
Effects of Lidocaine-Derived Organic Compounds on Eosinophil Activation and Survival

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Pharmacophore-Based Design of Lidocaine Modified Analogs
2.2. Effects of the Lidocaine Analogs on the Survival of Eosinophils
2.3. Effects of the Lidocaine Analogs on Superoxide Production from Eosinophils
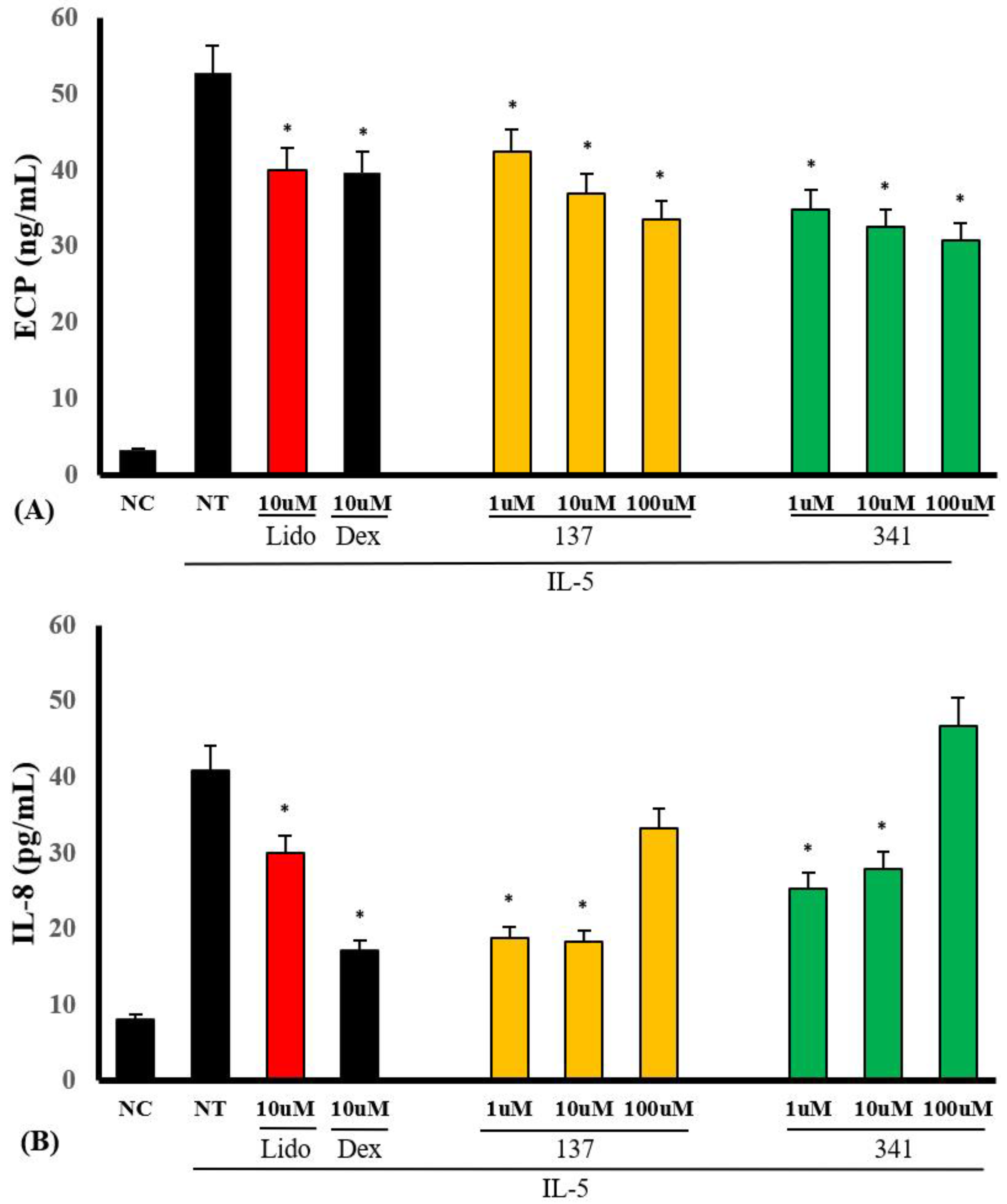
2.4. Effects of the Synthesized Compounds on Eosinophil Cationic Protein (ECP) and IL-8 Production from Eosinophils
2.5. Effects of the Synthesized Compounds on Transcription Factor Expression in Eosinophils
2.6. Effects of the Synthesized Compounds on the Migration of Eosinophils by Platelet-Activating Factor (PAF)
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Building a Pharmacophore Model
4.3. Isolation of Eosinophils
4.4. Cytotoxic Effect of Lidocaine-Derived Compounds on Eosinophils
4.5. Survival Analysis on Eosinophils
4.6. Superoxide Anion Production from Eosinophils
4.7. Production of ECP and IL-8 from Eosinophils
4.8. Expression of Transcription Factors from Eosinophils
4.9. Migration Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Jacobsen, E.A.; Taranova, A.G.; Lee, N.A.; Lee, J.J. Eosinophils: Singularly destructive effector cells or purveyors of immunoregulation? J. Allergy Clin. Immunol. 2007, 119, 1313–1320. [Google Scholar] [CrossRef]
- Kita, H. Eosinophils: Multifaceted biological properties and roles in health and disease. Immunol. Rev. 2011, 242, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Phipps, S.; Foster, P.S. Eosinophil trafficking in allergy and asthma. J. Allergy Clin. Immunol. 2007, 119, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Akdis, M.; Blaser, K.; Akdis, C.A. T regulatory cells in allergy: Novel concepts in the pathogenesis, prevention, and treatment of allergic diseases. J. Allergy Clin. Immunol. 2005, 116, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Daraz, Y.M.; Abdelghffar, O.H. Lidocaine Infusion: An Antiarrhythmic with Neurologic Toxicities. Cureus 2022, 14, e23310. [Google Scholar] [CrossRef]
- Muraki, M.; Iwanaga, T.; Haraguchi, R.; Kubo, H.; Tohda, Y. Continued inhalation of lidocaine suppresses antigen-induced airway hyperreactivity and airway inflammation in ovalbumin-sensitized guinea pigs. Int. Immunopharmacol. 2008, 8, 725–731. [Google Scholar] [CrossRef]
- Hunt, L.W.; Frigas, E.; Butterfield, J.H.; Kita, H.; Blomgren, J.; Dunnette, S.L.; Offord, K.P.; Gleich, G.J. Treatment of asthma with nebulized lidocaine: A randomized, placebo-controlled study. J. Allergy Clin. Immunol. 2004, 113, 853–859. [Google Scholar] [CrossRef]
- Serra, M.F.; Anjos-Valotta, E.A.; Olsen, P.C.; Couto, G.C.; Jurgilas, P.B.; Cotias, A.C.; Pao, C.R.; Ferreira, T.P.; Arantes, A.C.; Pires, A.L.; et al. Nebulized lidocaine prevents airway inflammation, peribronchial fibrosis, and mucus production in a murine model of asthma. Anesthesiology 2012, 117, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Olsen, P.C.; Coelho, L.P.; da Costa, J.C.; Cordeiro, R.S.; Silva, P.M.; Martins, M.A. Two for one: Cyclic AMP mediates the anti-inflammatory and anti-spasmodic properties of the non-anesthetic lidocaine analog JMF2-1. Eur. J. Pharmacol. 2012, 680, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Serra, M.F.; Neves, J.S.; Couto, G.C.; Cotias, A.C.; Pao, C.R.; Olsen, P.C.; de Carvalho, K.I.; Anjos-Valotta, E.A.; Faria, R.X.; Costa, J.C.; et al. JM25-1, a Lidocaine Analog Combining Airway Relaxant and Antiinflammatory Properties: Implications for New Bronchospasm Therapy. Anesthesiology 2016, 124, 109–120. [Google Scholar] [CrossRef]
- Grande, F.; Yamada, R.; Cao, X.; Aiello, F.; Garofalo, A.; Neamati, N. Synthesis and biological evaluation of novel hydrazide based cytotoxic agents. Expert. Opin. Investig. Drugs 2009, 18, 555–568. [Google Scholar] [CrossRef]
- Plasencia, C.; Grande, F.; Oshima, T.; Cao, X.; Yamada, R.; Sanchez, T.; Aiello, F.; Garofalo, A. Discovery of a novel quinoxalinhydrazide with a broad-spectrum anticancer activity. Cancer Biol. Ther. 2009, 8, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Kempe-Dustin, J.J.; Aboul-Fadl, T.; Christensen, C.; Palais, R.; Parsawar, K.; Gleich, G.J.; Wagner, L.A. Cell screening assay for identifying inhibitors of eosinophil proliferation. Drug Dev. Res. 2011, 72, 353–360. [Google Scholar] [CrossRef]
- Wong, C.K.; Ng, S.S.; Lun, S.W.; Cao, J.; Lam, C.W. Signalling mechanisms regulating the activation of human eosinophils by mast-cell-derived chymase: Implications for mast cell-eosinophil interaction in allergic inflammation. Immunology 2009, 126, 579–587. [Google Scholar] [CrossRef]
- Zinchuk, O.; Fukushima, A.; Zinchuk, V.; Fukata, K.; Ueno, H. Direct action of platelet activating factor (PAF) induces eosinophil accumulation and enhances expression of PAF receptors in conjunctivitis. Mol. Vis. 2005, 11, 114–123. [Google Scholar]
- Muraki, M.; Kita, H.; Gleich, G.J. Dexamethasone and lidocaine suppress eosinophilopoiesis from umbilical cord blood cells. Clin. Mol. Allergy 2020, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yan, W.; He, N. Lidocaine attenuates hypoxia/reoxygenation-induced inflammation, apoptosis and ferroptosis in lung epithelial cells by regulating the p38 MAPK pathway. Mol. Med. Rep. 2022, 25, 12666. [Google Scholar] [CrossRef]
- Olsen, P.C.; Ferreira, T.P.; Serra, M.F.; Farias-Filho, F.A.; Fonseca, B.P.; Viola, J.P.; Cordeiro, R.S.; Silva, P.M.; Costa, J.C.; Martins, M.A. Lidocaine-derivative JMF2-1 prevents ovalbumin-induced airway inflammation by regulating the function and survival of T cells. Clin. Exp. Allergy 2011, 41, 250–259. [Google Scholar] [CrossRef]
- Okada, S.; Hagan, J.B.; Kato, M.; Bankers-Fulbright, J.L.; Hunt, L.W.; Gleich, G.J.; Kita, H. Lidocaine and its analogues inhibit IL-5-mediated survival and activation of human eosinophils. J. Immunol. 1998, 160, 4010–4017. [Google Scholar] [CrossRef]
- Han, S.T.; Mosher, D.F. IL-5 induces suspended eosinophils to undergo unique global reorganization associated with priming. Am. J. Respir. Cell Mol. Biol. 2014, 50, 654–664. [Google Scholar] [CrossRef] [Green Version]
- Kouro, T.; Takatsu, K. IL-5- and eosinophil-mediated inflammation: From discovery to therapy. Int. Immunol. 2009, 21, 1303–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelaia, C.; Paoletti, G.; Puggioni, F.; Racca, F.; Pelaia, G.; Canonica, G.W.; Heffler, E. Interleukin-5 in the Pathophysiology of Severe Asthma. Front. Physiol. 2019, 10, 1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Kimura, H.; Kurabayashi, M.; Kozawa, K.; Kato, M. Interferon-gamma enhances human eosinophil effector functions induced by granulocyte-macrophage colony-stimulating factor or interleukin-5. Immunol. Lett. 2008, 118, 88–95. [Google Scholar] [CrossRef]
- El-Hashim, A.Z.; Khajah, M.A.; Orabi, K.Y.; Balakrishnan, S.; Sary, H.G.; Abdelali, A.A. Onion bulb extract downregulates EGFR/ERK1/2/AKT signaling pathway and synergizes with steroids to inhibit allergic inflammation. Front. Pharmacol. 2020, 2, 551683. [Google Scholar] [CrossRef]
- Lee, Y.A.; Shin, M.H. Mitochondrial respiration is required for activation of ERK1/2 and caspase-3 in human eosinophils stimulated with hydrogen peroxide. J. Investig. Allergol. Clin. Immunol. 2009, 19, 188–194. [Google Scholar]
- da Costa, J.C.; Olsen, P.C.; de Azeredo Siqueira, R.; de Frias Carvalho, V.; Serra, M.F.; Alves, L.A.; Faria, R.X.; Xisto, D.G.; Rocco, P.R.; Cordeiro, R.S.; et al. JMF2-1, a lidocaine derivative acting on airways spasm and lung allergic inflammation in rats. J. Allergy Clin. Immunol. 2007, 119, 219–225. [Google Scholar] [CrossRef]
- Kownatzki, E.; Uhrich, S. Enhancement of chemotactic migration by the local anesthetic tetracaine. Immunobiology 1983, 165, 97–106. [Google Scholar] [CrossRef]
- Kim, H.S.; Won, S.; Lee, E.K.; Chun, Y.H.; Yoon, J.S.; Kim, J.T.; Kim, H.H. Effect of proparacaine in a mouse model of allergic rhinitis. Clin. Exp. Otorhinolaryngol. 2017, 10, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Wardlaw, A.J.; Moqbel, R.; Cromwell, O.; Kay, A.B. Platelet-activating Factor. A Potent Chemotactic and Chemokinetic Factor for Human Eosinophils. J. Clin. Investig. 1986, 78, 1701–1706. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-H.; Ye, M.-K.; Chae, M.-H.; Geum, S.-Y.; Aboraia, A.S.; Abdel-Aal, A.-B.M.; Qayed, W.S.; Abd El-wahab, H.A.A.; Abou-Ghadir, O.F.; Aboul-Fadl, T. Effects of Lidocaine-Derived Organic Compounds on Eosinophil Activation and Survival. Molecules 2023, 28, 5696. https://doi.org/10.3390/molecules28155696
Shin S-H, Ye M-K, Chae M-H, Geum S-Y, Aboraia AS, Abdel-Aal A-BM, Qayed WS, Abd El-wahab HAA, Abou-Ghadir OF, Aboul-Fadl T. Effects of Lidocaine-Derived Organic Compounds on Eosinophil Activation and Survival. Molecules. 2023; 28(15):5696. https://doi.org/10.3390/molecules28155696
Chicago/Turabian StyleShin, Seung-Heon, Mi-Kyung Ye, Mi-Hyun Chae, Sang-Yen Geum, Ahmed S. Aboraia, Abu-Baker M. Abdel-Aal, Wesam S. Qayed, Hend A. A. Abd El-wahab, Ola F. Abou-Ghadir, and Tarek Aboul-Fadl. 2023. "Effects of Lidocaine-Derived Organic Compounds on Eosinophil Activation and Survival" Molecules 28, no. 15: 5696. https://doi.org/10.3390/molecules28155696