


Unraveling the Pivotal Roles of Various Metal Ion Centers in the Catalysis of Quercetin 2,4-Dioxygenases


Abstract
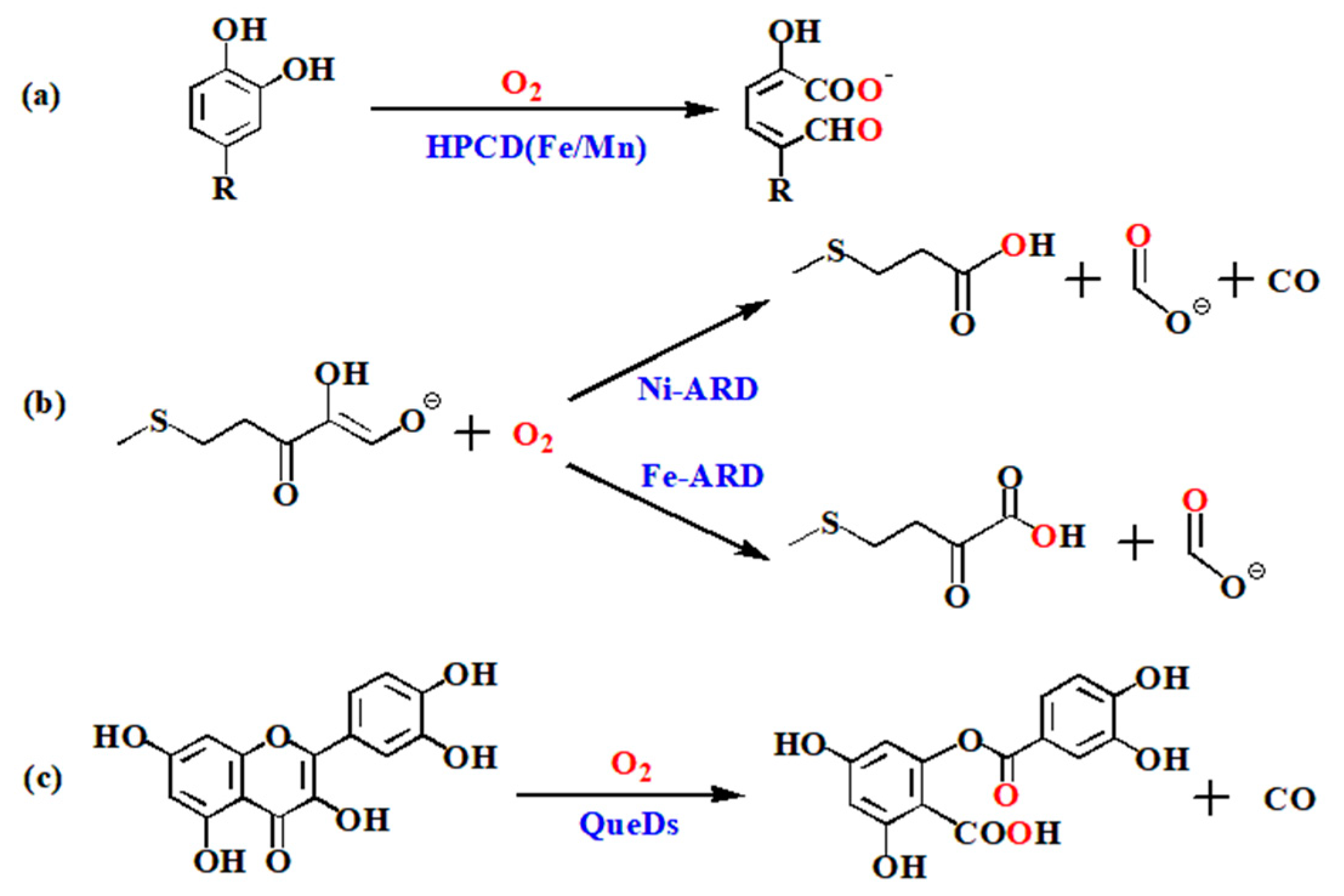
:1. Introduction
2. Results and Discussion
2.1. Influencing Factors of Dioxygen Activation
2.2. Electronic and Structural Characteristics of Metal-Containing QueDs Initial Reactants
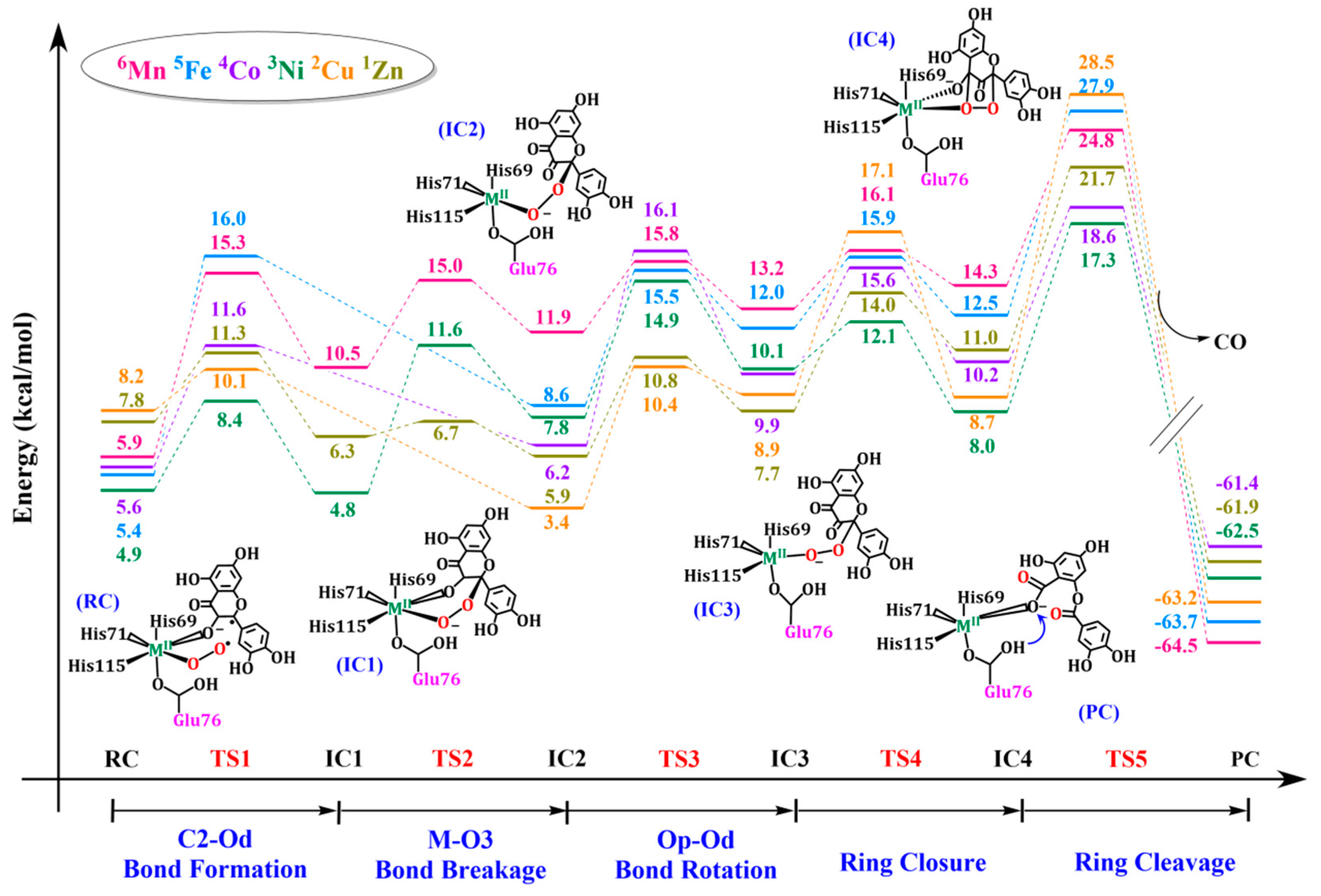
2.3. Comparison of Reaction Mechanism of QueDs Containing Different Metal Ions
3. Computational Details
3.1. System Preparation
3.2. QM/MM Calculations
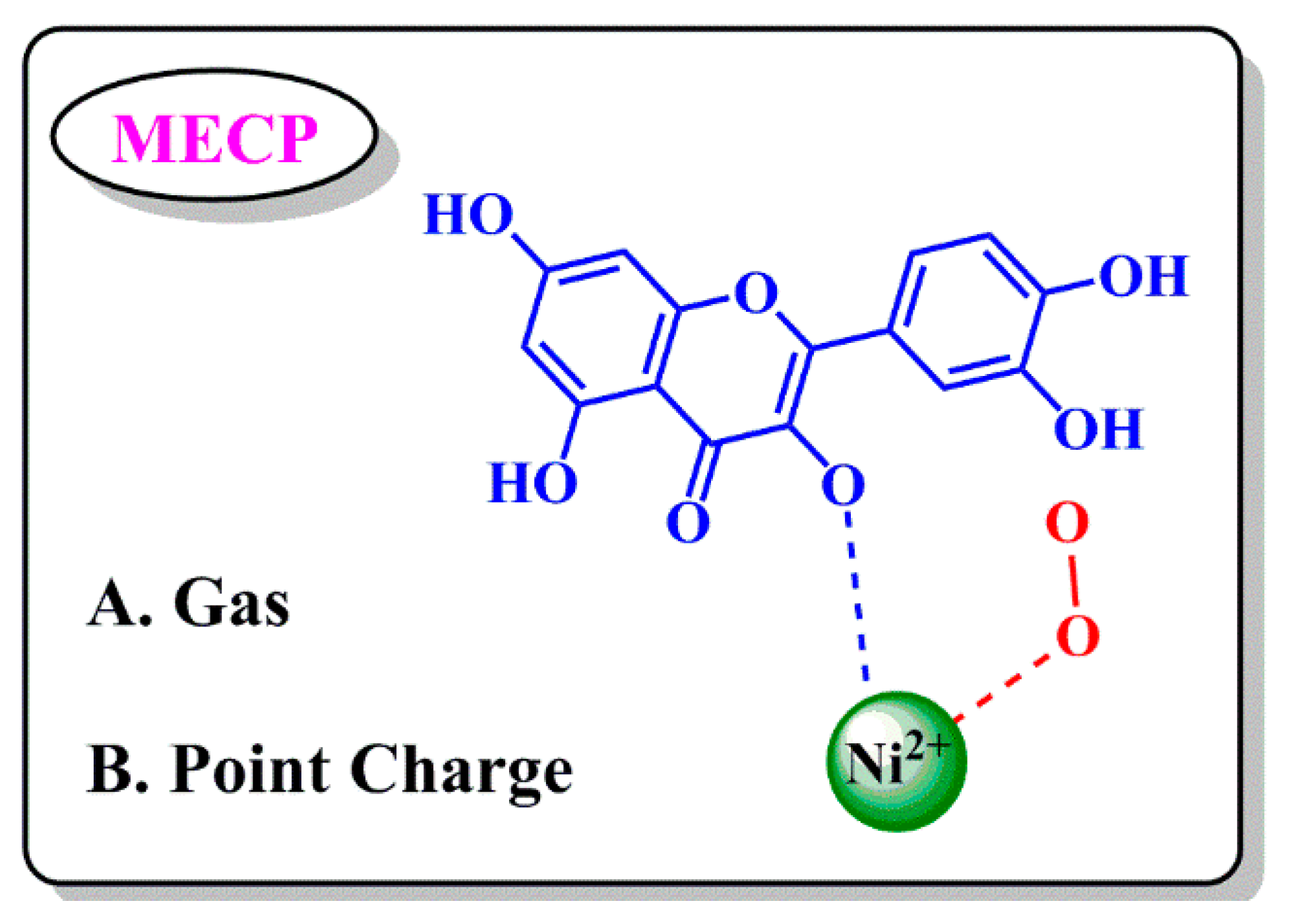
3.3. Calculation of MECP
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Sample Availability
References
- Vallee, B.L.; Williams, R.J. Metalloenzymes: The entatic nature of their active sites. Proc. Natl. Acad. Sci. USA 1968, 59, 498–505. [Google Scholar] [CrossRef]
- Siegbahn, P.E.M. Mechanisms of metalloenzymes studied by quantum chemical methods. Q. Rev. Biophys. 2003, 36, 91–145. [Google Scholar] [CrossRef]
- Nam, W. Dioxygen Activation by Metalloenzymes and Models. Acc. Chem. Res. 2007, 40, 465. [Google Scholar] [CrossRef]
- Sigel, H.; McCormick, D.B. Discriminating behavior of metal ions and ligands with regard to their biological significance. Acc. Chem. Res. 1970, 3, 201–208. [Google Scholar] [CrossRef]
- Sigel, R.K.O.; Pyle, A.M. Alternative Roles for Metal Ions in Enzyme Catalysis and the Implications for Ribozyme Chemistry. Chem. Rev. 2007, 107, 97–113. [Google Scholar] [CrossRef]
- Georgiev, V.; Borowski, T.; Blomberg, M.R.; Siegbahn, P.E. A comparison of the reaction mechanisms of iron- and manganese-containing 2,3-HPCD: An important spin transition for manganese. J. Biol. Inorg. Chem. 2008, 13, 929–940. [Google Scholar] [CrossRef]
- Christian, G.J.; Ye, S.; Neese, F. Oxygen activation in extradiol catecholate dioxygenases—A density functional study. Chem. Sci. 2012, 3, 1600–1611. [Google Scholar] [CrossRef]
- Dong, G.; Lai, W. Reaction mechanism of homoprotocatechuate 2,3-dioxygenase with 4-nitrocatechol: Implications for the role of substrate. J. Phys. Chem. B 2014, 118, 1791–1798. [Google Scholar] [CrossRef]
- Allpress, C.J.; Grubel, K.; Szajna-Fuller, E.; Arif, A.M.; Berreau, L.M. Regioselective aliphatic carbon–carbon bond cleavage by a model system of relevance to iron-containing acireductone dioxygenase. J. Am. Chem. Soc. 2013, 135, 659–668. [Google Scholar] [CrossRef]
- Sparta, M.; Valdez, C.E.; Alexandrova, A.N. Metal-dependent activity of Fe and Ni acireductone dioxygenases: How two electrons reroute the catalytic pathway. J. Mol. Biol. 2013, 425, 3007–3018. [Google Scholar] [CrossRef]
- Deshpande, A.R.; Wagenpfeil, K.; Pochapsky, T.C.; Petsko, G.A.; Ringe, D. Metal-Dependent Function of a Mammalian Acireductone Dioxygenase. Biochemistry 2016, 55, 1398–1407. [Google Scholar] [CrossRef]
- Miłaczewska, A.; Kot, E.; Amaya, J.A.; Makris, T.M.; Zając, M.; Korecki, J.; Chumakov, A.; Trzewik, B.; Kędracka-Krok, S.; Minor, W.; et al. On the Structure and Reaction Mechanism of Human Acireductone Dioxygenase. Chem. Eur. J. 2018, 24, 5225–5237. [Google Scholar] [CrossRef]
- Sun, Y.J.; Huang, Q.Q.; Tano, T.; Itoh, S. Flavonolate complexes of M(II) (M = Mn, Fe, Co, Ni, Cu, and Zn). Structural and functional models for the ES (enzyme-substrate) complex of quercetin 2,3-dioxygenase. Inorg. Chem. 2013, 52, 10936–10948. [Google Scholar] [CrossRef]
- Jeoung, J.H.; Nianios, D.; Fetzner, S.; Dobbek, H. Quercetin 2,4-Dioxygenase Activates Dioxygen in a Side-On O2-Ni Complex. Angew. Chem. Int. Ed. 2016, 55, 3281–3284. [Google Scholar] [CrossRef]
- Eom, H.; Cao, Y.; Kim, H.; de Visser, S.P.; Song, W.J. Underlying Role of Hydrophobic Environments in Tuning Metal Elements for Efficient Enzyme Catalysis. J. Am. Chem. Soc. 2023, 145, 5880–5887. [Google Scholar] [CrossRef]
- Fusetti, F.; Schröter, K.H.; Steiner, R.A.; van Noort, P.I.; Pijning, T.; Rozeboom, H.J.; Kalk, K.H.; Egmond, M.R.; Dijkstra, B.W. Crystal structure of the copper-containing quercetin 2,3-dioxygenase from Aspergillus japonicus. Structure 2002, 10, 259–268. [Google Scholar] [CrossRef]
- Gopal, B.; Madan, L.L.; Betz, S.F.; Kossiakoff, A.A. The Crystal Structure of a Quercetin 2,3-Dioxygenase from Bacillus subtilis Suggests Modulation of Enzyme Activity by a Change in the Metal Ion at the Active Site(s). Biochemistry 2005, 44, 193–201. [Google Scholar] [CrossRef]
- Tranchimand, S.; Ertel, G.; Gaydou, V.; Gaudin, C.; Tron, T.; Iacazio, G. Biochemical and molecular characterization of a quercetinase from Penicillium olsonii. Biochimie 2008, 90, 781–789. [Google Scholar] [CrossRef]
- Merkens, H.; Kappl, R.; Jakob, R.P.; Schmid, F.X.; Fetzner, S. Quercetinase QueD of Streptomyces sp. FLA, a Monocupin Dioxygenase with a Preference for Nickel and Cobalt. Biochemistry 2008, 47, 12185–12196. [Google Scholar] [CrossRef]
- Nianios, D.; Thierbach, S.; Steimer, L.; Lulchev, P.; Klostermeier, D.; Fetzner, S. Nickel quercetinase, a “promiscuous” metalloenzyme: Metal incorporation and metal ligand substitution studies. BMC Biochem. 2015, 16, 10. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.; Tian, G.; Liu, Y. Insights into the dioxygen activation and catalytic mechanism of the nickel-containing quercetinase. Catal. Sci. Technol. 2018, 8, 2340–2351. [Google Scholar] [CrossRef]
- Yan, X.; Song, J.; Lu, Q.; Li, C. Mechanistic insights into the crucial roles of Glu76 residue in nickel-dependent quercetin 2,4-dioxygenase for quercetin oxidative degradation. J. Catal. 2020, 387, 73–83. [Google Scholar] [CrossRef]
- El-Azhary, A.A.; Suter, H.U. Comparison between Optimized Geometries and Vibrational Frequencies Calculated by the DFT Methods. J. Phys. Chem. 1996, 100, 15056–15063. [Google Scholar] [CrossRef]
- Elstner, M.; Frauenheim, T.; Suhai, S. An approximate DFT method for QM/MM simulations of biological structures and processes. J. Mol. Struct. THEOCHEM 2003, 632, 29–41. [Google Scholar] [CrossRef]
- Caldeweyher, E.; Brandenburg, J.G. Simplified DFT methods for consistent structures and energies of large systems. J. Phys. Condens. Matter 2018, 30, 213001. [Google Scholar] [CrossRef]
- Mulholland, A.J. Chemical accuracy in QM/MM calculations on enzyme-catalysed reactions. Chem. Cent. J. 2007, 1, 19. [Google Scholar] [CrossRef]
- Senn, H.M.; Thiel, W. QM/MM studies of enzymes. Curr. Opin. Chem. Biol. 2007, 11, 182–187. [Google Scholar] [CrossRef]
- Jindal, G.; Warshel, A. Exploring the Dependence of QM/MM Calculations of Enzyme Catalysis on the Size of the QM Region. J. Phys. Chem. B 2016, 120, 9913–9921. [Google Scholar] [CrossRef]
- Al-Nu’airat, J.; Altarawneh, M.; Gao, X.; Westmoreland, P.R.; Dlugogorski, B.Z. Reaction of Aniline with Singlet Oxygen (O(2)(1)Delta(g)). J. Phys. Chem. A 2017, 121, 3199–3206. [Google Scholar] [CrossRef]
- Tian, G.; Su, H.; Liu, Y. Mechanism of Sulfoxidation and C–S Bond Formation Involved in the Biosynthesis of Ergothioneine Catalyzed by Ergothioneine Synthase (EgtB). ACS Catal. 2018, 8, 5875–5889. [Google Scholar] [CrossRef]
- Saito, T.; Kawakami, T.; Yamanaka, S.; Okumura, M. Computational Study of Catalytic Reaction of Quercetin 2,4-Dioxygenase. J. Phys. Chem. B 2015, 119, 6952–6962. [Google Scholar] [CrossRef] [PubMed]
- Siegbahn, P.E. Hybrid DFT study of the mechanism of quercetin 2,3-dioxygenase. Inorg. Chem. 2004, 43, 5944–5953. [Google Scholar] [CrossRef]
- Li, H.; Robertson, A.D.; Jensen, J.H. Very fast empirical prediction and rationalization of protein pKa values. Proteins 2005, 61, 704–721. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Brünger, A.T.; Karplus, M. Polar hydrogen positions in proteins: Empirical energy placement and neutron diffraction comparison. Proteins 1988, 4, 148–156. [Google Scholar] [CrossRef]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.; Mittal, J.; Feig, M.; Mackerell, A.D., Jr. Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone phi, psi and side-chain chi(1) and chi(2) dihedral angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Wang, B.; Johnston, E.M.; Li, P.; Shaik, S.; Davies, G.J.; Walton, P.H.; Rovira, C. QM/MM Studies into the H2O2-Dependent Activity of Lytic Polysaccharide Monooxygenases: Evidence for the Formation of a Caged Hydroxyl Radical Intermediate. ACS Catal. 2018, 8, 1346–1351. [Google Scholar] [CrossRef]
- Ahlrichs, R.; Bär, M.; Häser, M.; Horn, H.; Kölmel, C. Electronic structure calculations on workstation computers: The program system turbomole. Chem. Phys. Lett. 1989, 162, 165–169. [Google Scholar] [CrossRef]
- Smith, W.; Forester, T.R. DL_POLY_2.0: A general-purpose parallel molecular dynamics simulation package. J. Mol. Graph. Model. 1996, 14, 136–141. [Google Scholar] [CrossRef]
- Sherwood, P.; de Vries, A.H.; Guest, M.F.; Schreckenbach, G.; Catlow, C.A.; French, S.A.; Sokol, A.A.; Bromley, S.T.; Thiel, W.; Turner, A.J.; et al. QUASI: A general purpose implementation of the QM/MM approach and its application to problems in catalysis. J. Mol. Struct. THEOCHEM 2003, 632, 1–28. [Google Scholar] [CrossRef]
- Sherwood, P.; de Vries, A.H.; Collins, S.J.; Greatbanks, S.P.; Burton, N.A.; Vincent, M.A.; Hillier, I.H. Computer simulation of zeolite structure and reactivity using embedded cluster methods. Faraday Discuss. 1997, 106, 79–92. [Google Scholar] [CrossRef]
- Bakowies, D.; Thiel, W. Hybrid Models for Combined Quantum Mechanical and Molecular Mechanical Approaches. J. Phys. Chem. 1996, 100, 10580–10594. [Google Scholar] [CrossRef]
- Lee, C.; Yang, W.; Parr, R.G. Development of the Colle-Salvetti correlation-energy formula into a functional of the electron density. Phys. Rev. B 1988, 37, 785–789. [Google Scholar] [CrossRef]
- Weigend, F.; Ahlrichs, R. Balanced basis sets of split valence, triple zeta valence and quadruple zeta valence quality for H to Rn: Design and assessment of accuracy. Phys. Chem. Chem. Phys. 2005, 7, 3297–3305. [Google Scholar] [CrossRef] [PubMed]
- Kästner, J.; Carr, J.M.; Keal, T.W.; Thiel, W.; Wander, A.; Sherwood, P. DL-FIND: An Open-Source Geometry Optimizer for Atomistic Simulations. J. Phys. Chem. A 2009, 113, 11856–11865. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.C.; Nocedal, J. On the limited memory BFGS method for large scale optimization. Math. Program. 1989, 45, 503–528. [Google Scholar] [CrossRef]
- Kästner, J.; Sherwood, P. Superlinearly converging dimer method for transition state search. J. Chem. Phys. 2008, 128, 014106. [Google Scholar] [CrossRef]
- Grimme, S. Density functional theory with London dispersion corrections. WIREs Comput. Mol. Sci. 2011, 1, 211–228. [Google Scholar] [CrossRef]
- Neese, F. The ORCA program system. WIREs Comput. Mol. Sci. 2011, 2, 73–78. [Google Scholar] [CrossRef]
- Knizia, G.; Adler, T.B.; Werner, H.-J. Simplified CCSD(T)-F12 methods: Theory and benchmarks. J. Chem. Phys. 2009, 130, 054104. [Google Scholar] [CrossRef] [PubMed]
- Hohenstein, E.G.; Chill, S.T.; Sherrill, C.D. Assessment of the Performance of the M05−2X and M06−2X Exchange-Correlation Functionals for Noncovalent Interactions in Biomolecules. J. Chem. Theory Comput. 2008, 4, 1996–2000. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ΔE(Gas) (kcal/mol) | ΔE(PC) (kcal/mol) | ΔE(Gas-PC) (kcal/mol) |
|---|---|---|---|
| O2-Singlet | 10.5 | 10.5 | 0.0 |
| O2-Triplet | 0.0 | 0.0 | 0.0 |
| O2-MECP(S-T) | 27.0 | 25.9 | 1.1 |
| Ni+O2-Singlet | 0.0 | 0.0 | 0.0 |
| Ni+O2-Triplet | 6.4 | 6.2 | 0.2 |
| Ni+O2-MECP(S-T) | 18.5 | 16.1 | 2.4 |
| Que+O2-Singlet | 8.1 | 7.9 | 0.2 |
| Que+O2-Triplet | 0.0 | 0.0 | 0.0 |
| Que+O2-MECP(S-T) | 21.5 | 19.1 | 2.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Xiao, H.; Song, J.; Li, C. Unraveling the Pivotal Roles of Various Metal Ion Centers in the Catalysis of Quercetin 2,4-Dioxygenases. Molecules 2023, 28, 6238. https://doi.org/10.3390/molecules28176238
Yan X, Xiao H, Song J, Li C. Unraveling the Pivotal Roles of Various Metal Ion Centers in the Catalysis of Quercetin 2,4-Dioxygenases. Molecules. 2023; 28(17):6238. https://doi.org/10.3390/molecules28176238
Chicago/Turabian StyleYan, Xueyuan, Han Xiao, Jinshuai Song, and Chunsen Li. 2023. "Unraveling the Pivotal Roles of Various Metal Ion Centers in the Catalysis of Quercetin 2,4-Dioxygenases" Molecules 28, no. 17: 6238. https://doi.org/10.3390/molecules28176238
APA StyleYan, X., Xiao, H., Song, J., & Li, C. (2023). Unraveling the Pivotal Roles of Various Metal Ion Centers in the Catalysis of Quercetin 2,4-Dioxygenases. Molecules, 28(17), 6238. https://doi.org/10.3390/molecules28176238