
The Anti-Inflammatory Effects of Cannabis sativa Extracts on LPS-Induced Cytokines Release in Human Macrophages

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Analysis of Cannabinoids Content Using HPLC
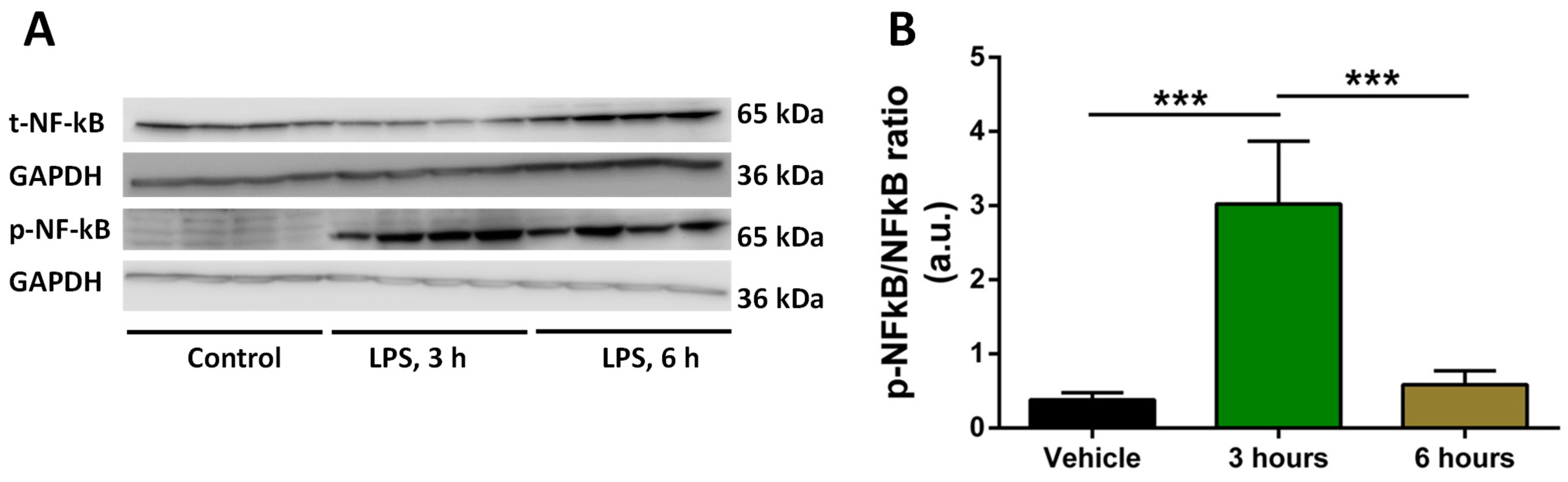
2.2. Analysis of Time of Induction of Inflammation by LPS
2.3. Treatment with Selected Concentrations of LPS, THC, CBD, and Extracts Does Not Reduce the Viability of Macrophages
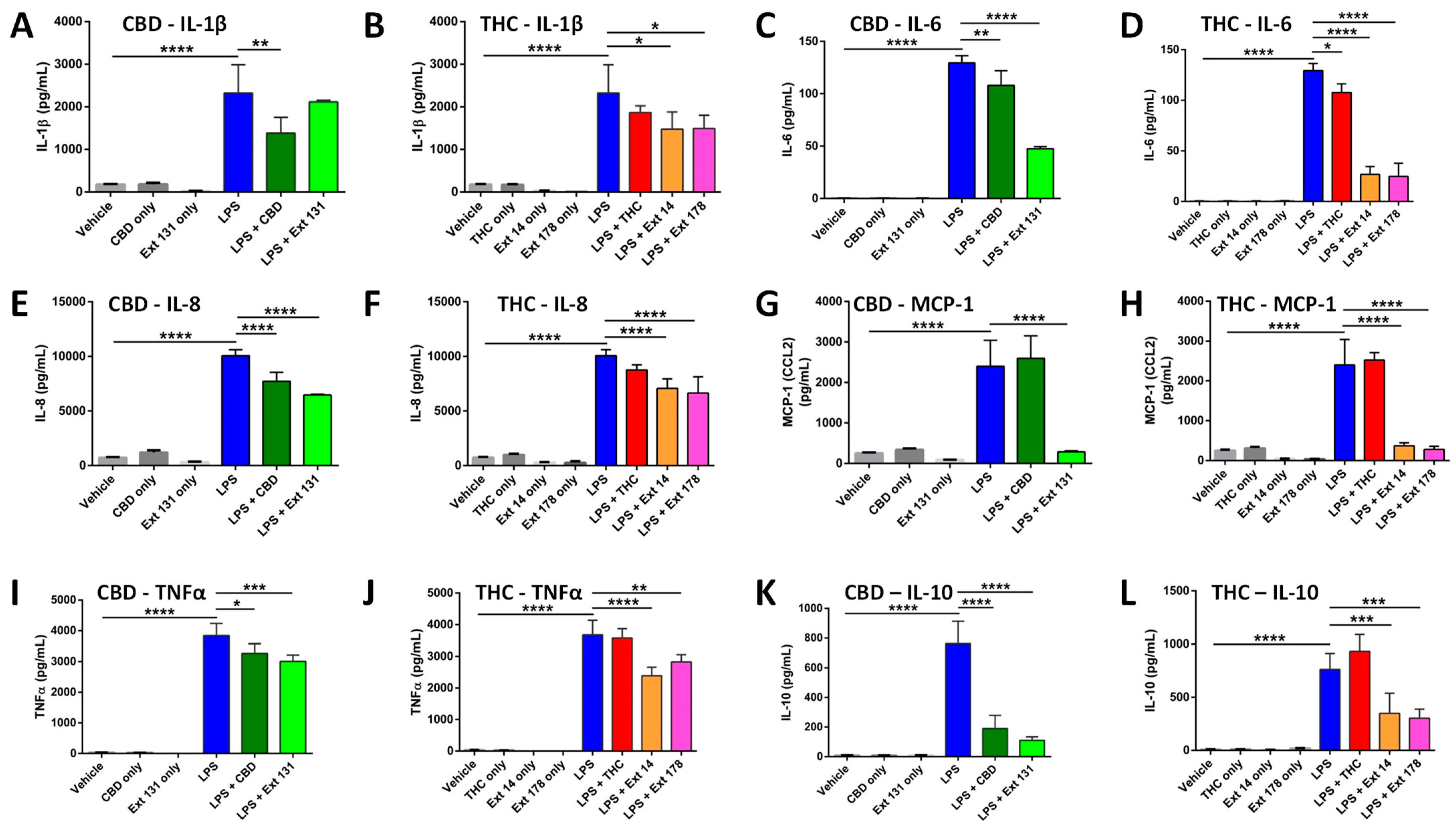
2.4. Cannabis sativa Extracts Attenuate TNF-α, IL-1β, IL-8, MCP-1, IL-10, and IL-6 Production in LPS-Stimulated THP-1 Macrophages
3. Materials and Methods
3.1. Main Reagents
3.2. Plant Growth and Extract Preparation
3.3. High Performance Liquid Chromatography (HPLC) Analysis of Cannabinoids
3.4. Terpene Analysis
3.5. Cell cultures and Treatments
3.6. Cell Viability Assay Using Trypan Blue
3.7. Multiplex ELISA
3.8. Statistical Analysis
4. Discussion
5. Conclusions and Limitations of This Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cohen, K.; Weizman, A.; Weinstein, A. Positive and Negative Effects of Cannabis and Cannabinoids on Health. Clin. Pharmacol. Ther. 2019, 105, 1139–1147. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Russo, E.B. The case for the entourage effect and conventional breeding of clinical cannabis: No ‘Strain,’ no gain. Front. Plant Sci. 2019, 9, 1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiurchiù, V. Endocannabinoids and Immunity. Cannabis Cannabinoid Res. 2016, 1, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galiègue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carrière, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Foxman, E.F.; Molony, R.D. Early local immune defences in the respiratory tract. Nat. Rev. Immunol. 2017, 17, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.C.; Weinlich, R.; Bortoluci, K.R. Pattern recognition receptors and the host cell death molecular machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the eye of the cytokine storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–325. [Google Scholar] [CrossRef]
- Benakanakere, M.R.; Finoti, L.S.; Tanaka, U.; Grant, G.R.; Scarel-Caminaga, R.M.; Kinane, D.F. Investigation of the functional role of human Interleukin-8 gene haplotypes by CRISPR/Cas9 mediated genome editing. Sci. Rep. 2016, 6, 31180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelová, H.; Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Martin-Sanchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.I.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.J.D.; Mortellaro, A.; et al. Inflammasome-dependent IL-1β release depends upon membrane permeabilisation. Cell Death Differ. 2016, 23, 1219–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Il-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, 16295–16296. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Nagarkatti, P.; Pandey, R.; Rieder, S.A.; Hegde, V.L.; Nagarkatti, M. Cannabinoids as novel anti-inflammatory drugs. Future Med. Chem. 2009, 1, 1333–1349. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, W.; Elgendy, N.; Salama, S.; Jawad, M.; Eltoukhy, K. The Effect of Cannabis on the Clinical and Cytokine Profiles in Patients with Multiple Sclerosis. Mult. Scler. Int. 2021, 2021, 6611897. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Wang, B.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, S.; Kovalchuk, I.; Kovalchuk, O. Fighting the Storm: Could Novel Anti-Tnfá And Anti-Il-6 C. Sativa Cultivars Tame Cytokine Storm in COVID-19? Aging 2021, 13, 1571–1590. [Google Scholar] [CrossRef]
- Gallily, R.; Yekhtin, Z.; Ondřej Hanuš, L. Overcoming the Bell-Shaped Dose-Response of Cannabidiol by Using Cannabis Extract Enriched in Cannabidiol. Pharmacol. Pharm. 2015, 6, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G.; Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci. Rep. 2021, 11, 1462. [Google Scholar] [CrossRef]
- Khuja, I.; Yekhtin, Z.; Or, R.; Almogi-Hazan, O. Cannabinoids Reduce Inflammation but Inhibit Lymphocyte Recovery in Murine Models of Bone Marrow Transplantation. Int. J. Mol. Sci. Artic. 2019, 20, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, D.; Fiselier, A.; Kovalchuk, I.; Kovalchuk, O. New AKT-dependent mechanisms of anti-COVID-19 action of high-CBD Cannabis sativa extracts. Cell Death Discov. 2022, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kovalchuk, A.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, Y.; Kovalchuk, I.; Kovalchuk, O. In search of preventive strategies: Novel high-CBD Cannabis sativa extracts modulate ACE2 expression in COVID-19 gateway tissues. Aging 2020, 12, 22425–22444. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, S.V.; Zaiachuk, M.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. Cannabinoids Alleviate the LPS-Induced Cytokine Storm via Attenuating NLRP3 Inflammasome Signaling and TYK2-Mediated STAT3 Signaling Pathways In Vitro. Cells 2022, 11, 1391. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Gatto, F.; Cagliani, R.; Catelani, T.; Guarnieri, D.; Moglianetti, M.; Pompa, P.P.; Bardi, G. PMA-induced THP-1 macrophage differentiation is not impaired by citrate-coated platinum nanoparticles. Nanomaterials 2017, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yin, S.; Chen, Y.; Wu, Y.; Zheng, W.; Dong, H.; Bai, Y.; Qin, Y.; Li, J.; Feng, S.; et al. LPS-induced proinflammatory cytokine expression in human airway epithelial cells and macrophages via NF-κB, STAT3 or AP-1 activation. Mol. Med. Rep. 2018, 17, 5484–5491. [Google Scholar] [CrossRef] [Green Version]
- Juknat, A.; Gao, F.; Coppola, G.; Vogel, Z.; Kozela, E. miRNA expression profiles and molecular networks in resting and LPS-activated BV-2 microglia—Effect of cannabinoids. PLoS ONE 2019, 14, e0212039. [Google Scholar] [CrossRef]
- Andrade, C. The P value and statistical significance: Misunderstandings, explanations, challenges, and alternatives. Indian J. Psycholog. Med. 2019, 41, 210–215. [Google Scholar] [CrossRef]
- Harikrishnan, H.; Jantan, I.; Haque, M.A.; Kumolosasi, E. Anti-inflammatory effects of Phyllanthus amarus Schum. & Thonn. through inhibition of NF-κB, MAPK, and PI3K-Akt signaling pathways in LPS-induced human macrophages. BMC Complement. Altern. Med. 2018, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Kong, Q.; Six, D.A.; Liu, Q.; Gu, L.; Wang, S.; Alamuri, P.; Raetz, C.R.; Curtiss, R., 3rd. Phosphate groups of lipid a are essential for Salmonella enterica serovar typhimurium virulence and affect innate and adaptive immunity. Infect. Immun. 2012, 80, 3215–3224. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.; Yang, L.; Dai, F.; Tang, L.; Le, Y. Macrophage differentiation induced by PMA is mediated by activation of RhoA/ROCK signaling. J. Toxicol. Sci. 2017, 42, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.F.; Garcia, J.; Wu, D.; Mitchell, D.C.; Zhang, Y.; Kado, N.Y.; Wong, P.; Trujillo, D.A.; Lollies, A.; Bennet, D.; et al. Activation of inflammatory responses in human U937 macrophages by particulate matter collected from dairy farms: An in vitro expression analysis of pro-inflammatory markers. Environ. Health 2012, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional profiling of the LPS induced NF-κB response in macrophages. BMC Immunol. 2007, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neustock, P.; Brand, J.M.; Kruse, A.; Kirchner, H. Cytokine production of the human monocytic cell line Mono Mac 6 in comparison to mature monocytes in peripheral blood mononuclear cells. Immunobiology 1993, 188, 293–302. [Google Scholar] [CrossRef]
- Hume Underhill, D.A.; Wainwright, B.J.; Aderem, A.; Timothy Ravasi, S.; Wells, C.; Forest, A. Probability of Individual Inducible Genes from Gene-Autonomous Transcriptional System: Macrophage Heterogeneity Arises Generation of Diversity in the Innate Immune. J. Immunol. Ref. 2021, 168, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Kovalchuk, O.; Kovalchuk, I. Cannabinoids as anticancer therapeutic agents. Cell Cycle 2020, 19, 961–989. [Google Scholar] [CrossRef]
- Ferber, S.G.; Namdar, D.; Hen-Shoval, D.; Eger, G.; Koltai, H.; Shoval, G.; Shbiro, L.; Weller, A. The ‘Entourage Effect’: Terpenes Coupled with Cannabinoids for the Treatment of Mood Disorders and Anxiety Disorders. Curr. Neuropharmacol. 2019, 18, 87–96. [Google Scholar] [CrossRef]
- Guo, H.; Callaway, J.B.; Ting, J.P. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat Med 2015, 21, 677–687. [Google Scholar] [CrossRef] [Green Version]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8, S3. [Google Scholar] [CrossRef] [Green Version]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat Rev Immunol 2023, 23, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Viedt, C.; Dechend, R.; Fei, J.; Hänsch, G.M.; Kreuzer, J.; Orth, S.R. MCP-1 Induces Inflammatory Activation of Human Tubular Epithelial Cells: Involvement of the Transcription Factors, Nuclear Factor-B and Activating Protein-1. J. Am. Soc. Nephrol. 2002, 13, 1534–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katona, S.; Kaminski, E.; Sanders, H.; Zajicek, J. Cannabinoid influence on cytokine profile in multiple sclerosis. Clin. Exp. Immunol. 2005, 140, 580–585. [Google Scholar] [CrossRef]
- Aswad, M.; Hamza, H.; Pechkovsky, A.; Zikrach, A.; Popov, T.; Zohar, Y.; Shahar, E.; Louria-Hayon, I. High-CBD Extract (CBD-X) Downregulates Cytokine Storm Systemically and Locally in Inflamed Lungs. Front Immunol. 2022, 13, 875546. [Google Scholar] [CrossRef]
- Muthumalage, T.; Rahman, I. Cannabidiol differentially regulates basal and LPS-induced inflammatory responses in macrophages, lung epithelial cells, and fibroblasts. Toxicol. Appl. Pharmacol. 2019, 382, 114713. [Google Scholar] [CrossRef]
- Yeisley, D.J.; Arabiyat, A.S.; Hahn, M.S. Cannabidiol-Driven Alterations to Inflammatory Protein Landscape of Lipopolysaccharide-Activated Macrophages In Vitro May Be Mediated by Autophagy and Oxidative Stress. Cannabis Cannabinoid Res. 2021, 6, 253–263. [Google Scholar] [CrossRef]
- Comalada, M.; Ballester, I.; Bailón, E.; Sierra, S.; Xaus, J.; Gálvez, J.; de Medina, F.S.; Zarzuelo, A. Inhibition of pro-inflammatory markers in primary bone marrow-derived mouse macrophages by naturally occurring flavonoids: Analysis of the structure-activity relationship. Biochem. Pharmacol. 2006, 72, 1010–1021. [Google Scholar] [CrossRef]
- Joffre, J.; Yeh, C.C.; Wong, E.; Thete, M.; Xu, F.; Zlatanova, I.; Lloyd, E.; Kobzik, L.; Legrand, M.; Hellman, J. Activation of CB 1 R Promotes Lipopolysaccharide-Induced IL-10 Secretion by Monocytic Myeloid-Derived Suppressive Cells and Reduces Acute Inflammation and Organ Injury. J. Immunol. 2020, 204, 3339–3350. [Google Scholar] [CrossRef]
- Kozela, E.; Juknat, A.; Health, M.; Kaushansky, N.; Rimmerman, N. Cannabinoids Decrease the Th17 Inflammatory Autoimmune Phenotype Mechanisms of Anti-depressive Treatments View project Cell death induced by delta-aminolevulinic acid in astrocytes View project. Artic. J. Neuroimmune Pharmacol. 2013, 8, 1265–1276. [Google Scholar] [CrossRef]
- Weiss, L.; Zeira, M.; Reich, S.; Har-Noy, M.; Mechoulam, R.; Slavin, S.; Gallily, R. Cannabidiol lowers incidence of diabetes in non-obese diabetic mice. Autoimmunity 2006, 39, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Total THC, % | Total CBD, % | THC (μM) | CBD (μM) | |
|---|---|---|---|---|
| #14 | 33.35 ± 2.75 | 2.81 ± 0.23 | 7.42 ± 0.56 | 0.63 ± 0.04 |
| #131 | 2.11 ± 0.19 | 19.65 ± 2.05 | 0.47 ± 0.03 | 4.37± 0.33 |
| #178 | 33.98 ± 3.12 | 1.01 ± 0.09 | 7.56 ± 0.64 | 0.22 ± 0.02 |
| Terpenes in mg/g | #131 | #178 | #14 |
|---|---|---|---|
| α-Pinene | 0.295 ± 0.055 | 0.649 ± 0.12 | 0.048 ± 0.008 |
| β-Pinene | 0.212 ± 0.046 | 0.245 ± 0.042 | 0.068 ± 0.012 |
| β-Myrcene | ND | 0.361 ± 0.064 | 0.124 ± 0.042 |
| Limonene | 0.262 ± 0.052 | 0.003 ± 0.001 | 0.263 ± 0.062 |
| Terpinolene | 0.025 ± 0.005 | 0.008 ± 0.002 | 0.004 ± 0.001 |
| Linalool | 0.058 ± 0.009 | 0.029 ± 0.005 | 0.193 ± 0.062 |
| α-Bisabolol | 0.003 ± 0.001 | 0.244 ± 0.06 | 0.061 ± 0.02 |
| trans-Caryophyllene | 0.04 ± 0.008 | 0.076 ± 0.014 | 0.545 ± 0.08 |
| α-Humulene | ND | 0.04 ± 0.009 | 0.136 ± 0.03 |
| trans-Nerolidol | ND | 0.008 ± 0.002 | 0.187 ± 0.04 |
| cis-Nerolidol | 0.001 ± 0.001 | 0.003 ± 0.001 | ND |
| Camphene | 0.022 ± 0.006 | 0.025 ± 0.001 | 0.015 ± 0.005 |
| β-Ocimene | ND | 0.089 ± 0.016 | ND |
| Fenchone isomers | ND | 0.003 ± 0.001 | 0.004 ± 0.001 |
| δ-3-Carene | 0.506 ± 0.12 | 0.001 ± 0.001 | ND |
| α-Terpinene | 0.006 ± 0.002 | 0.573 ± 0.11 | ND |
| Eucalyptol | 0.002 ± 0.001 | ND | ND |
| γ-Terpinene | ND | 0.001 ± 0.001 | ND |
| β-Cymene | ND | 0.048 ± 0.009 | ND |
| Camphor isomers | ND | 0.118 ± 0.04 | ND |
| Isopulegol | 0.01 ± 0.004 | 0.016 ± 0.004 | ND |
| Caryophyllene oxide | 0.017 ± 0.005 | 0.098 ± 0.003 | ND |
| Valencene | ND | 0.01 ± 0.003 | ND |
| Geraniol | 0.005 ± 0.002 | 0.004 ± 0.002 | ND |
| Guaiol | 0.029 ± 0.005 | 0.116 ± 0.04 | ND |
| trans-P-Ocimene | 0.016 ± 0.004 | ND | ND |
| α-Humulene | 0.025 ± 0.006 | ND | 0.136 ± 0.034 |
| Fenchyl Alcohol | ND | ND | 0.036 ± 0.007 |
| Borneol isomers | ND | ND | 0.012 ± 0.003 |
| α-Terpineol | ND | ND | 0.052 ± 0.009 |
| Total Terpenes | 1.534 ± 0.32 | 2.768 ± 0.46 | 1.75 ± 0.25 |
| Analyzed Cytokines | CBD | THC | #131 | #14 | #178 |
|---|---|---|---|---|---|
| IL-1β | ↓ | = | = | ↓ | ↓ |
| IL-6 | ↓ | ↓ | ↓ | ↓ | ↓ |
| IL-8 | ↓ | = | ↓ | ↓ | ↓ |
| IL-10 | ↓ | = | ↓ | ↓ | ↓ |
| MCP-1 | = | = | ↓ | ↓ | ↓ |
| TNF-α | ↓ | = | ↓ | ↓ | ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaiachuk, M.; Suryavanshi, S.V.; Pryimak, N.; Kovalchuk, I.; Kovalchuk, O. The Anti-Inflammatory Effects of Cannabis sativa Extracts on LPS-Induced Cytokines Release in Human Macrophages. Molecules 2023, 28, 4991. https://doi.org/10.3390/molecules28134991
Zaiachuk M, Suryavanshi SV, Pryimak N, Kovalchuk I, Kovalchuk O. The Anti-Inflammatory Effects of Cannabis sativa Extracts on LPS-Induced Cytokines Release in Human Macrophages. Molecules. 2023; 28(13):4991. https://doi.org/10.3390/molecules28134991
Chicago/Turabian StyleZaiachuk, Mariia, Santosh V. Suryavanshi, Nazar Pryimak, Igor Kovalchuk, and Olga Kovalchuk. 2023. "The Anti-Inflammatory Effects of Cannabis sativa Extracts on LPS-Induced Cytokines Release in Human Macrophages" Molecules 28, no. 13: 4991. https://doi.org/10.3390/molecules28134991
APA StyleZaiachuk, M., Suryavanshi, S. V., Pryimak, N., Kovalchuk, I., & Kovalchuk, O. (2023). The Anti-Inflammatory Effects of Cannabis sativa Extracts on LPS-Induced Cytokines Release in Human Macrophages. Molecules, 28(13), 4991. https://doi.org/10.3390/molecules28134991