
Potential of Natural Phenolic Compounds as Antimicrobial Agents against Multidrug-Resistant Staphylococcus aureus in Chicken Meat
 ,
,  ,
,  , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. Phenotypic Characterizations and Incidence of CPS
2.2. Coagulase-Positive Staphylococci Incidence
2.3. Antimicrobial Susceptibilities Test
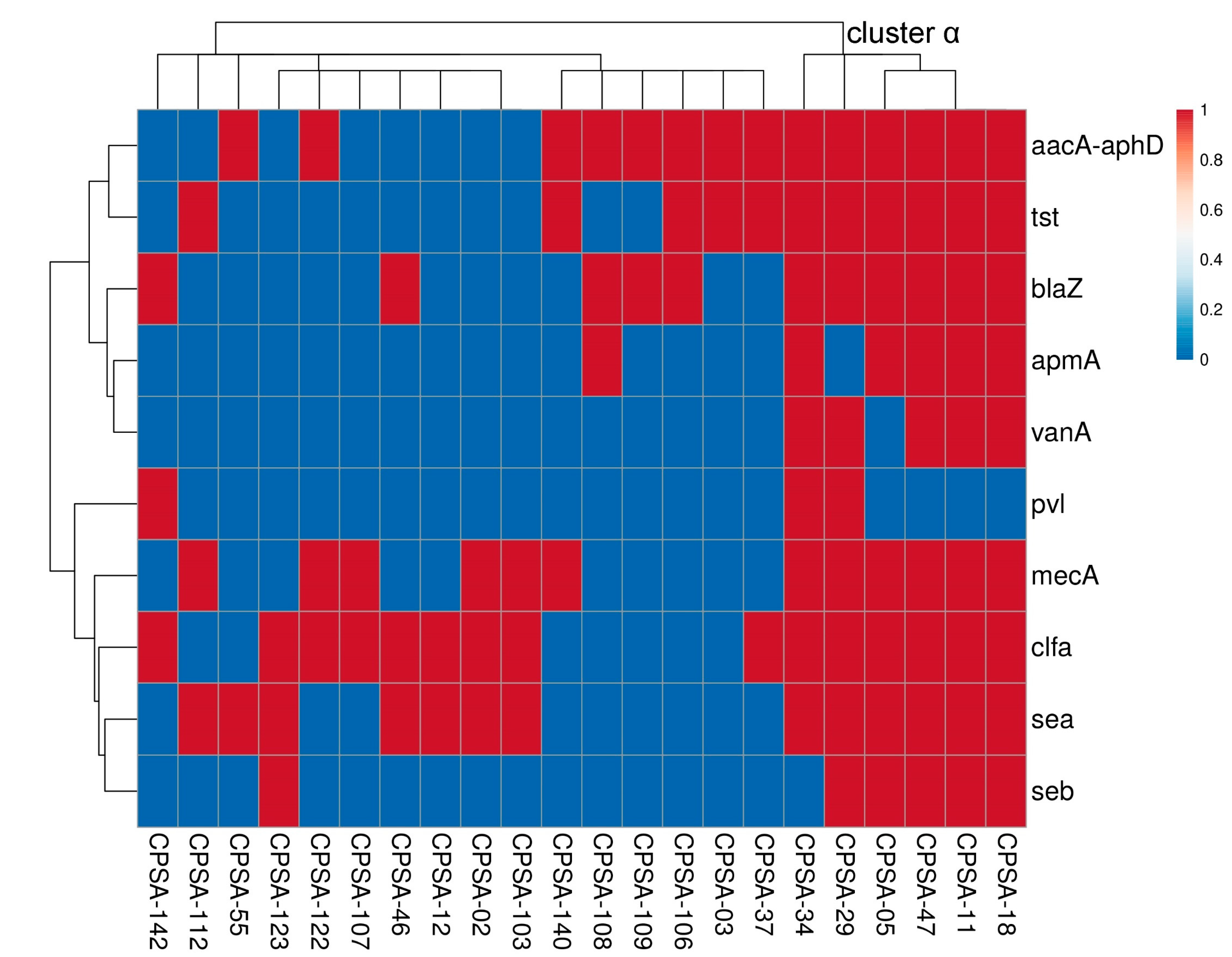
2.4. Molecular Characterization of S. aureus
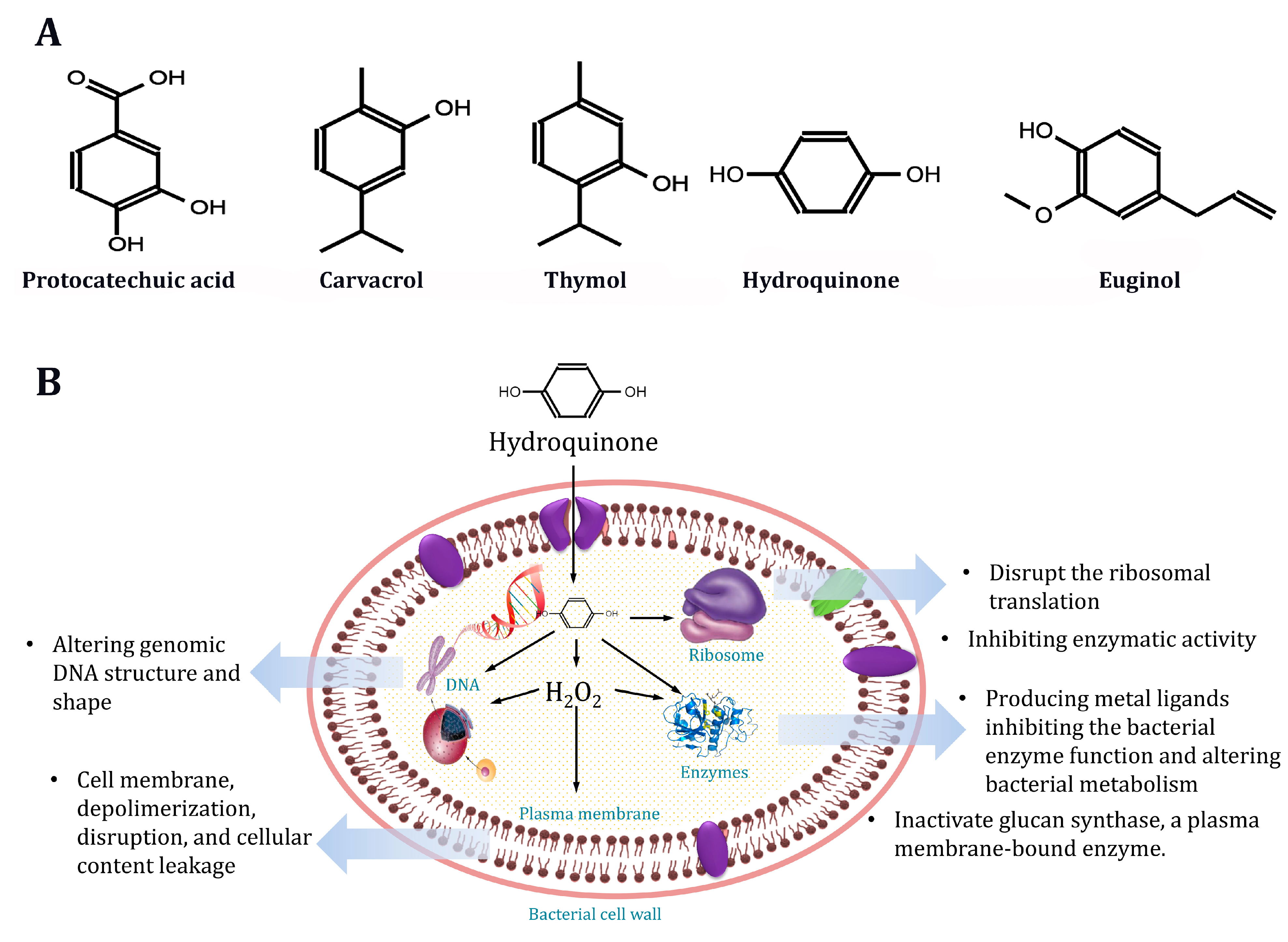
2.5. Antimicrobial Activities of Natural Phenolics
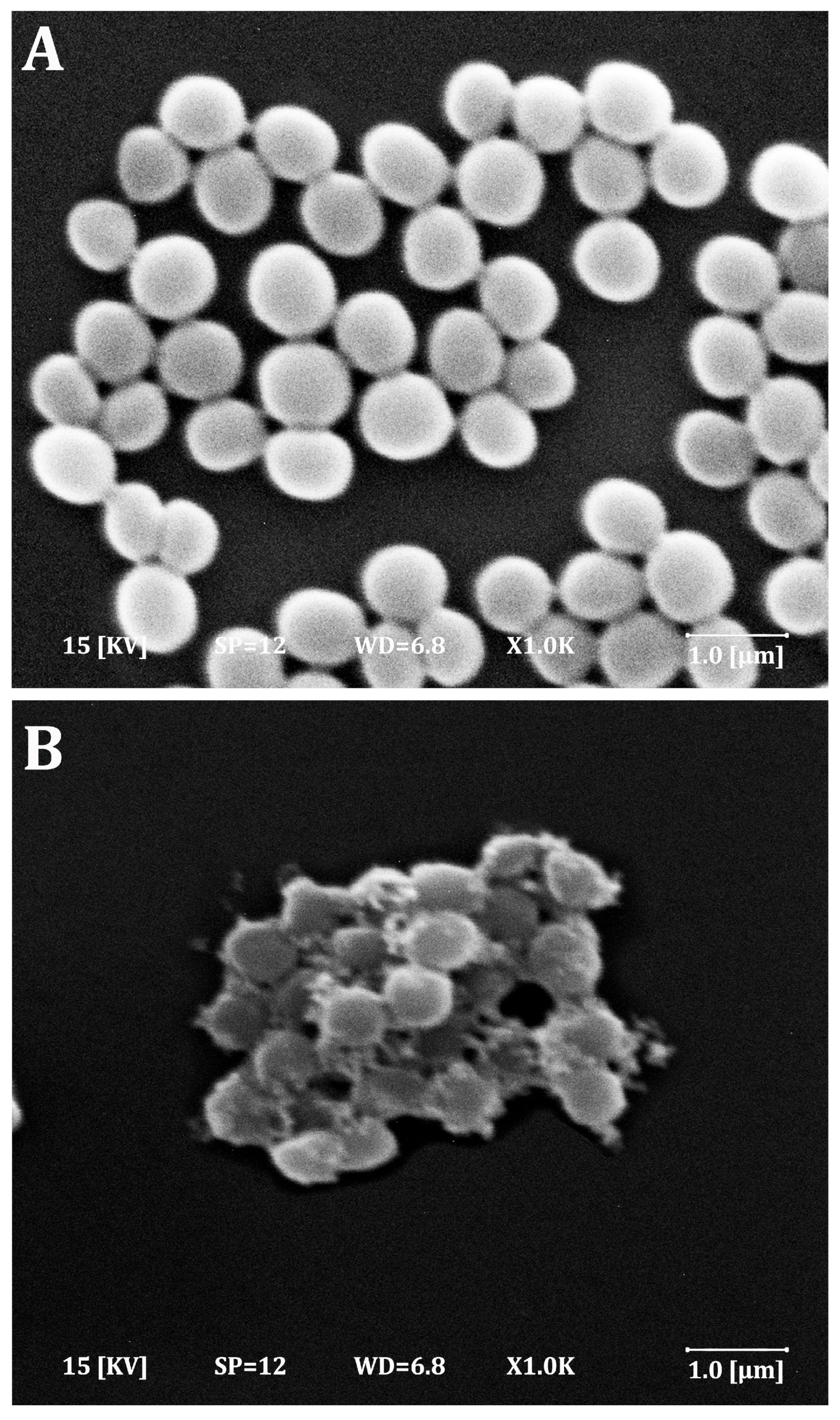
2.6. Effect of Phenolic Compounds on S. aureus Morphology
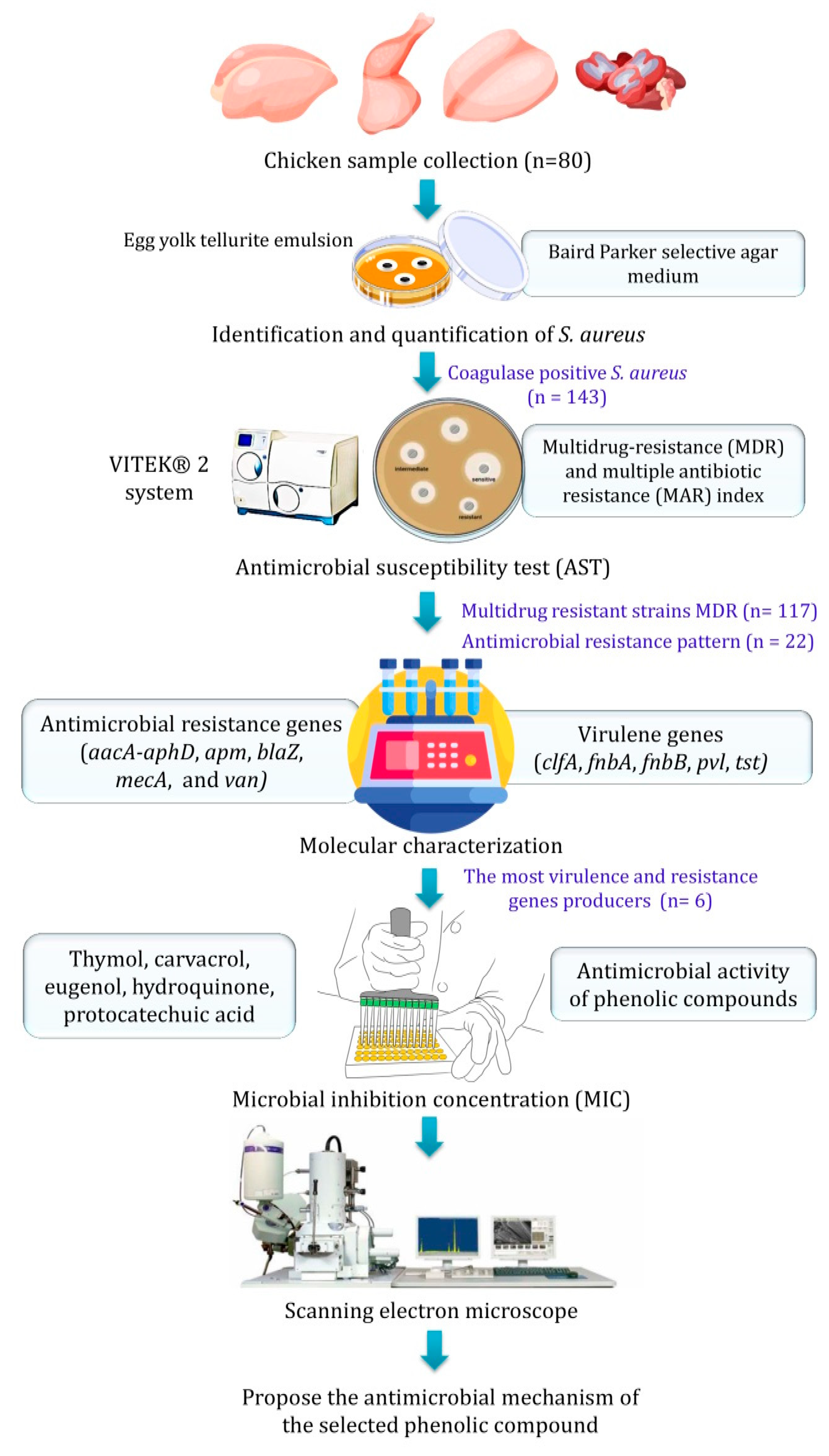
3. Materials and Methods
3.1. Chicken Sample Collection
3.2. Identification and Quantification of S. aureus
3.3. Antimicrobial Susceptibility Test (AST)
3.4. Molecular Identification
3.5. Phenolic Compounds Preparation
3.6. Microbial Inhibition Concentration (MIC)
3.7. Scanning Electron Microscopy (SEM)
3.8. Statistical Analysis
4. Discussion
4.1. Phenotypic Characterizations and Incidence of CPS
4.2. Coagulase-Positive Staphylococci Incidence
4.3. Antimicrobial Susceptibility Test
4.4. Molecular Characterization of S. aureus
4.5. Antimicrobial Activities of Natural Phenolics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Boqvist, S.; Söderqvist, K.; Vågsholm, I. Food Safety Challenges and One Health within Europe. Acta Vet. Scand. 2018, 60, 1. [Google Scholar] [CrossRef] [PubMed]
- Uyttendaele, M.; Franz, E.; Schlüter, O. Food Safety, a Global Challenge. Int. J. Environ. Res. Public Health 2015, 13, 67. [Google Scholar] [CrossRef]
- Lund, B.M. Provision of Microbiologically Safe Food for Vulnerable People in Hospitals, Care Homes and in the Community. Food Control 2019, 96, 535–547. [Google Scholar] [CrossRef]
- Jaffee, S.; Henson, S.; Unnevehr, L.; Grace, D.; Cassou, E. The Safe Food Imperative: Accelerating Progress in Low- and Middle-Income Countries; World Bank Publications: Washington, DC, USA, 2018. [Google Scholar]
- Morshdy, A.E.M.; Darwish, W.; Mohammed, F.M.; Mahmoud, A.F.A. Bacteriological Quality of Retailed Chicken Meat Products in Zagazig City, Egypt. J. Adv. Vet. Res. 2023, 13, 47–51. [Google Scholar]
- Mahmoud, A.F.; Escrivá, L.; Rodríguez-Carrasco, Y.; Moltó, J.C.; Berrada, H. Determination of Trichothecenes in Chicken Liver Using Gas Chromatography Coupled with Triple-Quadrupole Mass Spectrometry. LWT 2018, 93, 237–242. [Google Scholar] [CrossRef]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-Resistant Staphylococcus aureus. Nat. Rev. Dis. Prim. 2018, 4, 18033. [Google Scholar] [CrossRef]
- Grace, D.; Fetsch, A. Staphylococcus aureus—A foodborne pathogen: Epidemiology, detection, characterization, prevention, and control: An overview. In Staphylococcus aureus; Academic Press: Cambridge, MA, USA, 2018; pp. 3–10. [Google Scholar]
- Sugrue, I.; Tobin, C.; Ross, R.P.; Stanton, C.; Hill, C. Foodborne Pathogens and Zoonotic Diseases. In Raw Milk; Academic Press: Cambridge, MA, USA, 2019; pp. 259–272. [Google Scholar]
- Logue, C.M.; Barbieri, N.L.; Nielsen, D.W. Pathogens of food animals: Sources, characteristics, human risk, and methods of detection. Adv. Food Nutr. Res. 2017, 82, 277–365. [Google Scholar]
- Al-sherees, H.A.A. Molecular Diagnosis of Aminoglycoside Transferase Genes in Methicillin-Resistant Staphylococcus aureus (MRSA) Strains. Indian J. Nat. Sci. 2019, 10, 166–171. [Google Scholar]
- Mahros, M.A.; Abd-Elghany, S.M.; Sallam, K.I. Multidrug-, Methicillin-, and Vancomycin-Resistant Staphylococcus aureus Isolated from Ready-to-Eat Meat Sandwiches: An Ongoing Food and Public Health Concern. Int. J. Food Microbiol. 2021, 346, 109165. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Al Naiemi, N.; Elsohaby, I.; Mahmoud, A.F.A.; Salem, G.A.; Vandenbroucke-Grauls, C.M.J.E. Prevalence of Extended-Spectrum β-Lactamase-Producing Enterobacterales in Retail Sheep Meat from Zagazig City, Egypt. BMC Vet. Res. 2022, 18, 191. [Google Scholar] [CrossRef]
- Ali Alghamdi, B.; Al-Johani, I.; Al-Shamrani, J.M.; Musamed Alshamrani, H.; Al-Otaibi, B.G.; Almazmomi, K.; Yusnoraini Yusof, N. Antimicrobial Resistance in Methicillin-Resistant Staphylococcus aureus. Saudi J. Biol. Sci. 2023, 30, 103604. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jung, D.; O’Brien, B.; Ruffini, J.; Dussault, F.; Dube-Duquette, A.; Demontier, É.; Lucier, J.-F.; Malouin, F.; Dufour, S.; et al. Comparative Genomic Analysis of Staphylococcus aureus Isolates Associated with Either Bovine Intramammary Infections or Human Infections Demonstrates the Importance of Restriction-Modification Systems in Host Adaptation. Microb. Genom. 2022, 8, 779. [Google Scholar] [CrossRef] [PubMed]
- Mama, O.M.; Morales, L.; Ruiz-Ripa, L.; Zarazaga, M.; Torres, C. High Prevalence of Multidrug Resistant S. aureus-CC398 and Frequent Detection of Enterotoxin Genes among Non-CC398 S. aureus from Pig-Derived Food in Spain. Int. J. Food Microbiol. 2020, 320, 108510. [Google Scholar] [CrossRef] [PubMed]
- Morshdy, A.E.M.A.; Mahmoud, A.F.A.; Khalifa, S.M.; El-Dien, W.M.S.; Darwish, W.S.; El Bayomi, R. Prevalence of Staphylococcus aureus and Salmonella species in chicken meat products retailed in Egypt. Slov. Vet. Res. 2023, 60, 425–432. [Google Scholar] [CrossRef]
- Silva, V.; Vieira-Pinto, M.; Saraiva, C.; Manageiro, V.; Reis, L.; Ferreira, E.; Caniça, M.; Capelo, J.L.; Igrejas, G.; Poeta, P. Prevalence and Characteristics of Multidrug-Resistant Livestock-Associated Methicillin-Resistant Staphylococcus aureus (LA-MRSA) CC398 Isolated from Quails (Coturnix Coturnix Japonica) Slaughtered for Human Consumption. Animals 2021, 11, 2038. [Google Scholar] [CrossRef]
- Abolghait, S.K.; Fathi, A.G.; Youssef, F.M.; Algammal, A.M. Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Chicken Meat and Giblets Often Produces Staphylococcal Enterotoxin B (SEB) in Non-Refrigerated Raw Chicken Livers. Int. J. Food Microbiol. 2020, 328, 108669. [Google Scholar] [CrossRef]
- Uruén, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Biofilms as Promoters of Bacterial Antibiotic Resistance and Tolerance. Antibiotics 2020, 10, 3. [Google Scholar] [CrossRef]
- Artini, M.; Papa, R.; Scoarughi, G.L.; Galano, E.; Barbato, G.; Pucci, P.; Selan, L. Comparison of the Action of Different Proteases on Virulence Properties Related to the Staphylococcal Surface. J. Appl. Microbiol. 2012, 114, 266–277. [Google Scholar] [CrossRef]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum Sensing Mediated Pathogenicity in Pseudomonas Aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Dula, S.; Ajayeoba, T.A.; Ijabadeniyi, O.A. Bacterial Biofilm Formation on Stainless Steel in the Food Processing Environment and Its Health Implications. Folia Microbiol. 2021, 66, 293–302. [Google Scholar] [CrossRef]
- Senobar Tahaei, S.A.; Stájer, A.; Barrak, I.; Ostorházi, E.; Szabó, D.; Gajdács, M. Correlation Between Biofilm-Formation and the Antibiotic Resistant Phenotype in Staphylococcus aureus Isolates: A Laboratory-Based Study in Hungary and a Review of the Literature. Infect. Drug Resist. 2021, 14, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Craft, K.M.; Nguyen, J.M.; Berg, L.J.; Townsend, S.D. Methicillin-Resistant Staphylococcus aureus (MRSA): Antibiotic-Resistance and the Biofilm Phenotype. Medchemcomm 2019, 10, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, H.G.; Pan, H.Y.; Yan, S.M.; Qi, Z.X.; Zhao, X.L.; Luo, D.Q. Synthesis and Antifungal Activity of Polyphenol Ether Derivatives against Plant Pathogenic Fungi in Vitro and in Vivo. Rev. Roum. Chim. 2022, 67, 373–383. [Google Scholar]
- Morshdy, A.E.M.A.; El-Tahlawy, A.S.; Qari, S.H.; Qumsani, A.T.; Bay, D.H.; Sami, R.; Althubaiti, E.H.; Mansour, A.M.A.; Aljahani, A.H.; Hafez, A.E.-S.E.; et al. Anti-Biofilms’ Activity of Garlic and Thyme Essential Oils against Salmonella Typhimurium. Molecules 2022, 27, 2182. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A. Effect of Lettuce, Marjoram and Cumin Essential Oils on the Quality and Shelf Life of Minced Meat during Refrigerated Storage. Zagazig Vet. J. 2019, 47, 288–297. [Google Scholar] [CrossRef]
- El Bayomi, R.M.; Hebishy, R.M.; Darwish, W.S.; El-Atabany, A.I.M.; Mahmoud, A.F.A. Mould Contamination of Some Meat Products with Reference to Decontamination Trials of Aspergillus Flavus Using Essential Oils. Slov. Vet. Res. 2021, 58, 363–372. [Google Scholar]
- Morshdy, A.E.M.; Nahla, B.M.; Shafik, S.; Hussein, M.A. Antimicrobial Effect of Essential Oils on Multidrug-Resistant Salmonella Typhimurium in Chicken Fillets. Pak. Vet. J. 2021, 41, 545–551. [Google Scholar] [CrossRef]
- Liu, J.; Du, C.; Beaman, H.T.; Monroe, M.B.B. Characterization of Phenolic Acid Antimicrobial and Antioxidant Structure-Property Relationships. Pharmaceutics 2020, 12, 419. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A Major Update to the DrugBank Database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Zhang, J.; Xu, D.; Li, Y.; Liu, Y.; Zhang, X.; Zhang, R.; Wu, Z.; Weng, P. Antimicrobial Effect of Tea Polyphenols against Foodborne Pathogens: A Review. J. Food Prot. 2021, 84, 1801–1808. [Google Scholar] [CrossRef]
- Mahmoud, A.F.A.; Elshopary, N.; Hasab El-Naby, G.; El Bayomi, R. Reduction of biogenic amines production in chilled minced meat using antimicrobial seasonings. J. Microbiol. Biotechnol. Food Sci. 2021, 10, e3663. [Google Scholar] [CrossRef]
- Eldaly, E.A.; Mahmoud, A.F.A.; Abobakr, H.M. Preservative Effect of Chitosan Coating on Shelf Life and Sensory Properties of Chicken Fillets during Chilled Storage. J. Nutr. Food Secur. 2018, 3, 139–148. [Google Scholar]
- Sary, S.; Shiwei, X.; Wen, Y.; Darith, S.; Chorn, S. Household Food Consumption in Rural, Cambodia Almost Ideal Demand System Analysis. J. Phys. Conf. Ser. 2019, 1176, 42077. [Google Scholar] [CrossRef]
- Fasanmi, O.G.; Makinde, G.E.O.; Popoola, M.A.; Fasina, O.F.; Matere, J.; Kehinde, O.O.; Balogun, F.A.; Ogundare, S.T. Potential Risk Factors Associated with Carcass Contamination in Slaughterhouse Operations and Hygiene in Oyo State, Nigeria. Int. J. Livest. Prod. 2018, 9, 211–220. [Google Scholar] [CrossRef]
- Guédé, S.S.; Soumahoro, S.; Mbari, K.B.; Touré, A.; Soro, Y.R. Hygienic Quality of Chicken Grills Sold along the Streets of Korhogo Town (North of Côte d’Ivoire). World J. Adv. Res. Rev. 2022, 16, 539–551. [Google Scholar] [CrossRef]
- Augustin, J.-C.; Kooh, P.; Bayeux, T.; Guillier, L.; Meyer, T.; Jourdan-Da Silva, N.; Villena, I.; Sanaa, M.; Cerf, O.; on Behalf of the Anses Working Group on Consumer Information on Foodborne Biological Risks. Contribution of Foods and Poor Food-Handling Practices to the Burden of Foodborne Infectious Diseases in France. Foods 2020, 9, 1644. [Google Scholar] [CrossRef]
- Gutiérrez-Larraínzar, M.; Rúa, J.; Caro, I.; de Castro, C.; de Arriaga, D.; García-Armesto, M.R.; del Valle, P. Evaluation of Antimicrobial and Antioxidant Activities of Natural Phenolic Compounds against Foodborne Pathogens and Spoilage Bacteria. Food Control 2012, 26, 555–563. [Google Scholar] [CrossRef]
- Egyptian Organization for Standardization and Quality (EOS). Egyptian Organization for Standardization and Quality for Chilled Poultry and Rabbit, No. 1651; EOS: Cairo, Egypt, 2005. [Google Scholar]
- Health Protection Agency. Microbiological Safety of Ready-to-Eat Foods; Health Protection Agency: London, UK, 2009. [Google Scholar]
- ISO 6888-1:1999/AMD 2:2018; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Coagulase Positive Staphylococci (Staphylococcus aureus and Other Species)—Part 1: Technique Using Baird-Parker Agar Medium—Amendment 2: Inclusion of an Alternative Confirmation Test Using RPFA Stab Method. ISO: Geneva, Switzerland, 2018.
- Wang, H.; Wang, H.; Liang, L.; Xu, X.; Zhou, G. Prevalence, Genetic Characterization and Biofilm Formation in Vitro of Staphylococcus aureus Isolated from Raw Chicken Meat at Retail Level in Nanjing, China. Food Control 2018, 86, 11–18. [Google Scholar] [CrossRef]
- Deidei, H.B.; Aleruchi, O.; Stephenson, D.L.; Igoni, G.Y. Antibiogram of Staphylococcus Species Isolated from Some Abattoirs in Rivers State, Nigeria. South Asian J. Res. Microbiol. 2023, 15, 1–8. [Google Scholar] [CrossRef]
- Kmeť, V.; Drugdová, Z. Antimicrobial Susceptibility of Microflora from Ovine Cheese. Folia Microbiol. 2012, 57, 291–293. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing. In CLSI Supplement M100, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Rafailidis, P.I.; Kofteridis, D. Proposed Amendments Regarding the Definitions of Multidrug-Resistant and Extensively Drug-Resistant Bacteria. Expert Rev. Anti. Infect. Ther. 2021, 20, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for Detection of Genes for Staphylococcus aureus Enterotoxins, Exfoliative Toxins, Toxic Shock Syndrome Toxin 1, and Methicillin Resistance. J. Clin. Microbiol. 2000, 38, 1032–1035. [Google Scholar] [CrossRef]
- Tristan, A.; Ying, L.; Bes, M.; Etienne, J.; Vandenesch, F.; Lina, G. Use of Multiplex PCR to Identify Staphylococcus aureus Adhesins Involved in Human Hematogenous Infections. J. Clin. Microbiol. 2003, 41, 4465–4467. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, F.; Zhao, J.; Lou, N.; Li, C.; Huang, T.; Li, Y. Molecular Characteristics and Virulence Gene Profiles of Staphylococcus aureus Causing Bloodstream Infection. Braz. J. Infect. Dis. 2018, 22, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR Assay for Simultaneous Detection of Nine Clinically Relevant Antibiotic Resistance Genes in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 4089–4094. [Google Scholar] [CrossRef]
- Azimian, A.; Havaei, S.A.; Fazeli, H.; Naderi, M.; Ghazvini, K.; Samiee, S.M.; Soleimani, M.; Peerayeh, S.N. Genetic Characterization of a Vancomycin-Resistant Staphylococcus aureus Isolate from the Respiratory Tract of a Patient in a University Hospital in Northeastern Iran. J. Clin. Microbiol. 2012, 50, 3581–3585. [Google Scholar] [CrossRef]
- Martineau, F.; Picard, F.J.; Lansac, N.; Ménard, C.; Roy, P.H.; Ouellette, M.; Bergeron, M.G. Correlation between the Resistance Genotype Determined by Multiplex PCR Assays and the Antibiotic Susceptibility Patterns of Staphylococcus aureus and Staphylococcus epidermidis. Antimicrob. Agents Chemother. 2000, 44, 231–238. [Google Scholar] [CrossRef]
- Fessler, A.T.; Kadlec, K.; Hassel, M.; Hauschild, T.; Eidam, C.; Ehricht, R.; Monecke, S.; Schwarz, S. Characterization of Methicillin-Resistant Staphylococcus aureus Isolates from Food and Food Products of Poultry Origin in Germany. Appl. Environ. Microbiol. 2011, 77, 7151–7157. [Google Scholar] [CrossRef]
- ISO 20776-1:2006; Clinical Laboratory Testing and In Vitro Diagnostic Test Systems—Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices—Part 1: Reference Method for Testing the In Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases. International Organization for Standardization (ISO): Geneva, Switzerland, 2006.
- Konstantinovitch, K.Y.; Arsene, M.M.J.; Aliya, M.V.; Viktorovna, P.I.; Elena, V.G.; Azova, M.M.; Amira, A.A. Assessment of Antimicrobial Activity of Ethanolic and Aqueous Extracts of Aesculus hippocastanum L. (Horse Chestnut) Bark against Bacteria Isolated from Urine of Patients Diagnosed Positive to Urinary Tract Infections. Front. Biosci. 2022, 14, 11. [Google Scholar] [CrossRef]
- Amoako, D.G.; Somboro, A.M.; Abia, A.L.K.; Molechan, C.; Perrett, K.; Bester, L.A.; Essack, S.Y. Antibiotic Resistance in Staphylococcus aureus from Poultry and Poultry Products in UMgungundlovu District, South Africa, Using the “Farm to Fork” Approach. Microb. Drug Resist. 2020, 26, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Rortana, C.; Nguyen-Viet, H.; Tum, S.; Unger, F.; Boqvist, S.; Dang-Xuan, S.; Koam, S.; Grace, D.; Osbjer, K.; Heng, T.; et al. Prevalence of Salmonella spp. and Staphylococcus aureus in Chicken Meat and Pork from Cambodian Markets. Pathogens 2021, 10, 556. [Google Scholar] [CrossRef] [PubMed]
- Savariraj, W.R.; Ravindran, N.B.; Kannan, P.; Rao, V.A. Occurrence and Enterotoxin Gene Profiles of Staphylococcus aureus Isolated from Retail Chicken Meat. Food Sci. Technol. Int. 2020, 27, 619–625. [Google Scholar] [CrossRef]
- Narvaez-Bravo, C.; Toufeer, M.; Weese, S.J.; Diarra, M.S.; Deckert, A.E.; Reid-Smith, R.; Aslam, M. Prevalence of Methicillin-Resistant Staphylococcus aureus in Canadian Commercial Pork Processing Plants. J. Appl. Microbiol. 2016, 120, 770–780. [Google Scholar] [CrossRef]
- Davis, R.; Brown, P.D. Multiple Antibiotic Resistance Index, Fitness and Virulence Potential in Respiratory Pseudomonas aeruginosa from Jamaica. J. Med. Microbiol. 2016, 65, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Riedel, S.; Vijayakumar, D.; Berg, G.; Kang, A.D.; Smith, K.P.; Kirby, J.E. Evaluation of Apramycin against Spectinomycin-Resistant and -Susceptible Strains of Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2019, 74, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Igbinosa, E.O.; Beshiru, A.; Igbinosa, I.H.; Ogofure, A.G.; Ekundayo, T.C.; Okoh, A.I. Prevalence, Multiple Antibiotic Resistance and Virulence Profile of Methicillin-Resistant Staphylococcus aureus (MRSA) in Retail Poultry Meat from Edo, Nigeria. Front. Cell. Infect. Microbiol. 2023, 13, 1122059. [Google Scholar] [CrossRef]
- Ruzauskas, M.; Siugzdiniene, R.; Butrimaite-Ambrozeviciene, C.; Zymantiene, J.; KlimienE, I.; Vaskeviciute, L.; Mockeliunas, R.; Virgailis, M. Prevalence and Characterization of Multi-Resistant Staphylococcus spp. Isolated from Poultry Liver. J. Food Saf. 2016, 36, 508–514. [Google Scholar] [CrossRef]
- Parvin, M.S.; Ali, M.Y.; Talukder, S.; Nahar, A.; Chowdhury, E.H.; Rahman, M.T.; Islam, M.T. Prevalence and Multidrug Resistance Pattern of Methicillin Resistant S. aureus Isolated from Frozen Chicken Meat in Bangladesh. Microorganisms 2021, 9, 636. [Google Scholar] [CrossRef]
- Zehra, A.; Gulzar, M.; Singh, R.; Kaur, S.; Gill, J.P.S. Corrigendum to ‘Prevalence, multidrug resistance and molecular typing of methicillin-resistant Staphylococcus aureus (MRSA) in retail meat from Punjab, India’ [J. Glob. Antimicrob. Resist. 16 (2019) 152–158]. J. Glob. Antimicrob. Resist. 2020, 22, 866. [Google Scholar] [CrossRef]
- Rychshanova, R.; Mendybayeva, A.; Miciński, B.; Mamiyev, N.; Shevchenko, P.; Bermukhametov, Z.; Orzechowski, B.; Miciński, J. Antibiotic Resistance and Biofilm Formation in Staphylococcus aureus Isolated from Dairy Cows at the Stage of Subclinical Mastitis in Northern Kazakhstan. Arch. Anim. Breed. 2022, 65, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Schwarz, S.; Li, Y.; Shen, Z.; Zhang, Q.; Wu, C.; Shen, J. Transferable Multiresistance Plasmids Carrying Cfr in Enterococcus spp. from Swine and Farm Environment. Antimicrob. Agents Chemother. 2013, 57, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.; Stojković, V.; Noda-Garcia, L.; Young, I.D.; Myasnikov, A.G.; Kleinman, J.; Palla, A.; Floor, S.N.; Frost, A.; Fraser, J.S.; et al. Directed Evolution of the RRNA Methylating Enzyme Cfr Reveals Molecular Basis of Antibiotic Resistance. eLife 2022, 11, e70017. [Google Scholar] [CrossRef] [PubMed]
- Bordeleau, E.; Stogios, P.J.; Evdokimova, E.; Koteva, K.; Savchenko, A.; Wright, G.D. ApmA Is a Unique Aminoglycoside Antibiotic Acetyltransferase That Inactivates Apramycin. mBio 2021, 12, e02705-20. [Google Scholar] [CrossRef]
- Mun, S.-H.; Kong, R.; Seo, Y.-S.; Zhou, T.; Kang, O.-H.; Shin, D.-W.; Kwon, D.-Y. Subinhibitory Concentrations of Punicalagin Reduces Expression of Virulence-Related Exoproteins by Staphylococcus aureus. FEMS Microbiol. Lett. 2016, 363, fnw253. [Google Scholar] [CrossRef]
- Langley, R.J.; Fraser, J.D.; Proft, T. Bacterial Superantigens and Superantigen-like Toxins. In The Comprehensive Sourcebook of Bacterial Protein Toxins; Academic Press: London, UK, 2015; pp. 911–974. [Google Scholar]
- Chen, H.; Zhang, J.; He, Y.; Lv, Z.; Liang, Z.; Chen, J.; Li, P.; Liu, J.; Yang, H.; Tao, A.; et al. Exploring the Role of Staphylococcus aureus in Inflammatory Diseases. Toxins 2022, 14, 464. [Google Scholar] [CrossRef]
- Jhelum, H.; Čerina, D.; Harbort, C.J.; Lindner, A.; Hanitsch, L.G.; Leistner, R.; Schröder, J.-T.; von Bernuth, H.; Stegemann, M.S.; Schürmann, M.; et al. Patients with recurrent PVL+-Staphylococcus aureus infections show enhanced sensitivity to PVL-mediated formation of atypical NETs. bioRxiv 2021. [Google Scholar] [CrossRef]
- AL-Mamun, M.; Chowdhury, T.; Biswas, B.; Absar, N. Food Poisoning and Intoxication: A Global Leading Concern for Human Health. In Food Safety and Preservation; Academic Press: Cambridge, MA, USA, 2018; pp. 307–352. [Google Scholar]
- Negrón, O.; Weggeman, M.; Grimbergen, J.; Clark, E.G.; Abrahams, S.; Hur, W.S.; Koopman, J.; Flick, M.J. Fibrinogen γ′ Promotes Host Survival during Staphylococcus aureus Septicemia in Mice. J. Thromb. Haemost. 2023, 21, 2277–2290. [Google Scholar] [CrossRef]
- Hu, D.-L.; Li, S.; Fang, R.; Ono, H.K. Update on Molecular Diversity and Multipathogenicity of Staphylococcal Superantigen Toxins. Anim. Dis. 2021, 1, 7. [Google Scholar] [CrossRef]
- Pérez, V.K.C.; da Costa, G.M.; Guimarães, A.S.; Heinemann, M.B.; Lage, A.P.; Dorneles, E.M.S. Relationship between Virulence Factors and Antimicrobial Resistance in Staphylococcus aureus from Bovine Mastitis. J. Glob. Antimicrob. Resist. 2020, 22, 792–802. [Google Scholar] [CrossRef]
- Rúa, J.; Fernández-Álvarez, L.; de Castro, C.; del Valle, P.; de Arriaga, D.; García-Armesto, M.R. Antibacterial Activity Against Foodborne Staphylococcus aureus and Antioxidant Capacity of Various Pure Phenolic Compounds. Foodborne Pathog. Dis. 2011, 8, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; He, N.; Zhao, Y.; Xia, D.; Wei, J.; Kang, W. Antimicrobial Mechanism of Hydroquinone. Appl. Biochem. Biotechnol. 2019, 189, 1291–1303. [Google Scholar] [CrossRef]
- Lobiuc, A.; Pavăl, N.-E.; Mangalagiu, I.I.; Gheorghiță, R.; Teliban, G.-C.; Amăriucăi-Mantu, D.; Stoleru, V. Future Antimicrobials: Natural and Functionalized Phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef] [PubMed]
- Efenberger-Szmechtyk, M.; Nowak, A.; Czyzowska, A. Plant Extracts Rich in Polyphenols: Antibacterial Agents and Natural Preservatives for Meat and Meat Products. Crit. Rev. Food Sci. Nutr. 2020, 61, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wei, J.; Qiu, Y.; Niu, C.; Song, Z.; Yuan, Y.; Yue, T. Structure-Dependent Inhibition of Stenotrophomonas maltophilia by Polyphenol and Its Impact on Cell Membrane. Front. Microbiol. 2019, 10, 2646. [Google Scholar] [CrossRef]
- Li, Y.-Q.; Han, Q.; Feng, J.-L.; Tian, W.-L.; Mo, H.-Z. Antibacterial Characteristics and Mechanisms of Ɛ-Poly-Lysine against Escherichia Coli and Staphylococcus aureus. Food Control 2014, 43, 22–27. [Google Scholar] [CrossRef]
- Guo, L.; Sun, Q.; Gong, S.; Bi, X.; Jiang, W.; Xue, W.; Fei, P. Antimicrobial Activity and Action Approach of the Olive Oil Polyphenol Extract Against Listeria monocytogenes. Front. Microbiol. 2019, 10, 1586. [Google Scholar] [CrossRef]
- Gogoi, J.; Nakhuru, K.S.; Policegoudra, R.S.; Chattopadhyay, P.; Rai, A.K.; Veer, V. Isolation and Characterization of Bioactive Components from Mirabilis jalapa L. Radix. J. Tradit. Complement. Med. 2015, 6, 41–47. [Google Scholar] [CrossRef]
- Hirakawa, K.; Sano, S. Platinum Nanoparticle Catalyst Scavenges Hydrogen Peroxide Generated from Hydroquinone. Bull. Chem. Soc. Jpn. 2009, 82, 1299–1303. [Google Scholar] [CrossRef]
- Lin, S.; Wang, Z.; Lam, K.-L.; Zeng, S.; Tan, B.K.; Hu, J. Role of Intestinal Microecology in the Regulation of Energy Metabolism by Dietary Polyphenols and Their Metabolites. Food Nutr. Res. 2019, 63, 1518. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. P-Coumaric Acid Kills Bacteria through Dual Damage Mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Röttig, A.; Steinbüchel, A. Acyltransferases in Bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 277–321. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Samples (No.) | Positive Sample No. (%) | Coagulase-Positive S. aureus Counts (log CFU/g) | Microbiological Quality of Tested Samples (log CFU/g) | ||||
|---|---|---|---|---|---|---|---|
| Min. | Max. | Mean ± SE | Satisfactory (<2 log CFU/g) * | Unsatisfactory (2–<4 log CFU/g) | Unacceptable (≥4 log CFU/g) | ||
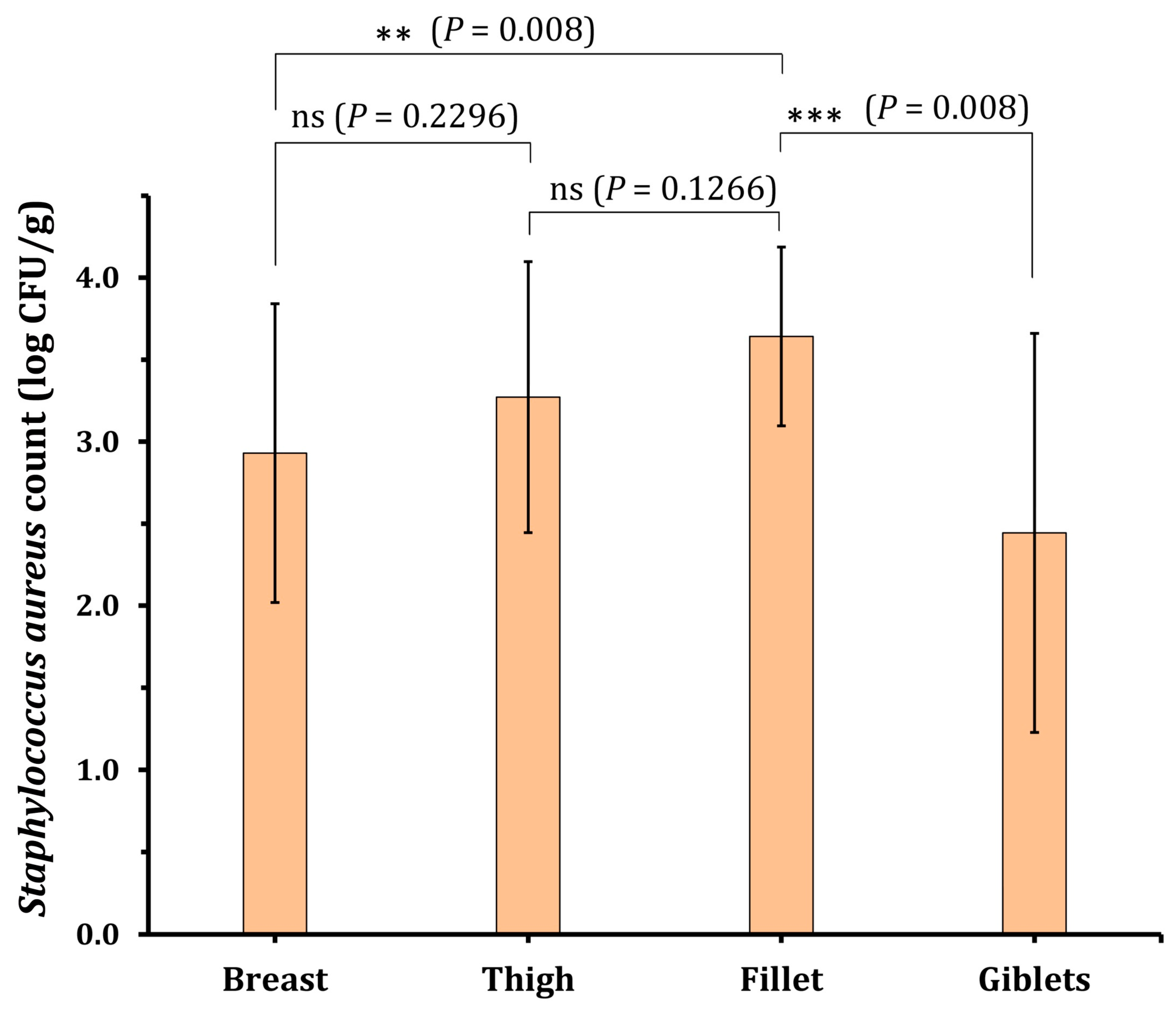
| Breast (20) | 20 (100) | 0.9 | 4.8 | 2.9 ± 0.9 | 3 (15.0%) | 14 (70.0%) | 3 (15.0%) |
| Thigh (20) | 19 (95) | 1.55 | 4.8 | 3.3 ± 0.8 | 1 (5.3%) | 13 (68.4%) | 5 (26.3%) |
| Giblets (20) | 17 (85) | 0.48 | 4.12 | 2.4 ± 1.2 | 6 (35.3%) | 9 (52.9%) | 2 (11.8%) |
| Fillet (20) | 17 (85) | 2.8 | 4.4 | 3.6 ± 0.5 | 0 (0.0%) | 10 (58.8%) | 7 (41.2%) |
| Total (80) | 73 (91.3) | 0.48 | 4.8 | 3.1 ± 0.9 | 10 (13.7%) | 46 (63.0%) | 17 (23.3%) |
| Antimicrobial Agent | Resistant | Intermediate | Susceptible | |||
|---|---|---|---|---|---|---|
| No. | (%) | No. | (%) | No. | (%) | |
| Chloramphenicol (CHL) | 72 | 50.3 | 2 | 1.4 | 69 | 48.3 |
| Ampicillin/sulbactam (SAM) | 73 | 51.0 | 2 | 1.4 | 68 | 47.6 |
| Chloramphenicol (CAF) | 73 | 51.0 | 1 | 0.7 | 69 | 48.3 |
| Erythromycin (ERY) | 84 | 58.7 | 3 | 2.1 | 56 | 39.2 |
| Gentamicin (GEN) | 86 | 60.1 | 2 | 1.4 | 55 | 38.5 |
| Imipenem (IMP) | 95 | 66.4 | 1 | 0.7 | 47 | 32.9 |
| Fusidic acid (FA) | 61 | 42.7 | 2 | 1.4 | 80 | 55.9 |
| Cefotaxime (CTX) | 101 | 70.6 | 3 | 2.1 | 39 | 27.3 |
| Polymyxin B (PMB) | 94 | 65.7 | 1 | 0.7 | 48 | 33.6 |
| Tetracycline (TET) | 45 | 31.5 | 1 | 0.7 | 97 | 67.8 |
| Vancomycin (VAN) | 49 | 34.3 | 4 | 2.8 | 90 | 62.9 |
| Code | Antimicrobial Resistance Pattern | No of Strains | MAR Index |
|---|---|---|---|
| P1 | IMP, CTX, and PMB | 4 | 0.27 |
| P2 | CHL, CAF, CTX, and PMB | 15 | 0.36 |
| P3 | Y and SAM | 10 | 0.36 |
| P4 | Y, SAM, and VAN | 3 | 0.45 |
| P5 | Y, SAM, and PMB | 3 | 0.45 |
| P6 | CAF, ERY, IMP, CTX, and TET | 9 | 0.45 |
| P7 | Y, SAM, PMB, and VAN | 3 | 0.55 |
| P8 | X, CTX, TET, and VAN | 7 | 0.55 |
| P9 | Y, SAM, CTX, and PMB | 2 | 0.55 |
| P10 | Y, Z, and CHL | 12 | 0.64 |
| P11 | Y, Z, CHL, and VAN | 3 | 0.73 |
| P12 | X, Z, GEN, and IMP | 7 | 0.73 |
| P13 | Y, Z, SAM, TET, and VAN | 4 | 0.82 |
| P14 | Y, Z, SAM, CAF, and TET | 6 | 0.82 |
| P15 | Y, Z, CHL, CAF, TET, and VAN | 4 | 0.91 |
| P16 | X, Y, Z, and TET | 4 | 0.91 |
| P17 | X, Y, Z, and VAN | 2 | 0.91 |
| P18 | X, Y, FA, CTX, TET, and VAN | 4 | 0.91 |
| P19 | X, Y, FA, PMB, TET, and VAN | 3 | 0.91 |
| P20 | X, Z, ERY, IMP, TET, and VAN | 5 | 0.91 |
| P21 | X, Z, ERY, IMP, TET, and VAN | 2 | 0.91 |
| P22 | X, Y, Z, TET, and VAN | 5 | 1.00 |
| Strain Code | Closest Related Strain | Accession Number | Similarity |
|---|---|---|---|
| CPSA-05 | Staphylococcus aureus UP_1097 | CP047803 | 98.70 |
| CPSA-11 | Staphylococcus aureus Min-175 | CP086121 | 98.15 |
| CPSA-18 | Staphylococcus aureus AATYW | CP116909 | 99.13 |
| CPSA-29 | Staphylococcus aureus CHU15-080 | CP065871 | 98.80 |
| CPSA-34 | Staphylococcus aureus 1549-SCV | LT992435 | 98.60 |
| CPSA-47 | Staphylococcus aureus SA 1807 | CP041634 | 99.16 |
| Strain Code | Thymol | Carvacrol | Eugenol | Hydroquinone | Protocatechuic Acid |
|---|---|---|---|---|---|
| CPSA-5 | 600 ± 100 | 320 ± 0 | 1600 ± 400 | 100 ± 0 | 1200 ± 200 |
| CPSA-11 | 500 ± 100 | 400 ± 0 | 2400 ± 400 | 100 ± 0 | 800 ± 0 |
| CPSA-18 | 400 ± 0 | 600 ± 100 | 2400 ± 400 | 12.5 ± 0 | 1000 ± 300 |
| CPSA-29 | 600 ± 100 | 300 ± 0 | 1600 ± 0 | 50 ± 0 | 1200 ± 200 |
| CPSA-34 | 400 ± 100 | 300 ± 0 | 1600 ± 400 | 12.5 ± 0 | 600 ± 100 |
| CPSA-47 | 400 ± 100 | 400 ± 100 | 2400 ± 400 | 50 ± 0 | 800 ± 0 |
| Antimicrobial Group | Antimicrobial Agent | Concentration (µg/mL) | Abb. | Breakpoints | ||
|---|---|---|---|---|---|---|
| S | I | R | ||||
| Phenicol | Chloramphenicol | 30 | CHL | ≥18 | 13–17 | ≤12 |
| β-lactam | Ampicillin/sulbactam | 10/10 | SAM | ≥2 | - | ≤2 |
| Chloramphenicol | Chloramphenicol | 30 | CAF | ≥18 | 13–17 | ≤12 |
| Macrolides | Erythromycin | 15 | ERY | ≥23 | 14–22 | ≤13 |
| Aminoglycosides | Gentamicin | 10 | GEN | ≥15 | 13–14 | ≤12 |
| Carbapenem | Imipenem | 10 | IMP | ≥19 | 16–18 | ≤15 |
| Fusidane | Fusidic acid | 10 | FA | ≥22 | 20–21 | ≤19 |
| Cephalosporins | Cefotaxime | 30 | CTX | ≥26 | - | ≤2 |
| Polymixins | Polymyxin B | 300 | PMB | ≥12 | 9–11 | ≤8 |
| Tetracycline | Tetracycline | 30 | TET | ≥15 | 12–14 | ≤11 |
| Glycopeptide | Vancomycin | 30 | VAN | ≥17 | 15–16 | ≤14 |
| Target Gene | Sequence (5′-3′) | References |
|---|---|---|
| sea | F-GGTTATCAATGTGCGGGTGG | [50] |
| R-CGGCACTTTTTTCTCTTCGG | ||
| seb | F-GTATGGTGGTGTAACTGAGC | [50] |
| R-CCAAATAGTGACGAGTTAGG | ||
| clfa | F-ATTGGCGTGGCTTCAGTGCT | [51] |
| R-CGTTTCTTCCGTAGTTGCATTTG | ||
| tst | F-TTCACTATTTGTAAAAGTGTCAGACCCACT | [52] |
| R-TACTAATGAATTTTTTTATCGTAAGCCCTT | ||
| pvl | F-ATCATTAGGTAAAATGTCTGGACATGATCCA | [52] |
| R-GCATCAASTGTATTGGATAGCAAAAGC | ||
| aacA–aphD | F-TAATCC AAG AGC AAT AAG GGC | [53] |
| R-GCCACACTATCATAACCACTA | ||
| vanA | F-GGCAAGTCAGGTGAAGATG | [54] |
| R-ATCAAGCGGTCAATCAGTTC | ||
| mecA | F-AGAAGATGGTATGTGGAAGTTAG | [54] |
| R-ATGTATGTGCGATTGTATTGC | ||
| blaZ | F-ACTTCAACACCTGCTGCTTTC | [55] |
| R-TGACCACTTTTATCAGCAACC | ||
| apmA | F-CGTTTGCTTCGTGCATTAAA | [56] |
| R-TTGACACGAAGGAGGGTTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morshdy, A.E.M.A.; Abdallah, K.M.E.; Abdallah, H.E.; Algahtani, F.D.; Elabbasy, M.T.; Atique, S.; Ahmad, K.; Al-Najjar, M.A.A.; Abdallah, H.M.; Mahmoud, A.F.A. Potential of Natural Phenolic Compounds as Antimicrobial Agents against Multidrug-Resistant Staphylococcus aureus in Chicken Meat. Molecules 2023, 28, 6742. https://doi.org/10.3390/molecules28186742
Morshdy AEMA, Abdallah KME, Abdallah HE, Algahtani FD, Elabbasy MT, Atique S, Ahmad K, Al-Najjar MAA, Abdallah HM, Mahmoud AFA. Potential of Natural Phenolic Compounds as Antimicrobial Agents against Multidrug-Resistant Staphylococcus aureus in Chicken Meat. Molecules. 2023; 28(18):6742. https://doi.org/10.3390/molecules28186742
Chicago/Turabian StyleMorshdy, Alaa Eldin M. A., Karima M. E. Abdallah, Heba E. Abdallah, Fahad D. Algahtani, Mohamed Tharwat Elabbasy, Suleman Atique, Khursheed Ahmad, Mohammad A. A. Al-Najjar, Hossam M. Abdallah, and Abdallah Fikry A. Mahmoud. 2023. "Potential of Natural Phenolic Compounds as Antimicrobial Agents against Multidrug-Resistant Staphylococcus aureus in Chicken Meat" Molecules 28, no. 18: 6742. https://doi.org/10.3390/molecules28186742
APA StyleMorshdy, A. E. M. A., Abdallah, K. M. E., Abdallah, H. E., Algahtani, F. D., Elabbasy, M. T., Atique, S., Ahmad, K., Al-Najjar, M. A. A., Abdallah, H. M., & Mahmoud, A. F. A. (2023). Potential of Natural Phenolic Compounds as Antimicrobial Agents against Multidrug-Resistant Staphylococcus aureus in Chicken Meat. Molecules, 28(18), 6742. https://doi.org/10.3390/molecules28186742