
Volatile Short-Chain Aliphatic Aldehydes Act as Taste Modulators through the Orally Expressed Calcium-Sensing Receptor CaSR


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Taste-Enhancing Effects of IVAH on Basic Taste Solution
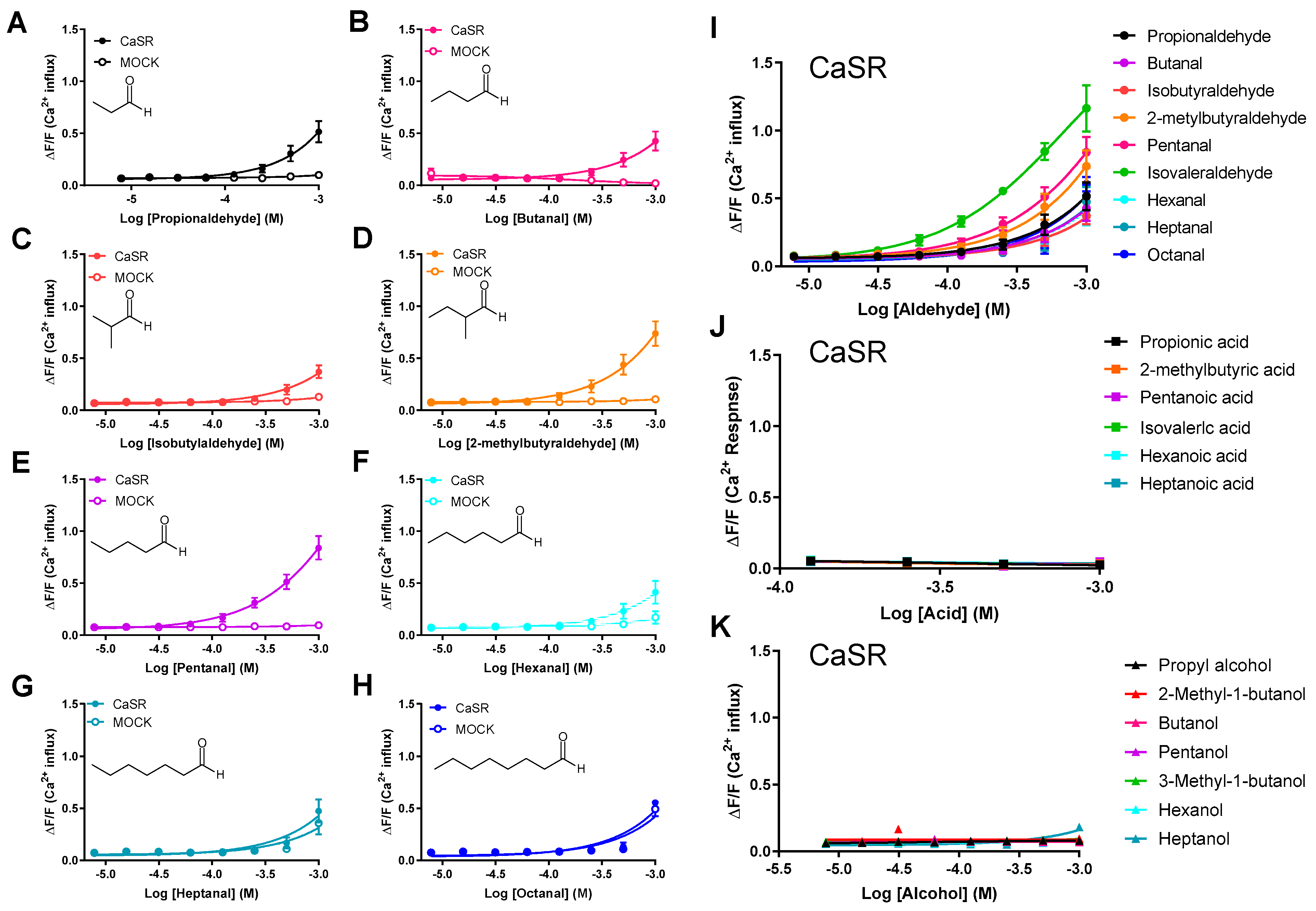
2.2. Human CaSR Responds to Aldehydes
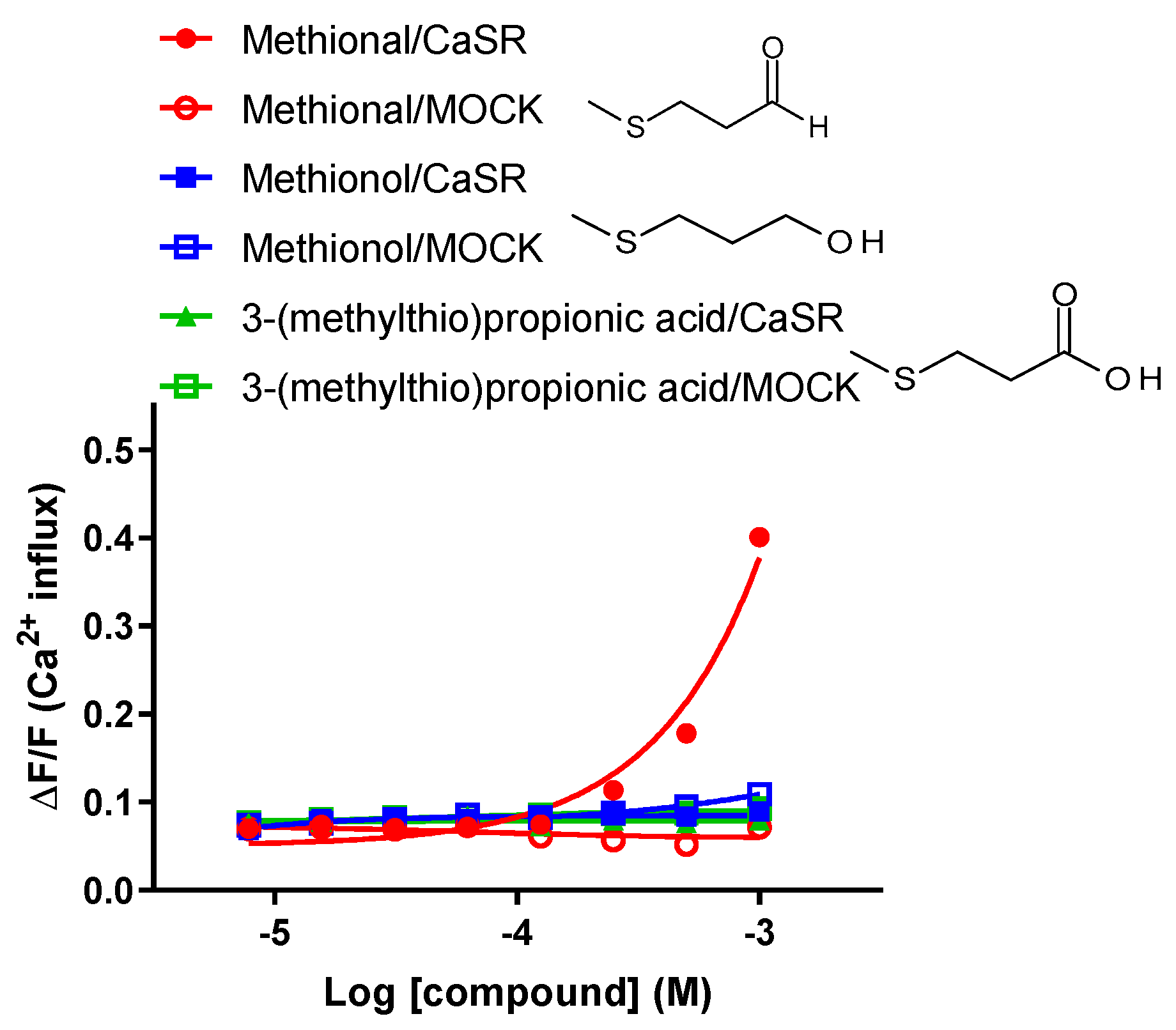
2.3. Human CaSR Responds to the Sulfur-Containing Aldehyde Methional
2.4. IVAH and Methional Enhanced the Responses of CaSR to Calcium Ions
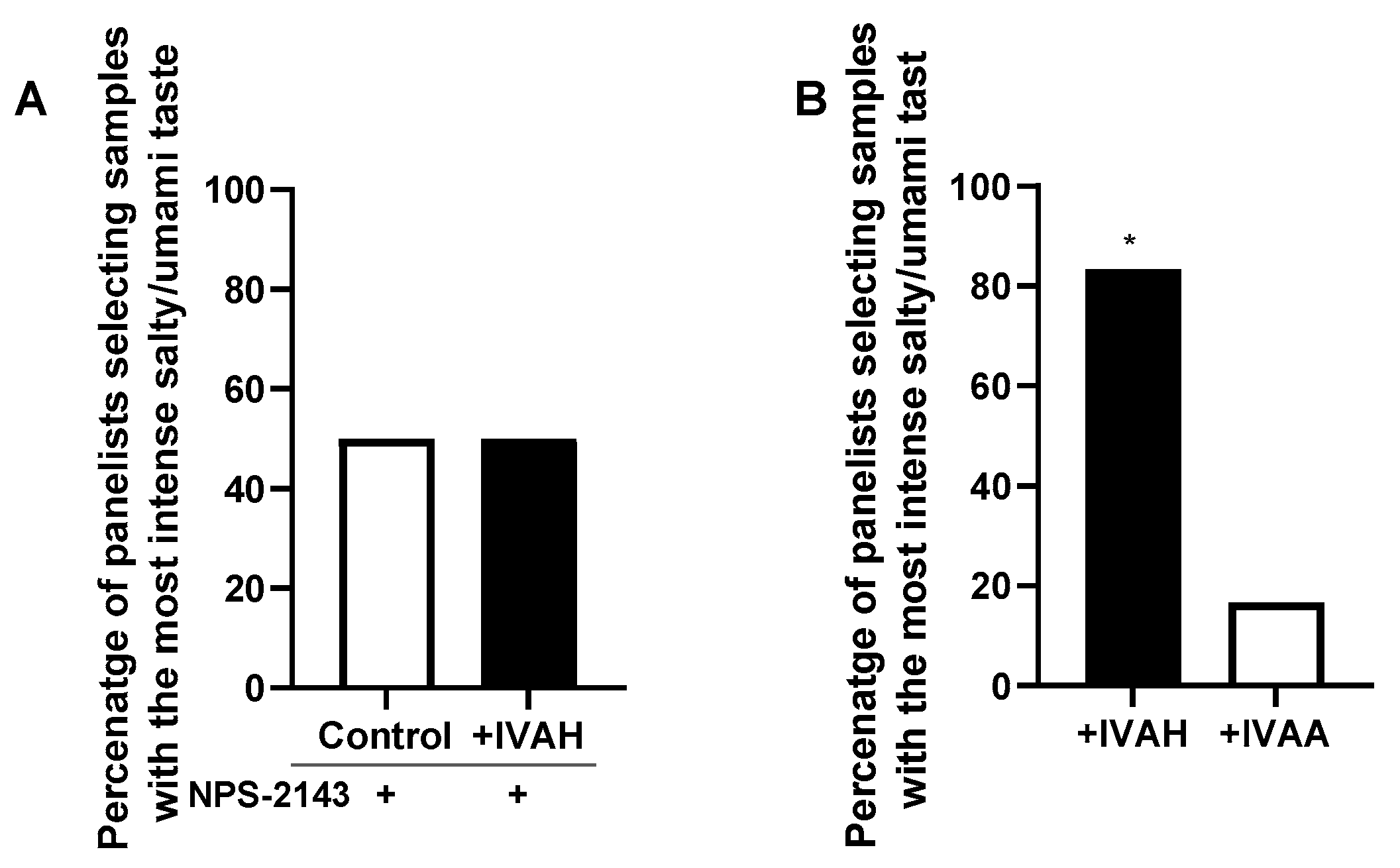
2.5. Relationship between Taste-Enhancing Effects and CaSR Activity
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Measurement of CaSR Activity in CaSR-Expressing PEAKrapid Cells
4.3. Sensory Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fontanini, A. Taste. Curr. Biol. 2023, 33, R130–R135. [Google Scholar] [CrossRef]
- Yoshida, R.; Ninomiya, Y. Taste information derived from T1R-expressing taste cells in mice. Biochem. J. 2016, 473, 525–536. [Google Scholar] [CrossRef]
- Spence, C. Factors affecting odour-induced taste enhancement. Food Qual. Prefer. 2022, 96, 104393. [Google Scholar] [CrossRef]
- Dalton, P.; Doolittle, N.; Nagata, H.; Breslin, P.A. The merging of the senses: Integration of subthreshold taste and smell. Nat. Neurosci. 2000, 3, 431–432. [Google Scholar] [CrossRef]
- Batenburg, M.; Velden, R. Saltiness enhancement by savory aroma compounds. J. Food Sci. 2011, 76, S280–S288. [Google Scholar] [CrossRef]
- Djordjevic, J.; Zatorre, R.J.; Jones-Gotman, M. Odor-induced changes in taste perception. Exp. Brain Res. 2004, 159, 405–408. [Google Scholar] [CrossRef]
- Bonkohara, K.; Fuji, M.; Nakao, A.; Igura, N.; Shimoda, M. Preparation of reminiscent aroma mixture of Japanese soy sauce. Biosci. Biotechnol. Biochem. 2016, 80, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Smit, B.A.; Engels, W.J.; Smit, G. Branched chain aldehydes: Production and breakdown pathways and relevance for flavour in foods. Appl. Microbiol. Biotechnol. 2009, 81, 987–999. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhou, W.; Yu, H.; Yuan, J.; Tian, H. Evaluation of the Perceptual Interactions among Aldehydes in a Cheddar Cheese Matrix According to Odor Threshold and Aroma Intensity. Molecules 2020, 25, 4308. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Ushio, H.; Iwanaga, D.; Shinoda, A.; Fujiwara, H.; Kiyohara, R. Taste Enhancing Agent. WO/2011/030650, 16 August 2010. [Google Scholar]
- Kitajima, S.; Nonaka, M. Taste-Improved Food and Drink, and Method for Producing the Same. JP2011172508, 8 September 2011. [Google Scholar]
- Toda, Y.; Nakagita, T.; Hirokawa, T.; Yamashita, Y.; Nakajima, A.; Narukawa, M.; Ishimaru, Y.; Uchida, R.; Misaka, T. Positive/Negative Allosteric Modulation Switching in an Umami Taste Receptor (T1R1/T1R3) by a Natural Flavor Compound, Methional. Sci. Rep. 2018, 8, 11796. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, N.; Kitajima, S.; Tanaka, E.; Tanaka, H.; Nakagawa, N. Salty Taste Reinforcing Agent. JP2009240684, 28 April 2011. [Google Scholar]
- Yamashita, Y.; Nakajima, A.; Uchida, R. Liquid Flavouring Having Enhanced Glutamic Acid Taste. WO/2014/014093, 23 January 2014. [Google Scholar]
- Ueda, Y.; Yonemitsu, M.; Tsubuku, T.; Sakaguchi, M.; Miyajima, R. Flavor characteristics of glutathione in raw and cooked foodstuffs. Biosci. Biotechnol. Biochem. 1997, 61, 1977–1980. [Google Scholar] [CrossRef] [Green Version]
- Ueda, Y.; Tsubuku, T.; Miyajima, R. Composition of Sulfur-Containing Components in Onion and Their Flavor Characters. Biosci. Biotechnol. Biochem. 1994, 58, 108–110. [Google Scholar] [CrossRef]
- Ohsu, T.; Amino, Y.; Nagasaki, H.; Yamanaka, T.; Takeshita, S.; Hatanaka, T.; Maruyama, Y.; Miyamura, N.; Eto, Y. Involvement of the calcium-sensing receptor in human taste perception. J. Biol. Chem. 2010, 285, 1016–1022. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Miyamura, N. Mechanism of the perception of “kokumi” substances and the sensory characteristics of the “kokumi” peptide, γ-Glu-Val-Gly. Flavour 2015, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Dunkel, A.; Koster, J.; Hofmann, T. Molecular and sensory characterization of gamma-glutamyl peptides as key contributors to the kokumi taste of edible beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2007, 55, 6712–6719. [Google Scholar] [CrossRef]
- Miyamura, N.; Jo, S.; Kuroda, M.; Kouda, T. Flavour improvement of reduced-fat peanut butter by addition of a kokumi peptide, γ-glutamyl-valyl-glycine. Flavour 2015, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, S.; Maruyama, Y.; Sasaki, K.; Tajima, T.; Kuroda, M. Increases in the pungency of allyl isothiocyanate and piperine by CaSR agonists, glutathione and gamma-glutamyl-valyl-glycine. Physiol. Behav. 2022, 256, 113952. [Google Scholar] [CrossRef]
- Ueda, Y.; Sakaguchi, M.; Hirayama, K.; Miyajima, R.; Kimizuka, A. Characteristic Flavor Constituents in Water Extract of Garlic. Agric. Biol. Chem. 1990, 54, 163–169. [Google Scholar] [CrossRef]
- Maruyama, Y.; Yasuda, R.; Kuroda, M.; Eto, Y. Kokumi substances, enhancers of basic tastes, induce responses in calcium-sensing receptor expressing taste cells. PLoS ONE 2012, 7, e34489. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M.; Gamba, G.; Riccardi, D.; Lombardi, M.; Butters, R.; Kifor, O.; Sun, A.; Hediger, M.A.; Lytton, J.; Hebert, S.C. Cloning and characterization of an extracellular Ca(2+)-sensing receptor from bovine parathyroid. Nature 1993, 366, 575–580. [Google Scholar] [CrossRef]
- Chattopadhyay, N.; Vassilev, P.M.; Brown, E.M. Calcium-sensing receptor: Roles in and beyond systemic calcium homeostasis. Biol. Chem. 1997, 378, 759–768. [Google Scholar]
- Tian, H.Y.; Zhang, J.; Sun, B.G.; Huang, M.Q.; Li, J.R.; Han, X.X. Preparation of natural isovaleraldehyde by the Maillard reaction. Chin. Chem. Lett. 2007, 18, 1049–1052. [Google Scholar] [CrossRef]
- Paula Dionísio, A.; Molina, G.; Souza de Carvalho, D.; dos Santos, R.; Bicas, J.L.; Pastore, G.M. 11 - Natural flavourings from biotechnology for foods and beverages. In Natural Food Additives, Ingredients and Flavourings; Baines, D., Seal, R., Eds.; Woodhead Publishing: Cambridge, UK, 2012; pp. 231–259. [Google Scholar]
- Brennan, S.C.; Davies, T.S.; Schepelmann, M.; Riccardi, D. Emerging roles of the extracellular calcium-sensing receptor in nutrient sensing: Control of taste modulation and intestinal hormone secretion. Br. J. Nutr. 2014, 111, S16–S22. [Google Scholar] [CrossRef] [Green Version]
- Conigrave, A.D.; Mun, H.C.; Lok, H.C. Aromatic L-amino acids activate the calcium-sensing receptor. J. Nutr. 2007, 137, 1524S–1527S; discussion 1548S. [Google Scholar] [CrossRef] [Green Version]
- Odoemelam, C.S.; Percival, B.; Wallis, H.; Chang, M.W.; Ahmad, Z.; Scholey, D.; Burton, E.; Williams, I.H.; Kamerlin, C.L.; Wilson, P.B. G-Protein coupled receptors: Structure and function in drug discovery. RSC Adv. 2020, 10, 36337–36348. [Google Scholar] [CrossRef]
- Geng, Y.; Mosyak, L.; Kurinov, I.; Zuo, H.; Sturchler, E.; Cheng, T.C.; Subramanyam, P.; Brown, A.P.; Brennan, S.C.; Mun, H.C.; et al. Structural mechanism of ligand activation in human calcium-sensing receptor. Elife 2016, 5. [Google Scholar] [CrossRef]
- Wen, T.; Wang, Z.; Chen, X.; Ren, Y.; Lu, X.; Xing, Y.; Lu, J.; Chang, S.; Zhang, X.; Shen, Y.; et al. Structural basis for activation and allosteric modulation of full-length calcium-sensing receptor. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Ahmad, R.; Dalziel, J.E. G Protein-Coupled Receptors in Taste Physiology and Pharmacology. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Winnig, M.; Bufe, B.; Kratochwil, N.A.; Slack, J.P.; Meyerhof, W. The binding site for neohesperidin dihydrochalcone at the human sweet taste receptor. BMC Struct. Biol. 2007, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M.; Katz, C.; Butters, R.; Kifor, O. Polyarginine, polylysine, and protamine mimic the effects of high extracellular calcium concentrations on dispersed bovine parathyroid cells. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1991, 6, 1217–1225. [Google Scholar] [CrossRef]
- Nakajima, S.; Hira, T.; Hara, H. Calcium-sensing receptor mediates dietary peptide-induced CCK secretion in enteroendocrine STC-1 cells. Mol. Nutr. Food Res. 2012, 56, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S.; Hira, T.; Eto, Y.; Asano, K.; Hara, H. Soybean beta 51-63 peptide stimulates cholecystokinin secretion via a calcium-sensing receptor in enteroendocrine STC-1 cells. Regul. Pept. 2010, 159, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Doroszewicz, J.; Waldegger, P.; Jeck, N.; Seyberth, H.; Waldegger, S. pH dependence of extracellular calcium sensing receptor activity determined by a novel technique. Kidney Int. 2005, 67, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, A.W.; DeSouza, M.; Morgan, M.Y. Plasma 3-methyIbutanal in man and its relationship to hepatic encephalopathy. Clin. Physiol. 1985, 5, 53–62. [Google Scholar] [CrossRef]
- Degenhardt, A.G.; Hofmann, T. Bitter-tasting and kokumi-enhancing molecules in thermally processed avocado (Persea americana Mill.). J. Agric. Food Chem. 2010, 58, 12906–12915. [Google Scholar] [CrossRef]
- Rybczynska, A.; Lehmann, A.; Jurska-Jasko, A.; Boblewski, K.; Orlewska, C.; Foks, H.; Drewnowska, K. Hypertensive effect of calcilytic NPS 2143 administration in rats. J. Endocrinol. 2006, 191, 189–195. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitajima, S.; Maruyama, Y.; Kuroda, M. Volatile Short-Chain Aliphatic Aldehydes Act as Taste Modulators through the Orally Expressed Calcium-Sensing Receptor CaSR. Molecules 2023, 28, 4585. https://doi.org/10.3390/molecules28124585
Kitajima S, Maruyama Y, Kuroda M. Volatile Short-Chain Aliphatic Aldehydes Act as Taste Modulators through the Orally Expressed Calcium-Sensing Receptor CaSR. Molecules. 2023; 28(12):4585. https://doi.org/10.3390/molecules28124585
Chicago/Turabian StyleKitajima, Seiji, Yutaka Maruyama, and Motonaka Kuroda. 2023. "Volatile Short-Chain Aliphatic Aldehydes Act as Taste Modulators through the Orally Expressed Calcium-Sensing Receptor CaSR" Molecules 28, no. 12: 4585. https://doi.org/10.3390/molecules28124585
APA StyleKitajima, S., Maruyama, Y., & Kuroda, M. (2023). Volatile Short-Chain Aliphatic Aldehydes Act as Taste Modulators through the Orally Expressed Calcium-Sensing Receptor CaSR. Molecules, 28(12), 4585. https://doi.org/10.3390/molecules28124585