at this temperature | [56] |
| TM0077 | Thermotoga maritima | 100 °C | 7.5 | Unstable at 100 °C with a half-life of <5 min. | [57] |
| AxeA | Thermotoga maritima | 90 °C | 6.5 | Retained over 60% residual activity after 8 h incubation at 98 °C | [58] |
| AXE | Ochrovirga pacifica | 50 °C | 8 | Maintained over 70% residual activity after incubation at 55 °C for 120 min | [25] |
| rAoAXEC | Aspergillus oryzae | 50 °C | 7 | Stable up to 50 °C, with a half-life of approximately 2 h at 50 °C and 40 min at 60 °C | [28] |
| LaAXE | Lactobacillus antri | 50 °C | 7 | At 70 °C, the residual activity decreased to 48.4% | [40] |
| AXE1 | Thermoanaerobacterium sp. | 80 °C | 7 | At 75 °C, the enzyme showed a half-life of 1 h. | [59] |
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Bioinformatics
4.2.2. Amplification, Cloning, and Bacterial Transformation
4.2.3. Protein Expression and Purification
4.2.4. Enzyme Assay and Biochemical Properties
4.2.5. Biochemical Properties
4.2.6. Thermostability
4.2.7. Effect of Organic Solvents and Additives
4.2.8. Acyl Chain Substrate
4.2.9. 4-Methylumberiferyl Acetate (4-MUA)
4.2.10. Hydrocinnamate Substrates
4.2.11. Acetic Acid Release from Acetylated Substrates
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Kumar, R.; Strezov, V.; Weldekidan, H.; He, J.; Singh, S.; Kan, T.; Dastjerdi, B. Lignocellulose Biomass Pyrolysis for Bio-Oil Production: A Review of Biomass Pre-Treatment Methods for Production of Drop-in Fuels. Renew. Sustain. Energy Rev. 2020, 123, 109763. [Google Scholar] [CrossRef]
- Aghbashlo, M.; Khounani, Z.; Hosseinzadeh-Bandbafha, H.; Gupta, V.K.; Amiri, H.; Lam, S.S.; Morosuk, T.; Tabatabaei, M. Exergoenvironmental Analysis of Bioenergy Systems: A Comprehensive Review. Renew. Sustain. Energy Rev. 2021, 149, 111399. [Google Scholar] [CrossRef]
- Rittmann, B.E. Opportunities for Renewable Bioenergy Using Microorganisms. Biotechnol. Bioeng. 2008, 100, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Lorenci Woiciechowski, A.; Dalmas Neto, C.J.; Porto de Souza Vandenberghe, L.; de Carvalho Neto, D.P.; Novak Sydney, A.C.; Letti, L.A.J.; Karp, S.G.; Zevallos Torres, L.A.; Soccol, C.R. Lignocellulosic Biomass: Acid and Alkaline Pretreatments and Their Effects on Biomass Recalcitrance—Conventional Processing and Recent Advances. Bioresour. Technol. 2020, 304, 122848. [Google Scholar] [CrossRef]
- Abraham, A.; Mathew, A.K.; Park, H.; Choi, O.; Sindhu, R.; Parameswaran, B.; Pandey, A.; Park, J.H.; Sang, B.I. Pretreatment Strategies for Enhanced Biogas Production from Lignocellulosic Biomass. Bioresour. Technol. 2020, 301, 122725. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Pandey, A. Biological Pretreatment of Lignocellulosic Biomass—An Overview. Bioresour. Technol. 2016, 199, 76–82. [Google Scholar] [CrossRef]
- Hazeena, S.H.; Sindhu, R.; Pandey, A.; Binod, P. Lignocellulosic Bio-Refinery Approach for Microbial 2,3-Butanediol Production. Bioresour. Technol. 2020, 302, 122873. [Google Scholar] [CrossRef]
- Kumar, B.; Bhardwaj, N.; Agrawal, K.; Chaturvedi, V.; Verma, P. Current Perspective on Pretreatment Technologies Using Lignocellulosic Biomass: An Emerging Biorefinery Concept. Fuel Process. Technol. 2020, 199, 106244. [Google Scholar] [CrossRef]
- Rebello, S.; Anoopkumar, A.N.; Aneesh, E.M.; Sindhu, R.; Binod, P.; Pandey, A. Sustainability and Life Cycle Assessments of Lignocellulosic and Algal Pretreatments. Bioresour. Technol. 2020, 301, 122678. [Google Scholar] [CrossRef]
- Raghavi, S.; Sindhu, R.; Binod, P.; Gnansounou, E.; Pandey, A. Development of a Novel Sequential Pretreatment Strategy for the Production of Bioethanol from Sugarcane Trash. Bioresour. Technol. 2016, 199, 202–210. [Google Scholar] [CrossRef]
- Sindhu, R.; Kuttiraja, M.; Prabisha, T.P.; Binod, P.; Sukumaran, R.K.; Pandey, A. Development of a Combined Pretreatment and Hydrolysis Strategy of Rice Straw for the Production of Bioethanol and Biopolymer. Bioresour. Technol. 2016, 215, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, X.; Yao, Q.; Zong, W.; Dai, S.; Deng, Z.; Liu, S.; Yun, J.; Yang, X.; Li, H. Cloning, Characterization of a Novel Acetyl Xylan Esterase, and Its Potential Application on Wheat Straw Utilization. All Life 2021, 14, 622–635. [Google Scholar] [CrossRef]
- Farhat, W.; Venditti, R.; Ayoub, A.; Prochazka, F.; Fernández-de-Alba, C.; Mignard, N.; Taha, M.; Becquart, F. Towards Thermoplastic Hemicellulose: Chemistry and Characteristics of Poly-(ε-Caprolactone) Grafting onto Hemicellulose Backbones. Mater. Des. 2018, 153, 298–307. [Google Scholar] [CrossRef]
- Huang, L.Z.; Ma, M.G.; Ji, X.X.; Choi, S.E.; Si, C. Recent Developments and Applications of Hemicellulose from Wheat Straw: A Review. Front. Bioeng. Biotechnol. 2021, 9, 690773. [Google Scholar] [CrossRef]
- Ahmad, N.; Tayyeb, D.; Ali, I.; Alruwaili, N.K.; Ahmad, W.; ur Rehman, A.; Khan, A.H.; Iqbal, M.S. Development and Characterization of Hemicellulose-Based Films for Antibacterial Wound-Dressing Application. Polymers 2020, 12, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hameleers, L.; Penttinen, L.; Ikonen, M.; Jaillot, L.; Fauré, R.; Terrapon, N.; Deuss, P.J.; Hakulinen, N.; Master, E.R.; Jurak, E. Polysaccharide Utilization Loci-Driven Enzyme Discovery Reveals BD-FAE: A Bifunctional Feruloyl and Acetyl Xylan Esterase Active on Complex Natural Xylans. Biotechnol. Biofuels 2021, 14, 127. [Google Scholar] [CrossRef]
- Marasinghe, S.D.; Jo, E.; Hettiarachchi, S.A.; Lee, Y.; Eom, T.Y.; Gang, Y.; Kang, Y.H.; Oh, C. Characterization of Glycoside Hydrolase Family 11 Xylanase from Streptomyces Sp. Strain J103; Its Synergetic Effect with Acetyl Xylan Esterase and Enhancement of Enzymatic Hydrolysis of Lignocellulosic Biomass. Microb. Cell Factories 2021, 20, 129. [Google Scholar] [CrossRef]
- Binod, P.; Gnansounou, E.; Sindhu, R.; Pandey, A. Enzymes for Second Generation Biofuels: Recent Developments and Future Perspectives. Bioresour. Technol. Rep. 2019, 5, 317–325. [Google Scholar] [CrossRef]
- Shen, D.K.; Gu, S.; Bridgwater, A.V. The Thermal Performance of the Polysaccharides Extracted from Hardwood: Cellulose and Hemicellulose. Carbohydr. Polym. 2010, 82, 39–45. [Google Scholar] [CrossRef]
- Park, S.H.; Yoo, W.; Lee, C.W.; Jeong, C.S.; Shin, S.C.; Kim, H.W.; Park, H.; Kim, K.K.; Kim, T.D.; Lee, J.H. Crystal Structure and Functional Characterization of a Cold-Active Acetyl Xylan Esterase (PbAcE) from Psychrophilic Soil Microbe Paenibacillus sp. PLoS ONE 2018, 13, e0206260. [Google Scholar] [CrossRef]
- Joshi, N.; Sharma, M.; Singh, S.P. Characterization of a Novel Xylanase from an Extreme Temperature Hot Spring Metagenome for Xylooligosaccharide Production. Appl. Microbiol. Biotechnol. 2020, 104, 4889–4901. [Google Scholar] [CrossRef] [PubMed]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from Enzymatic Degradation of Cellulose and Hemicellulose to Fermentable Sugars—A Review. Biomass Bioenergy 2020, 134, 105481. [Google Scholar] [CrossRef]
- Ohta, K.; Fujii, S.; Higashida, C. Characterization of a Glycoside Hydrolase Family-51 α-l-Arabinofuranosidase Gene from Aureobasidium Pullulans ATCC 20524 and Its Encoded Product. J. Biosci. Bioeng. 2013, 116, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Han, X.; Wang, Y.; Wei, X.; Liu, S.; Shao, S.; Yang, S.; Sun, L.; Xin, F. Rational Design for Broadened Substrate Specificity and Enhanced Activity of a Novel Acetyl Xylan Esterase from Bacteroides Thetaiotaomicron. J. Agric. Food Chem. 2021, 69, 6665–6675. [Google Scholar] [CrossRef]
- Hettiarachchi, S.A.; Kwon, Y.K.; Lee, Y.; Jo, E.; Eom, T.Y.; Kang, Y.H.; Kang, D.H.; de Zoysa, M.; Marasinghe, S.D.; Oh, C. Characterization of an Acetyl Xylan Esterase from the Marine Bacterium Ochrovirga Pacifica and Its Synergism with Xylanase on Beechwood Xylan. Microb. Cell Fact. 2019, 18, 122. [Google Scholar] [CrossRef] [Green Version]
- Puchart, V.; Gjermansen, M.; Mastihubová, M.; Mørkeberg Krogh, K.B.R.; Biely, P. Positional Specificity of Flavobacterium Johnsoniae Acetylxylan Esterase and Acetyl Group Migration on Xylan Main Chain. Carbohydr. Polym. 2020, 232, 115783. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, H.; Yu, X.; Kong, H.; Chen, J.; Luo, H.; Bai, Y.; Yao, B. Synergistic Effect of Acetyl Xylan Esterase from Talaromyces Leycettanus JCM12802 and Xylanase from Neocallimastix Patriciarum Achieved by Introducing Carbohydrate-Binding Module-1. AMB Express 2019, 9, 13. [Google Scholar] [CrossRef]
- Kato, T.; Shiono, Y.; Koseki, T. Identification and Characterization of an Acetyl Xylan Esterase from Aspergillus Oryzae. J. Biosci. Bioeng. 2021, 132, 337–342. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, H.-T.; Jiang, W.-X.; Zhang, X.; Cao, H.-Y.; Wang, J.-P.; Li, C.-Y.; Huang, F.; Zhang, X.-Y.; Chen, X.-L. Active Site Architecture of an Acetyl Xylan Esterase Indicates a Novel Cold Adaptation Strategy. J. Biol. Chem. 2021, 297, 100841. [Google Scholar] [CrossRef]
- Sista Kameshwar, A.K.; Qin, W. Understanding the Structural and Functional Properties of Carbohydrate Esterases with a Special Focus on Hemicellulose Deacetylating Acetyl Xylan Esterases. Mycology 2018, 9, 273–295. [Google Scholar] [CrossRef] [Green Version]
- Burlacu, A.; Israel-Roming, F.; Cornea, C.P. Screening of microorganisms displaying acetyl xylan esterase activity. Sci. Pap. Ser. B Hortic. 2018, 62, 715–720. [Google Scholar]
- Rashamuse, K.; Ronneburg, T.; Sanyika, W.; Mathiba, K.; Mmutlane, E.; Brady, D. Metagenomic Mining of Feruloyl Esterases from Termite Enteric Flora. Appl. Microbiol. Biotechnol. 2014, 98, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Golyshina, O.V.; Chernikova, T.N.; Khachane, A.N.; Reyes-Duarte, D.; Martins Dos Santos, V.A.P.; Strompl, C.; Elborough, K.; Jarvis, G.; Neef, A.; et al. Novel Hydrolase Diversity Retrieved from a Metagenome Library of Bovine Rumen Microflora. Environ. Microbiol. 2005, 7, 1996–2010. [Google Scholar] [CrossRef]
- Adesioye, F.A.; Makhalanyane, T.P.; Biely, P.; Cowan, D.A. Phylogeny, Classification and Metagenomic Bioprospecting of Microbial Acetyl Xylan Esterases. Enzym. Microb. Technol. 2016, 93–94, 79–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razeq, F.M.; Jurak, E.; Stogios, P.J.; Yan, R.; Tenkanen, M.; Kabel, M.A.; Wang, W.; Master, E.R. A Novel Acetyl Xylan Esterase Enabling Complete Deacetylation of Substituted Xylans. Biotechnol. Biofuels 2018, 11, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peláez, M.L.; Valdecasas, A.G.; Martinez, D.; Horreo, J.L. Towards the Unravelling of the Slug A. Ater-A. Rufus Complex (Gastropoda Arionidae): New Genetic Approaches. Web Ecol. 2018, 18, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Joynson, R.; Pritchard, L.; Osemwekha, E.; Ferry, N. Metagenomic Analysis of the Gut Microbiome of the Common Black Slug Arion Ater in Search of Novel Lignocellulose Degrading Enzymes. Front. Microbiol. 2017, 8, 2181. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Pawar, P.M.A.; Derba-Maceluch, M.; Hedenström, M.; Chong, S.L.; Tenkanen, M.; Jönsson, L.J.; Mellerowicz, E.J. Hybrid Aspen Expressing a Carbohydrate Esterase Family 5 Acetyl Xylan Esterase Under Control of a Wood-Specific Promoter Shows Improved Saccharification. Front. Plant Sci. 2020, 11, 380. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, J.; Liu, J.; Wang, R.; Liu, L.; Wang, F.; Yuan, H. The Composition of Accessory Enzymes of Penicillium Chrysogenum P33 Revealed by Secretome and Synergistic Effects with Commercial Cellulase on Lignocellulose Hydrolysis. Bioresour. Technol. 2018, 257, 54–61. [Google Scholar] [CrossRef]
- Kim, M.J.; Jang, M.U.; Nam, G.H.; Shin, H.; Song, J.R.; Kim, T.J. Functional Expression and Characterization of Acetyl Xylan Esterases CE Family 7 from Lactobacillus Antri and Bacillus Halodurans. J. Microbiol. Biotechnol. 2020, 30, 155–162. [Google Scholar] [CrossRef]
- McKay, A.M. Microbial Carboxylic Ester Hydrolases (EC 3.1.1) in Food Biotechnology. Lett. Appl. Microbiol. 1993, 16, 1–6. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. Microbial Carboxyl Esterases: Classification, Properties and Application in Biocatalysis. FEMS Microbiol. Rev. 2002, 26, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kyu Kim, K.; Kyu Song, H.; Hae Shin, D.; Yeon Hwang, K.; Choe, S.; Joon Yoo, O.; Won Suh, S. Crystal Structure of Carboxylesterase from Pseudomonas Fluorescens, an a/b Hydrolase with Broad Substrate Specificity. Structure 1997, 5, 1571–1584. [Google Scholar] [CrossRef] [Green Version]
- Khan, F.I.; Lan, D.; Durrani, R.; Huan, W.; Zhao, Z.; Wang, Y. The Lid Domain in Lipases: Structural and Functional Determinant of Enzymatic Properties. Front. Bioeng. Biotechnol. 2017, 5, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verger, R. ‘Interfacial Activation’ of Lipases: Facts and Artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Sood, S.; Sharma, A.; Sharma, N.; Kanwar, S.S. Carboxylesterases: Sources, Characterization and Broader Applications. Insights Enzym. Res. 2018, 1, 1. [Google Scholar] [CrossRef]
- Li, X.; Griffin, K.; Langeveld, S.; Frommhagen, M.; Underlin, E.N.; Kabel, M.A.; de Vries, R.P.; Dilokpimol, A. Functional Validation of Two Fungal Subfamilies in Carbohydrate Esterase Family 1 by Biochemical Characterization of Esterases From Uncharacterized Branches. Front. Bioeng. Biotechnol. 2020, 8, 694. [Google Scholar] [CrossRef]
- Ali, S.; Mahmood, S. Mutagenesis of a Thermophilic Alkalibacillus Flavidus for Enhanced Production of an Extracellular Acetyl Xylan Esterase in Semi-Solid Culture of Linseed Meal. Waste Biomass Valorization 2020, 11, 3327–3335. [Google Scholar] [CrossRef]
- Qaseem, M.F.; Wu, A.M. Balanced Xylan Acetylation Is the Key Regulator of Plant Growth and Development, and Cell Wall Structure and for Industrial Utilization. Int. J. Mol. Sci. 2020, 21, 7875. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Verkerk, B.; Bellemare, A.; Lavallee, M.; Frommhagen, M.; Underlin, E.N.; Kabel, M.A.; Powlowski, J.; Tsang, A.; de Vries, R.P. Characterization of New Fungal Carbohydrate Esterase Family 1 Proteins Leads to the Discovery of Two Novel Dual Feruloyl/Acetyl Xylan Esterases. 2020. Available online: https://www.researchsquare.com/article/rs-17222/latest.pdf (accessed on 1 March 2022).
- Yin, C.-F.; Xu, Y.; Deng, S.-K.; Yue, W.-L.; Zhou, N.-Y. A Novel Esterase, DacA Pva, from Comamonas Sp. Strain NyZ500 with Deacetylation Activity for the Acetylated Polymer Polyvinyl Alcohol. Appl Environ Microbiol. 2021, 8, e03016-20. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Mirończuk, A.M.; García-Martín, A.; Saborido, A.; de la Mata, I.; Arroyo, M. Biochemical Properties and Biotechnological Applications of Microbial Enzymes Involved in the Degradation of Polyester-Type Plastics. Biochim. Et Biophys. Acta-Proteins Proteom. 2020, 1868, 140315. [Google Scholar] [CrossRef] [PubMed]
- Koseki, T.; Furuse, S.; Iwano, K.; Sakai, H.; Matsuzawa, H. An Aspergillus Awamori Acetylesterase: Purification of the Enzyme, and Cloning and Sequencing of the Gene. Biochem. J. 1997, 326, 485–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.-J.; Park, S.-M.; Kim, H.-R.; Yang, M.-S.; Kim, D.-H. Cloning the Gene Encoding Acetyl Xylan Esterase from Aspergillus Ficuum and Its Expression in Pichia Pastoris. Enzym. Microb. Technol. 2002, 31, 384–391. [Google Scholar] [CrossRef]
- Koseki, T.; Miwa, Y.; Akao, T.; Akita, O.; Hashizume, K. An Aspergillus Oryzae Acetyl Xylan Esterase: Molecular Cloning and Characteristics of Recombinant Enzyme Expressed in Pichia Pastoris. J. Biotechnol. 2006, 121, 381–389. [Google Scholar] [CrossRef]
- Degrassi, G.; Okeke, B.C.; Bruschi, C.V.; Venturi, V. Purification and Characterization of an Acetyl Xylan Esterase from Bacillus Pumilus. Appl. Environ. Microbiol. 1998, 64, 789–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levisson, M.; Han, G.W.; Deller, M.C.; Xu, Q.; Biely, P.; Hendriks, S.; ten Eyck, L.F.; Flensburg, C.; Roversi, P.; Miller, M.D.; et al. Functional and Structural Characterization of a Thermostable Acetyl Esterase from Thermotoga Maritima. Proteins: Struct. Funct. Bioinform. 2012, 80, 1545–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drzewiecki, K.; Angelov, A.; Ballschmiter, M.; Tiefenbach, K.J.; Sterner, R.; Liebl, W. Hyperthermostable Acetyl Xylan Esterase. Microb. Biotechnol. 2010, 3, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Shao, W.; Wiegel, J. Purification and Characterization of Two Thermostable Acetyl Xylan Esterases from Thermoanaerobacterium sp. Strain JW/SL-YS485. Appl. Environ. Microbiol. 1995, 61, 729–733. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 6, 1547–1549. [Google Scholar] [CrossRef]
- Ontañon, O.M.; Ghio, S.; Marrero Díaz de Villegas, R.; Piccinni, F.E.; Talia, P.M.; Cerutti, M.L.; Campos, E. EcXyl43 β-Xylosidase: Molecular Modeling, Activity on Natural and Artificial Substrates, and Synergism with Endoxylanases for Lignocellulose Deconstruction. Appl. Microbiol. Biotechnol. 2018, 102, 6959–6971. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. Protein Structure and Function Prediction Using I-TASSER. Curr. Protoc. Bioinform. 2015, 52, 5.8.1–5.8.15. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicka-Woś, A.; Henneberger, R.; Batista-García, R.A.; Martínez-Ávila, L.; Jackson, S.A.; Kennedy, J.; Dobson, A.D.W. Biochemical Characterization of a Novel Monospecific Endo-β-1,4-Glucanase Belonging to GH Family 5 from a Rhizosphere Metagenomic Library. Front. Microbiol. 2019, 10, 1342. [Google Scholar] [CrossRef] [PubMed]




| CAZy Family | Activity * | No. of Genes |
|---|---|---|
| CE 1 | Acetyl xylan esterase | 120 |
| CE 10 | Esterase | 38 |
| GH 43 | β-Xylosidase, α-L-arabinofuranosidase, xylanase and arabinanase | 55 |
| GH 5 | Endo-β-1,4-xylanase, β-mannosidase | 15 |
| GH 12 | Endoglucanase, xyloglucan hydrolase | 13 |
| GH 16 | Xyloglucanse, endo-1,3-β-glucanase | 117 |
| GH 10 | Endo-1,4-β-xylanase, endo-1,3-β-xylanase | 16 |
| GH 67 | α-Glucuronidase, xylan α-1,2-glucuronidase | 1 |
| GH 51 | Endoglucanase, endo-β-1,4-xylanase, β-xylosidase, α-L-arabinofuranosidase | 3 |
| CE 6 and CE 7 | Acetyl xylan esterase | 5 |
| GH 8 | Endo-1,4-β-xylanase | 11 |
| GH 38 | α-Mannosidase | 39 |
| GH 39 | β-Xylosidase | 279 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madubuike, H.; Ferry, N. Characterisation of a Novel Acetyl Xylan Esterase (BaAXE) Screened from the Gut Microbiota of the Common Black Slug (Arion ater). Molecules 2022, 27, 2999. https://doi.org/10.3390/molecules27092999
Madubuike H, Ferry N. Characterisation of a Novel Acetyl Xylan Esterase (BaAXE) Screened from the Gut Microbiota of the Common Black Slug (Arion ater). Molecules. 2022; 27(9):2999. https://doi.org/10.3390/molecules27092999
Chicago/Turabian StyleMadubuike, Henry, and Natalie Ferry. 2022. "Characterisation of a Novel Acetyl Xylan Esterase (BaAXE) Screened from the Gut Microbiota of the Common Black Slug (Arion ater)" Molecules 27, no. 9: 2999. https://doi.org/10.3390/molecules27092999
APA StyleMadubuike, H., & Ferry, N. (2022). Characterisation of a Novel Acetyl Xylan Esterase (BaAXE) Screened from the Gut Microbiota of the Common Black Slug (Arion ater). Molecules, 27(9), 2999. https://doi.org/10.3390/molecules27092999