Abstract
Cancer is a multifactorial organic dysfunction for which great efforts are being devoted in searching for new treatments and therapeutic adjuvants. Annona muricata is a fruit that has promising activity against several types of cancer, as it contains acetogenins, the metabolite group associated with this action. Thus, the objective of this study was to evaluate, in experimental models, the toxic behavior of an extract and fraction rich in acetogenins from A. muricata seeds and study the acetogenin, Annonacin, in silico. Phytochemical characterization was made by thin layer chromatography, spectroscopy in the infrared region and nuclear magnetic resonance. Toxicity was evaluated by tests of Allium cepa and Artemia salina, and in silico studies using the SwissDock servers DockThor, PharmMapper, ADMETLab, PreADME, Osiris and ProTox. The extract and fraction showed genotoxic activity against meristematic cells of A. cepa, reducing the mitotic index; however, the extract produced great deleterious effects on the system, even causing cell necrosis. In A. Saline, the extract was more toxic than the fraction, but both samples were considered toxic. Annonacin was effectively linked to complex I, and presented different activities regarding toxicity. Thus, the results of this study are promising, highlighting the anticancer potential of acetogenins.
1. Introduction
Cancer is a disease defined by abnormal cell proliferation resulting from multiple factors. The National Cancer Institute (INCA) estimates that between 2020 and 2022, there will be 625,000 new cases of cancer (non-melanoma skin). Conventional antineoplastic therapies tend to present disadvantages in the treatment adhesion, toxicity, and general improvement of patients’ quality of life [1]. Research to develop or characterize new antineoplastic or adjuvant therapies are essential in terms of the aforementioned aspects. Therapeutic searches using plant species can be important in developing new medicines. Phytochemical analyzes elucidate new compounds that produce desired therapeutic actions, opening a range of possibilities [2].
Annona muricata is a plant belonging to the Annonaceae family of the Magnoliales order. It has great economic and cultural value, and is used in various culinary preparations. In the pharmacological field, its medicinal use is highly widespread in Central and South America, as well as countries in Africa and Asia. Among its popular uses are treatments for bone disorders, neuralgia, digestive disorders, febrile illnesses, abrasions, antiparasitic, cystitis, diabetes, headaches, insomnia, abscesses, ulcers caused by leishmaniasis, conditions associated with cancer, and as an insecticide, larvicidal and piscicidal agent [3].
In vivo studies have shown that A. muricata leaf extracts have an anti-rheumatic effect by decreasing the production of pro-inflammatory cytokines. Another study reached the same result using fruits with antinoceptive activity that results from a decrease of mediators and activity of the opioidergic pathway. Antiparasitic activity was demonstrated in vitro against Trypanosoma cruzi and against several strains of Leishmania, using, in the latter case, mainly isolated acetogenins. Among the bioactive compounds isolated from A. muricata, alkaloids and acetogenins stand out, and megastigmans, flavonoids, phenols and cyclopeptides can also be found [4].
As much as A. muricata is recognized for generating these numerous metabolite groups, in recent years, studies have shifted from alkaloids to acetogenins. This highly bioactive group includes compounds obtained by the acetic acid-polyketide derivatives of long-chain fatty acids. In addition, these aliphatic chains have functional groups attached to them, such as hydroxyl, acetyl and carbonyl groups, as well as a terminal y-lactone ring, tetrahydrofuran, or tetrahydropyran rings. The structural organization of these functional groups has been used to establish the cytotoxic potential of acetogenins. The hydroxyl groups, the presence of the THF ring, and α,β-unsaturated y-lactone subunit, are essential to produce the toxic effect of these compounds, in addition to their concentration in the extract [3].
Studies using extracts from different parts of Annona muricata showed that the fruit has targeted activity against different neoplasms, with acetogenins the main group responsible for this activity. The ability of these secondary metabolites to produce cytotoxic effects in carcinogenic cells is mainly, but not exclusively, linked to their ability to inhibit mitochondrial Complex I [5], which is the enzyme responsible for initiating the process of cellular respiration. This inhibition may lead to cell death of neoplastic cells [6,7].
Studies that assess the toxic capacity of a compound, including cytotoxicity and genotoxicity studies, are essential to determine the safety profile and future uses of the compound via its pharmacological action. The in vitro and in vivo evaluation of extracts or isolates from plant species has become a powerful sieve in the selection of bioactive agents capable of becoming new drugs. In addition, in silico studies provide evidence that can be investigated through other experimental assays, which allows creating a "shortcut" that is based on a complex series of algorithms. Making small changes in the structure of these chemical compounds, making them more active, selective and less toxic, can lead to a reduction in the production costs of a new drug. This work investigates the genotoxic and toxic effects of an extract and a fraction obtained from A. muricata seeds, and includes an in silico study with Annonacin as a target [4].
2. Results and Discussion
2.1. Preparation of Extract and Fraction, and Phytochemical Characterization
After seed maceration, an oily extract with a mass of 5.28 g was obtained, which represented 17.6% of the initial mass of the extractive process. From this final mass, 3 g was used for liquid-liquid partition, 1.5 g in each and, after exploratory TLC, the dichloromethane fraction (FDSAM) was chosen for the following tests. FDSAM represented most of the mass after partition, and 0.76 g was used in the analysis as a fraction rich in acetogenins.
TLC chromatographic profiles suggested that acetogenins were obtained for both the EESAM (ethanolic extract) and its fraction; however, when observed under ultraviolet light, profiles indicative of alkaloids were also noted (Figure 1). Both alkaloids and acetogenins act as phytochemical markers in Annonaceae. Portion A showed a chromatoplac under ultraviolet light (365 nm), the fluorescent blue bands being suggestive of alkaloids, which was confirmed by the reaction with Dragendorff, as demonstrated in portion C. Portion B demonstrates the presence of acetogenins by the reaction with Kedde, which generates a color between rose and lilac, usually weak, that dissipates in a few seconds, revealing acetogenins that have α, β-unsaturated γ-lactonic ring. As observed in portions B and C, acetogenins and alkaloids appear at the same point of retention, which may indicate there is a mixture of the two metabolite groups. To determine if FDSAM is rich in acetogenins, analyzes were performed by more sensitive methods.

Figure 1.
TLC chromatographic profile of the extract and fraction of A. muricata. EESAM—ethanolic extract from seeds; FDSAM—Dichlomethane fraction from seeds. Mobile phase: Ethyl acetate: Methanol (9:1). Developers: (A)—365 nm ultraviolet light; (B)—Kedde; (C)—Dragendorff. Source: authors.
We observe the presence of C=O deformations in the infrared spectrum (Figure 2) (1745 and 1653 cm−1), indicating the lactone portion of acetogenins, and we also observed the axial deformation of C=C (1458 and 1465 cm−1) and the axial deformation C-O (1060 cm−1) that suggests the portion connected to the THF ring (tetrahydrofuran), while the deformations C-H (3003 cm−1), OH (3338 cm−1), CH2 (2850 cm−1) and CH3 (2920 cm−1) indicated the elongated chain of acetogenins linked to the lactone ring and the THF ring. Thus, the absorptions described here indicate the presence of long chains of fatty acids that point to the presence of acetogenins [8,9].

Figure 2.
Infrared spectra of the extract and fraction of A. muricata. EESAM—ethanolic extract from seeds; FDSAM—dichlomethane fraction from seeds. Source: Authors.
In the 1H NMR spectra for the two samples, we found signals of acetogenins, where the α,β-unsaturated y-lactone subunit was evidenced by δ 1.4; 5.0 and 7.0 or 7.2 for acetogenins with OH in their chain, for carbons linked to hydroxyls in shifts δ 3.0 and signals linked to the presence of the THF ring δ 3.8. We also observed characteristic signals of H linked to olefinic carbons at δ 5.0, as well as a large amount of δ 2.0 shifts that are related both to the lactone moiety and to chemical shifts of hydrogens present in the acetogenins chain (Figure 3 and Figure 4; Table 1) [3,10].

Figure 3.
EESAM 1H NMR spectrum. Source: Authors.
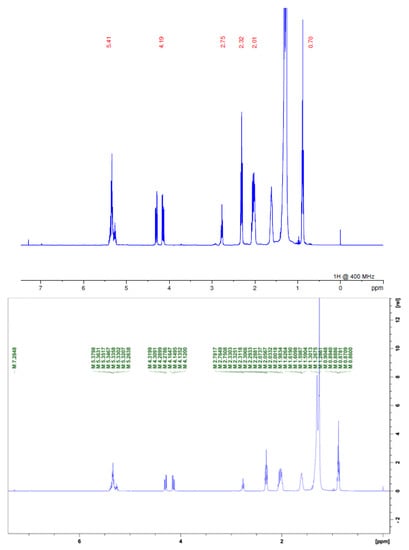
Figure 4.
1H NMR spectrum of FDSAM. Source: Authors.

Table 1.
Values of chemical shifts δ characteristics of Annonacin.
Thus, the extract and the fraction showed the presence of acetogenins in the three tests performed. Studies with A. muricata seeds have generally focused on the presence of acetogenins; however, phytochemical surveys of leaves have demonstrated the presence of a range of other metabolite groups, such as tannins, flavonoids, triterpenes and alkaloids [11]. In our study with seeds, alkaloids and acetogenins were present in this part of the plant, and could be used as pharmacogens in future studies.
2.2. Genotoxicity in Allium cepa
The experimental model of Allium cepa allows the evaluation of the genotoxic effects of organic and inorganic compounds from alterations in mitotic index (MI) and the presence of alterations in the mitotic cycle, multinucleated cells or micronuclei, formation of bridges (linked nuclei), index of aberrations (IA), these being cheap and simple tests [12]. The results generated by such tests, as in this study, allow us to understand the mechanism of cytotoxic effects already described for extracts of A. muricata.
In this study, the extract and fraction demonstrated genotoxic and cytotoxic actions by decreasing MI and increasing IA. EESAM and FDSAM showed, at the lowest concentrations, an elevation of the MI that decreased as the concentration increased and the time elapsed, which may indicate they are time-dose dependent compounds. The elevation of MI compared to the positive control indicates a cytotoxic effect based on promotion of cell growth and eventual tumorization, while a decrease of MI indicates antiproliferative characteristics of the compounds [13], as observed samples in Table 2, which tend to decrease cell proliferation.
Table 2.
Mitotic index and cellular aberrations in Allium cepa induced by the extract and fraction of Annona muricata.
Other studies using A. cepa, showed that the extracts of A. muricata have a cytotoxic effect; however, many of these studies used the leaves or bark [14,15,16,17,18,19], and only in a few studies were seeds and fractions rich in acetogenins used, which, in recent years, have been associated with the antitumor effects of A. muricata [20].
Studies have demonstrated the cytotoxic activity of acetogenins isolated from A. muricata against tumor strains demonstrated with IC50 values ranging from 8 × 10−3–3.3 × 10−1 µg/mL, i.e., against A-549 (human lung carcinoma) find activities for Annomurin with A, B, C and E; Annomutacin, Annopentocin with A, B and C; Arianacin, cis-annonacin, cis-annonacin-10-one, cis-goniothalamicin, cis-trans-annomuricin-D-one, Goniotha-lamycin, Javoricin, Muricatetrocin C, Muricatocin with A, B and C; and Muricoreacin and Murihexocin with A, B and C. In MCF-7 (mammary carcinoma) the same acetogenins added to 2.4-cis and trans-10R-annonacin-A-one presented IC50 values between 5.70 × 10−2 and 17.93 µg/mL. Against HT-29 (human colon adenocarcinoma), the same isolates showed an IC50 of 9 × 10−4–4.0 µg/mL. IC50 values ranged from 1.71 × 10−2–1.14 µg/mL for PC-3 (prostate adenocarcinoma) Annomuricin with E, and Annopentocin with A, B and C; and for cis-trans-annomuricin-D-one, Muricapentocin, Muricoreacin, and Murihexocin with A, B and C [21].
When comparing the results, there was no statistical difference between the positive control and the samples, demonstrating the similarity between the toxic effects of colchicine, EESAM and FDSAM. Colchicine has been evaluated as an inducer of chromosomal alterations due to meiotic changes, mainly in chromosomal adherence and lag, and abnormalities related to mitotic spindle dysfunction [22].
Studies that evaluated the mechanism of action by which acetogenins exert their cytotoxic activity pointed to an inhibition of the NADH-ubiquinone oxidoreductase complex, where acetogenins act by inhibiting electron transport by blocking complex I [23,24]. The electron transport chain produces energy for cells by the oxidative phosphorylation of glucose and fatty acids. Complex I reduces NADH to NAD+, in addition to ubiquinone to ubiquinol, and carries protons through the inner mitochondrial membrane through an imbalance of the redox potential. In this way, cells that require a large amount of energy for division, such as neoplastic cells, would be strongly affected by this effect [5,25].
As observed by this study, EESAM and FDSAM produced deleterious effects on cells, leading to visible damage in the cellular environment. The extract and the fraction showed vagrant chromosomes, chromosomal loss, telophase bridges and micronuclei (Figure 5 and Figure 6), which indicates there were alterations in the mitotic process. EESAM produced harmful effects on A. cepa meristem cells such as loss of shape, chromosomal viscosity and necrosis. In this sense, the fractionation process maintained the cytotoxic effects without damage caused by the extract that could come from a synergistic effect with other, non-alkaloid compounds existing in the seed.

Figure 5.
Chromosomal aberrations caused by the extract from A. muricata seeds. Source: Author, 2022. (A-1–2): Wandering chromosomes; (B-1): chromosomal loss; (C-1–2): micronuclei; (D-1): telophase bridge; (E-1): shape changes; (F-1): viscosity; (G-1): necrosis—detected by the presence of vacuoles in the cytoplasm.

Figure 6.
Chromosomal aberrations caused by the fraction from A. muricata seeds. Source: Author, 2022. (A-1): Wandering chromosomes; (B-1–2): chromosomal loss; (C-1–2): micronuclei; (D-1): telophase bridge.
2.3. Toxicity in the Artemia salina Leach
Seeking to expand the results discussed in this study, the toxicity test was performed against Artemia salina. The same concentrations used in the genotoxicity test were used to observe how the samples would behave in a more complex organism. EESAM had an LC50 value of 1.26 µg/mL, while that of FDSAM was LC50 = 4.58 µg/mL (Figure 7). In the consulted literature, there were no studies that used extracts or fractions of seeds in concentrations close to those of our study. However, Luna et al. [26] found CL50 = 0.49 µg mL−1 for leaf extracts. Another study showed the higher the concentration, the higher the percentage of deaths, thus demonstrating a concentration-dependent action [27]. Studies by Hoe et al. [28] showed that extracts rich in acetogenins are more toxic against A. salina larvae, confirming the results presented in this study.

Figure 7.
Determination of lethal dose in A. salina larvae. Source: Authors.
2.4. In Silico Study
In silico studies have emerged as a prerequisite for the development of new drugs or analyzes of molecular changes made to existing drugs. In this study, we studied Annonacin, an acetogenin isolated from the seeds of A. muricata, as a target. This molecule was chosen because it shows signals in the infrared spectrum and the 1H NMR method used in this study [3,10]
When performing molecular anchoring targeting with Complex I (Figure 7), we obtained an affinity of −10.72 Kcal/mol (T. Energy −54.543 Kcal/mol) on SwissDock and −8.13 Kcal/mol (T. Energy −64.897 Kcal/mol) on the DockThor. The activity of a molecule is related to the energy it gives off to bind to a target. In this relationship, a more stable complex requires less energy for the interaction to occur [28]. Thus, the negative values found in this study demonstrate Annonacin, theoretically, effectively binds to mitochondrial Complex I, according to our hypothesis.
By simulating the bonds that Annonacin can make with Complex I (Figure 8) we obtained evidence for the presence of two hydrogen bonds, one conventional with a Glycine-Gly89 residue (Å 3.03), and another hydrogen pi-bridge with Fenylalamine-Phe93 (Å 3.09) and two hydrophobic alkyl ones with residues Alanina-Ala91 (Å 4.47) and Ala426 (Å 4.04) (Figure 9). The smaller the distance between the amino acid residues and the ligand, the stronger the bond, even if these bonds are weak, as with hydrogen bonds [29]. The distances observed in this study, up to 4.5 Å, indicate good coupling between Annonacin and Complex I [24].

Figure 8.
2D representation of molecular anchorage obtained by SwissDock. Source: Authors.

Figure 9.
Simulation of connections made between the receptor-ligand. Source: Authors.
When generating reverse pharmacophoric mapping, we obtained 300 possible targets for Annonacin. The first 10 were studied in more detail and are presented in Table 3, with emphasis on GTPase HRAS, transthyretin, mitogen-activated protein kinase and methionine aminopeptidase 2, which were chosen because they relate, directly or indirectly, to carcinogenic processes or/and tumor deactivation, which was the subject of this in silico study.
Table 3.
Prediction of possible targets for Annonacin.
The first predicted target was the GTPase HRAS, an isoform of the GTPase RAS protein expressed in several types of cancer. RAS proteins are considered “undruggable” because, in addition to not having favorable binding sites, they make high-affinity bonds with nucleotides. Thus, drugs that seek to inhibit this pathway competitively inhibit the docking of RAS-bound GTP to the membrane. Drugs that block GTP are prime targets in the search for new antineoplastic therapies [30,31].
The second target was Transthyretin (TTR). Another protein involved in the development of cancer, it is one of the biomarkers of cell proliferation. In lung cancer, evidence was found that recombinant TTR increased the proliferation of carcinogenic cells acting by several mechanisms, such as increased CLL melanoma cells, regulation of immune cells and promotion of angiogenesis. In addition, it acts as a carrier of cellular retinoic acid binding protein (third predicted target) [32,33]. Thus, drugs that modulate TTR can be used in specific cases of lung cancer.
The seventh predicted target was the mitogen-activated protein kinase (MAPK) family, specifically p38 MAP kinase, which shows dual characteristics in cancer pathogenesis. p38 acts both in the G1/S and G2/M phases by checkpoint responses. As an antitumor, p38 has mechanisms of negative regulation of Cyclin D1 and, through cellular senescence, as a pro-oncogenesis, it increases resistance, survival and even assists migration (metastasis) of cancer cells from the isoform p38γ [34,35]. Therefore, they may be targets of new therapies, as long as they are selective for the p38γ isoform.
Methionine aminopeptidase 2 (MetAP2) was the eighth target predicted in this study, and is related to tumor activities. MetAP2 is a bifunctional enzyme that removes the amino-terminal methionine from newly formed proteins. MetAP2 seems to be related to the proliferation of endothelial cells at the stage of tumor angiogenesis, and has thus become a popular target for antitumor drugs [36,37].
Table 4 shows toxicity predictions for Annonacin in different servers. We found resonances between the predictions made for hepatotoxicity, hERG inhibition (cardiotoxicity), irritant effects and mutagenicity; however, we found dissonant results for carcinogenesis. While Osiris and ProTox showed negative results, PreADME showed positivity in mice, which were more sensitive models than rats, in which a range of toxic substances were generated and resulted in a prediction of carcinogenicity [38]. PreADME was also predicted to be toxic to algae, Medaka, Daphnia and Minnow; however, highly lipophilic compounds tend to be predicted as toxic in these models because of their natural accumulation [39]. Studies using Annonacin focused mainly on evaluating its mutagenic action, being negative in these studies [40,41].
Table 4.
Annonacin toxicity prediction.
Studies have shown that Annonacin can have a neurotoxic effect, inducing an atypical Parkinsonian state. This could occur due to these compounds being deposited in cells of the nervous system, delays in the mitochondrial distribution of ATPs in the cell soma and interference in the intracellular distribution of the protein tau [42,43,44]. The authors of the studies were not categorical regarding this action, and there may even be synergism for the toxic effect [42]. In addition, preliminary studies have demonstrated that even though acetogeins, such as Annonacin, can cross the blood-brain barrier, there is low brain exposure to acetogenins at doses of 10 mg/kg [45].
Thus, what is envisaged is a compound with high potential for use against neoplasms. However, more detailed studies are needed not only on the pharmacokinetics of this compound, but also on the pre-formulation of its possible pharmaceutical form, to guarantee its efficacy and safety as a therapy, as well as molecular modeling studies in which its therapeutic effect can be guaranteed, reducing its toxicity.
3. Materials and Methods
3.1. Plant Material, Extract and Fractions
A. muricata seeds were donated by Miriam Beta. They came from the municipality of Castanhal—PA. They were sanitized, dried and ground. An exsiccata was prepared and it is in the process of being integrated into the Herbarium Prof. Dr. Marlene Freitas da Silva—Center for Social Sciences, at the University of the State of Pará.
The ground seeds (±30 g) were subjected to exhaustive maceration with 96° GL ethanol for 21 days. Every 7 days the extractive solution was filtered, and fresh solvent was added. At the end of 21 days, the extractive solutions were concentrated in a rotary evaporator under reduced pressure, obtaining EESAM from the seeds. To obtain the acetogenin-rich fractions, liquid-liquid partitions were carried out, where 1.5 mg of EESAM was solubilized in methanol and water (1:1) and extracted with organic solvents, as in the scheme described in Figure 10.
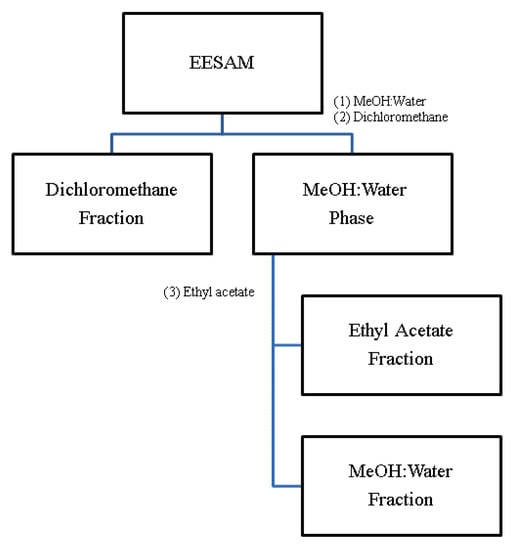
Figure 10.
Proposed partition scheme. Source: Silva, 2017 (Adapted).
3.2. Thin Layer Chromatography—TLC
Silica gel was used as the stationary phase, and 18 mL of ethyl acetate and 2 mL of methanol (9:1 ratio) as a mobile phase. The reagents were: Kedde (for solution A (3.5 dihydrobenzoic acid (2 g) was mixed in methanol (50 mL)); for solution B (potassium hydroxide (5.7 g) was dissolved in methanol (100 mL)) and solutions were mixed in a 1:1 ratio at the time of use) and Dragendorff (bismuth subnitrate (8 g) potassium iodide (27 g) was mixed with concentrated hydrochloric acid (20 mL) and distilled water qsp. (100 mL)) [3].
3.3. Spectroscopy in the Infrared Region—Fourier Transform (FT-IR)
Spectra were obtained using an IR Prestige-21 Shimadzu spectrophotometer, in which 1 mL of the samples was dispersed over a diamond crystal with attenuated total reflectance (ATR), and 20 scans were performed with a resolution of 2 cm−1 in the spectral range of 4000–400 cm−1.
3.4. 1H Nuclear Magnetic Resonance
NMR analyzes were performed on a Bruker Ascend 400 spectrometer (operating at 400 MHz for hydrogen). Samples were solubilized in chloroform. Chemical shifts (δ) were measured in ppm and coupling constants (J) in Hertz (Hz). Tetramethylsilane (TMS) was used as an internal reference.
3.5. Allium cepa Assay
About 1 mg of A. cepa seeds was placed in a petri dish lined with filter paper and treated with different concentrations of ethanol extract and dichloromethane fraction from A. muricata seeds (25; 12.5; 6.25; 3.125 and 1.5625 µg/mL). After radial growth of 1 cm of the seeds, they were removed at 24, 48 and 72 h and were counted and fixed in Carnoy (absolute ethyl alcohol: glacial acetic acid 3:1).
For the control groups, distilled water (negative control) and colchicine (positive control) at the same concentrations of the extract and fraction were used. To prepare the slides, the shoots were removed from the fixative and transferred to a petri dish where they were washed (3 washes of 5 min). After that, they were subjected to acid hydrolysis in 1N hydrochloric acid for 15 min, then washed again and stained in acetic acid orcein 2% for 10 min. Following this, the shoots were placed on a slide and a drop of acetic orcein was placed on the cut. After 1 min, crushing (squash) was performed, and the slides were observed under the light microscope at 400× magnification [12]. The results were analyzed with the PAST program (Paleontological Statistics) using ANOVA followed by Tukey’s test for multiple comparisons. Values in which the p-value was less than or equal to 0.05 were considered statistically significant.
3.6. Artemia Salina Leach Assay
To hatch A. salina cysts, a solution of sea salt and distilled water at a concentration of 35 g/L was used. The pH of the solution was adjusted to 9–10 with sodium hydroxide (NaOH) at 0.1 mol/L, then the solution was homogenized and filtered. The saline environment was transferred to an aquarium with a light/dark environment, where 20 mg of A. salina cysts were added and remained until complete hatching cysts (±24 h) at a temperature of 27–30 °C. The extract and the fraction were prepared at the same concentration as the A. cepa assay with a final volume of 5 mL. This assay was carried out with five replicates, with 10 crustaceans being added to each and kept for 24 h and 48 h. The live and dead nauplius were counted at the end of each cycle [46,47] (both with adaptation). To calculate the Lethal Dose 50% (LD50) the IC50 Calculator tool [48] was used.
3.7. In Silico Study
The following programs were used in the in silico study: ChemSketch (version 2.1, 2019), to design the chemical structure of the Annonacin, which was subjected to molecular anchoring using the SwissDock (version 2022), and DockThor (version 2.0) against coenzyme NADH: ubiquinone oxidoreductase—Complex I (ID: 5XTB). These online servers were comprised within the fast-docking system. The Dockthor server was powered by the National Laboratory of Scientific Computing, which works with the C++ language, using generic algorithms based on the molecular force field MMFF94 (Merck Molecular Force Field). SwissDock, on the other hand, is a server that is confined to a system based on multiobjective scoring function designed around the CHARMM22 force field and the FACTS solvation model. The coenzyme and the ligand were optimized in the Discovery Studio program. In the program, the forces involved in the docking, receptor-ligand interactions, as distance and binding types, were observed. In addition to reverse pharmacophoric mapping using the PharmMapper server (2008–2022), PreADMET (2012–2021), ADMETLab (version 2.0) and Protox (2021), were used in addition to free software OSIRIS Property Explore (2017) [49,50,51,52,53].
4. Conclusions
Both EESAM and FDSAM showed genotoxic action against A. cepa and were toxic against A. salina; however, EESAM showed greater deleterious effects on cell division and microcrustaceans, which may result from synergistic action or the presence of specific toxicant groups. From the assays developed in this study, and the available literature, it is evident that acetogenins are a group of compounds that can be tested as drug candidates based on in vivo seminal studies and, in the future, may be used in broad or narrow antineoplastic therapy.
Author Contributions
Experiment, data processing, writing, G.G.F.; validation of methodology, A.C.S.Q.; NMR spectrum, A.M.d.R.M. and J.E.d.S.S.; FT-IR analysis, K.L.C. and J.O.C.S.-J.; review, S.P.; supervision, D.L.d.N.B. and M.F.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Edital Universal 2018 (Processo 432458/2018.2), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—CAPES, Brazil. PROPESP/UFPA support the publication of this article.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from the corresponding author upon request.
Acknowledgments
The authors extend their appreciation to the Federal University of Pará for granting the scientific initiation scholarship.
Conflicts of Interest
The authors declare no conflict of interest.
Sample Availability
Samples of the compounds are not available from authors.
References
- Instituto Nacional de Câncer José Alencar Gomes da Silva. Estimativa 2020: Incidência de Câncer No Brasil; Instituto Nacional de Câncer José Alencar Gomes da Silva INCA: Rio de Janeiro, Brazil, 2019. Available online: https://www.inca.gov.br/sites/ufu.sti.inca.local/files//media/document//estimativa-2020-incidencia-de-cancer-no-brasil.pdf (accessed on 22 July 2022).
- Vieira, V.; Cruz, V.; Soares, N.; Silva, J.; Araújo, E. Quimioterápicos antineoplásicos derivados de plantas. Enciclopédia Biosf. 2020, 17, 444–461. [Google Scholar] [CrossRef]
- Silva, T.F. Análise Fitoquímica e Atividade Leishmanicida de Frações de Sementes de Annona muricata L. (annonaceae) de um Cultivo em Tomé-açu. Master’s Thesis, Universidade Federal do Pará, Pará, Brazil, 2017. [Google Scholar]
- Moghadamtousi, S.Z.; Fadaeinasab, M.; Nikzad, S.; Mohan, G.; Ali, H.M.; Kadir, H.A. Annona muricata (Annonaceae): A Review of Its Traditional Uses, Isolated Acetogenins and Biological Activities. Int. J. Mol. Sci. 2015, 16, 15625–15658. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.K.; Lu, J.; Bai, Y. Mitochondrial Respiratory Complex I: Structure, Function and Implication in Human Diseases. Curr. Med. Chem. 2009, 16, 1266–1277. [Google Scholar] [CrossRef]
- Lima, N.N.d.C.; Faustino, D.C.; Allahdadi, K.J.; França, L.S.D.A.; Pinto, L.C. Acetogenins from Annonaceae plants: Potent antitumor and neurotoxic compounds. PharmaNutrition 2022, 20, 100295. [Google Scholar] [CrossRef]
- Fontaine, E. Metformin-Induced Mitochondrial Complex I Inhibition: Facts, Uncertainties, and Consequences. Front. Endocrinol. 2018, 9, 753. [Google Scholar] [CrossRef] [PubMed]
- Barreto, F.S. Estudo da Atividade Citotóxica de Compostos Obtidos do Extrato Acetônico das Folhas de Annona muricata L. por Fracionamento Bioguiado. Master’s Thesis, Universidade Federal do Ceára, Ceára, Brazil, 2014. [Google Scholar]
- Nunes, C.R. Estudo Químico e Avaliação da Atividade Antineoplásica de Annona muricata L. Doctoral´s Thesis, Universidade Estadual do Norte Fluminense Darcy Ribeiro, Rio de Janeiro, Brazil, 2015. [Google Scholar]
- Lima, M.D. Perfil Cromatográfico dos Extratos Brutos das Sementes de Annona muricata L. e Annona squamosa L. Através da Cromatografia Líquida de alta Eficiência. Master’s Thesis, Universidade Federal de Alagoas, Maceió, Brazil, 2007. [Google Scholar]
- Simões, C.F.D.S. Perfil Fitoquímico e Estudo das Atividades Antimicrobiana, Citotóxica e Anti-Inflamatória de Annona muricata L. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2015. [Google Scholar]
- Castro, A.L.G.; Cruz, J.N.; Sodré, D.F.; Correa-Barbosa, J.; Azonsivo, R.; de Oliveira, M.S.; Siqueira, J.E.d.S.; Galucio, N.C.D.R.; Bahia, M.D.O.; Burbano, R.M.R.; et al. Evaluation of the genotoxicity and mutagenicity of isoeleutherin and eleutherin isolated from Eleutherine plicata herb. using bioassays and in silico approaches. Arab. J. Chem. 2021, 14, 103084. [Google Scholar] [CrossRef]
- Farias, M.S. Dano ao DNA, Citotoxicidade, Efeito Antiproliferativo e Antitumoral de 1, 4-Naftoquinonas Substituídas. Doctoral’s Thesis, Universidade Federal de Santa Catarina, Florianópolis, Brazil, 2014. [Google Scholar]
- Araujo Soria, J.M. Efecto Protector del Extracto Etanolico de Annona Muricata Sobre la Citotoxicidad y Genotoxicidad del Sorbato de Potasio en Meristemos Radiculares de Allium cepa. Monograph’s Thesis, Universidad Nacional de Trujillo, Trujillo, Peru, 2012. [Google Scholar]
- Chávez Aldave, M.I. Efecto del Extracto Etanólico de Annona Muricata a Diferentes Tiempos y Concentraciones Sobre el Ciclo Celular en Allium Cepa. Doctoral’s Thesis, Universidad Nacional de Trujillo, Trujillo, Peru, 2015. [Google Scholar]
- Bibi, R. Cytotoxic and Genotoxic Efects of Morinda Citrifolia and Annona Muricata on Allium Cepa Root Tip Cells. Master’s Thesis, University of the South Pacific, Suva, Fiji, 2017. [Google Scholar]
- Fajardo, V.G.F. Cytological Study on Allium cepa (onion) root tip treated with Annona muricata (guyabano) bark and leaves extract. Ascend. Asia J. Multidiscip. Res. Abstr. 2018, 2, 3. Available online: https://ojs.aaresearchindex.com/index.php/AAJMRA/article/view/4045 (accessed on 15 July 2022).
- Rehana, B.; Ketan, C.; Marta, F.; Nwangburuka, C. Analysis of the mitotic effect of Annona muricata leaf extracts on Allium cepa root tip cells. Afr. J. Biotechnol. 2019, 18, 155–162. [Google Scholar] [CrossRef]
- Ruiz Baca, J. Efecto de Diferentes Concentraciones del Extracto Clorofórmico de Hojas de Annona muricata L. “Guanábana” y Tiempos de Exposición Sobre el Índice Mitótico de Allium Cepa L. “Cebolla” var. Arequipeña. Monograph´s Thesis, Universidad Nacional de Trujillo, Trujillo, Peru, 2019. [Google Scholar]
- Jacobo-Herrera, N.; Perez-Plasencia, C.; Castro-Torres, V.A.; Martínez-Vázquez, M.; González-Esquinca, A.R.; Zentella-Dehesa, A. Selective Acetogenins and Their Potential as Anticancer Agents. Front. Pharmacol. 2019, 10, 783. [Google Scholar] [CrossRef]
- Paes, M.M.; Vega, M.R.G.; Cortes, D.; Kanashiro, M.M. Potencial citotóxico das acetogeninas do gênero Annona. Rev. Virtual De Química 2016, 8, 945–980. [Google Scholar] [CrossRef]
- Khah, M.A.; Alshehri, M.A.; Filimban, F.Z.; Alam, Q.; Aloufi, S. Influence of Colchicine in Causing Severe Chromosomal Damage in Microsporocytes of Hard Wheat (Triticum durum Desf.): Possible Mechanisms and Genotoxic Relevance. Cytologia 2022, 87, 137–143. [Google Scholar] [CrossRef]
- Zafra-Polo, M.C.; Figadère, B.; Gallardo, T.; Tormo, J.; Cortes, D. Natural acetogenins from annonaceae, synthesis and mechanisms of action. Phytochemistry 1998, 48, 1087–1117. [Google Scholar] [CrossRef]
- Grba, D.N.; Blaza, J.N.; Bridges, H.R.; Agip, A.-N.A.; Yin, Z.; Murai, M.; Miyoshi, H.; Hirst, J. Cryo-electron microscopy reveals how acetogenins inhibit mitochondrial respiratory complex I. J. Biol. Chem. 2022, 298, 1–13. [Google Scholar] [CrossRef]
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial Respiratory Chain Complexes. In Membrane Protein Complexes: Structure and Function. Subcellular Biochemistry; Harris, J., Boekema, E., Eds.; Springer: Singapore, 2018; Volume 87. [Google Scholar] [CrossRef]
- Luna, J.D.S.; De Carvalho, J.M.; De Lima, M.R.F.; Bieber, L.W.; Bento, E.D.S.; Franck, X.; Sant’Ana, A.E.G. Acetogenins in Annona muricata L. (annonaceae) leaves are potent molluscicides. Nat. Prod. Res. 2006, 20, 253–257. [Google Scholar] [CrossRef]
- Silva, E.M.F.; Nascimento, R.B.D.C.; Barreto, F.S.; Moraes Filho, M.O.D.; Griz, S.D.A.S.; Santos, A.F.D.; Mousinho, K.C. Estudo in vitro do potencial citotóxico da Annona muricata L. Rev. De Ciências Farm. Básica Apl. 2015, 36, 277–283. [Google Scholar]
- Hoe, P.K.; Yiu, P.H.; Ee, G.C.L.; Wong, S.C.; Rajan, A.; Bong, C.F. Biological Activity of Annona muricata Seed Extracts. Malays. J. Sci. 2010, 29, 153–159. [Google Scholar] [CrossRef]
- Barros, M.E.S.B. Estudos de Docking Molecular, Síntese e Atividade Biológica de Análogos da (-)-Massoialactona e da Combretastatina A-4. Master’s Thesis, Universidade Federal de Pernambuco, Recife, Brazil, 2015. [Google Scholar]
- Sugita, S.; Enokida, H.; Yoshino, H.; Miyamoto, K.; Yonemori, M.; Sakaguchi, T.; Osako, Y.; Nakagawa, M. HRAS as a potential therapeutic target of salirasib RAS inhibitor in bladder cancer. Int. J. Oncol. 2018, 53, 725–736. [Google Scholar] [CrossRef]
- Hodge, R.G.; Schaefer, A.; Howard, S.V.; Der, C.J. RAS and RHO family GTPase mutations in cancer: Twin sons of different mothers? Crit. Rev. Biochem. Mol. Biol. 2020, 55, 386–407. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.; Sengupta, S.; McComb, M.E.; Théberge, R.; Wilson, W.G.; Costello, C.E.; Jacobsen, D.W. In Vitro and in Vivo Interactions of Homocysteine with Human Plasma Transthyretin. J. Biol. Chem. 2003, 278, 49707–49713. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-C.; Ding, X.; Zhao, T.; Wu, L.; Perkins, S.; Du, H.; Yan, C. Transthyretin Stimulates Tumor Growth through Regulation of Tumor, Immune, and Endothelial Cells. J. Immunol. 2018, 202, 991–1002. [Google Scholar] [CrossRef]
- Haagenson, K.K.; Wu, G.S. Mitogen activated protein kinase phosphatases and cancer. Cancer Biol. Ther. 2010, 9, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Ahmadi, A.; Mortezaee, K. Extracellular-signal-regulated kinase/mitogen-activated protein kinase signaling as a target for cancer therapy: An updated review. Cell Biol. Int. 2019, 43, 1206–1222. [Google Scholar] [CrossRef]
- Selvakumar, P.; Lakshmikuttyamma, A.; Dimmock, J.R.; Sharma, R.K. Methionine aminopeptidase 2 and cancer. Biochim. Et Biophys. Acta 2006, 1765, 148–154. [Google Scholar] [CrossRef]
- Grocin, A.G.; Kallemeijn, W.W.; Tate, E.W. Targeting methionine aminopeptidase 2 in cancer, obesity, and autoimmunity. Trends Pharmacol. Sci. 2021, 42, 870–882. [Google Scholar] [CrossRef]
- Carvalho, L.S. Efeito Depressor e Toxicidade do Extrato Etanólico da Casca de Aspidosperma Subincanum (Apocynaceae) em Camundongos. Master’s Thesis, Universidade Federal de Goiás, Goiânia, Brazil, 2013. [Google Scholar]
- Djouad, S.-E.; Berredjem, M.; Aoul, F.Z.H.; Bouchareb, F.; Guerfi, M.; Ben Hadda, T.; Aissaoui, M.; Belhani, B. In silico drug design and molecular docking of novel amidophosphonates and sulfamidophosphonates as inhibitors of urokinase-type plasminogen activator. J. Indian Chem. Soc. 2022, 99, 100650. [Google Scholar] [CrossRef]
- Vilar, J.B.; Ferreira, F.L.; Ferri, P.H.; Guillo, L.A.; Chen, L.C. Assessment of the mutagenic, antimutagenic and cytotoxic activities of ethanolic extract of araticum (Annona crassiflora Mart. 1841) by micronucleus test in mice. Braz. J. Biol. 2008, 68, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Colom, O.; Salvatore, A.; Willink, E.; Ordóñez, R.; Isla, M.I.; Neske, A.; Bardón, A. Insecticidal, Mutagenic and Genotoxic Evaluation of Annonaceous Acetogenins. Nat. Prod. Commun. 2010, 5, 391–394. [Google Scholar] [CrossRef]
- Champy, P.; Höglinger, G.; Féger, J.; Gleye, C.; Hocquemiller, R.; Laurens, A.; Guérineau, V.; Laprévote, O.; Medja, F.; Lombès, A.; et al. Annonacin, a lipophilic inhibitor of mitochondrial complex I, induces nigral and striatal neurodegeneration in rats: Possible relevance for atypical parkinsonism in Guadeloupe. J. Neurochem. 2003, 88, 63–69. [Google Scholar] [CrossRef]
- Escobar-Khondiker, M.; Hollerhage, M.; Muriel, M.-P.; Champy, P.; Bach, A.; Depienne, C.; Respondek, G.; Yamada, E.S.; Lannuzel, A.; Yagi, T.; et al. Annonacin, a Natural Mitochondrial Complex I Inhibitor, Causes Tau Pathology in Cultured Neurons. J. Neurosci. 2007, 27, 7827–7837. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.; Ryan, S.; McDonald, M.; Thomas, A.L.; Maia, J.G.S.; Smith, R.E. Annonacin and Squamocin Contents of Pawpaw (Asimina triloba) and Marolo (Annona crassiflora) Fruits and Atemoya (A. squamosa × A. cherimola) Seeds. Biol. Trace Elem. Res. 2020, 199, 2320–2329. [Google Scholar] [CrossRef] [PubMed]
- Bonneau, N.; Schmitz-Afonso, I.; Brunelle, A.; Touboul, D.; Champy, P. Method development for quantification of the environmental neurotoxin annonacin in Rat plasma by UPLC–MS/MS and application to a pharmacokinetic study. J. Chromatogr. B 2015, 1004, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.N.; Ferrigni, N.R.; Putnam, J.E.; Jacobsen, L.B.; Nichols, D.E.; McLaughlin, J.L. Brine Shrimp: A Convenient General Bioassay for Active Plant Constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Amarante, C.B.D.; Müller, A.H.; Póvoa, M.M.; Dolabela, M.F. Estudo fitoquímico biomonitorado pelos ensaios de toxicidade frente à Artemia salina e de atividade antiplasmódica do caule de aninga (Montrichardia linifera). Acta Amaz. 2011, 41, 431–434. [Google Scholar] [CrossRef]
- Bioquest, A.A.T. Inc. Quest Graph™ IC50 Calculator; Bioquest, A.A.T. Inc.: Pleasanton, CA, USA, 2020. [Google Scholar]
- Almeida, D.M. Dockthor: Implementação, Aprimoramento e Validação de um Programa de Atracamento Receptor-Ligante. Master’s Thesis, Laboratório Nacional de Computação Científica, Petrópolis, Brazil, 2011. [Google Scholar]
- Bitencourt-Ferreira, G.; Azevedo, W.F.D. Docking with SwissDock. In Docking Screens for Drug Discovery; Humana: New York, NY, USA, 2019; pp. 189–202. [Google Scholar] [CrossRef]
- Ferreira, G.G.; Brandão, D.L.D.N.; Dolabela, M.F. Predição do comportamento farmacocinético, toxicidade e de atividades biológicas de alcaloides isolados de Geissospermum laeve (Vell.) Miers. Res. Soc. Dev. 2020, 9, e27991211056. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef]
- Grosdidier, A.; Zoete, V.; Michielin, O. Fast docking using the CHARMM force field with EADock DSS. J. Comput. Chem. 2011, 32, 2149–2159. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).