



Therapeutic Study of Cinnamic Acid Derivative for Oxidative Stress Ablation: The Computational and Experimental Answers

 ,
,  ,
,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
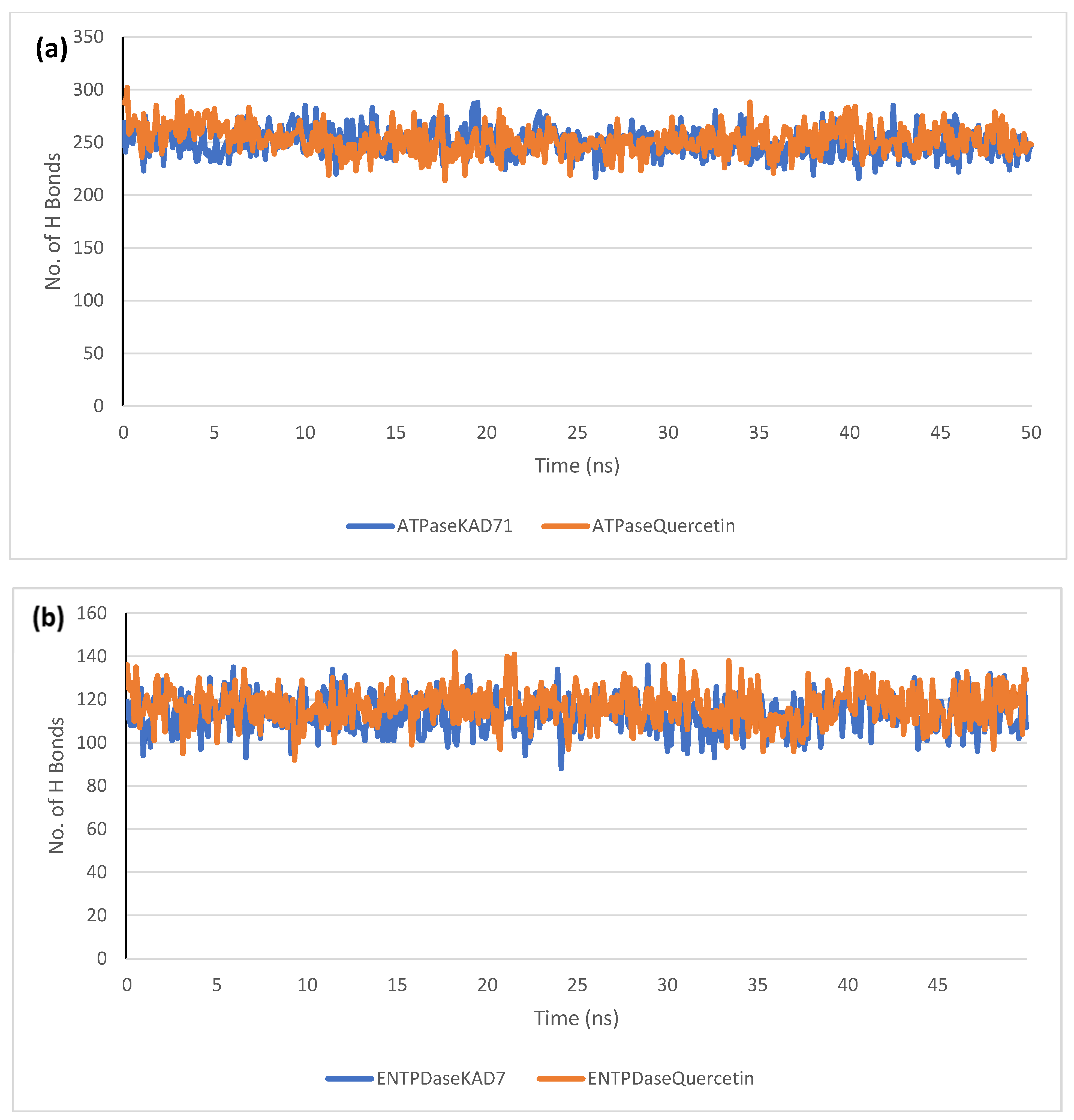
2. Results
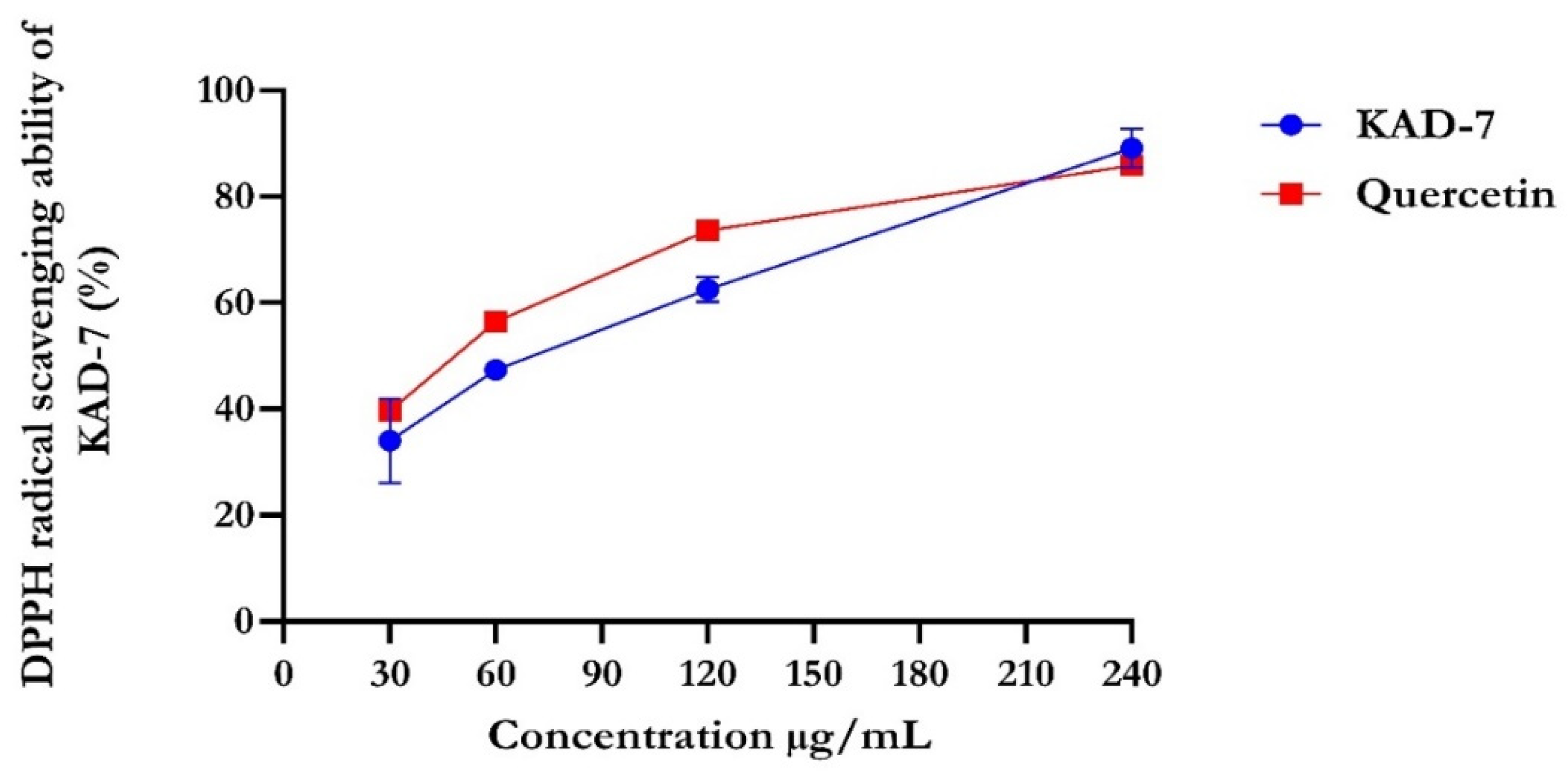
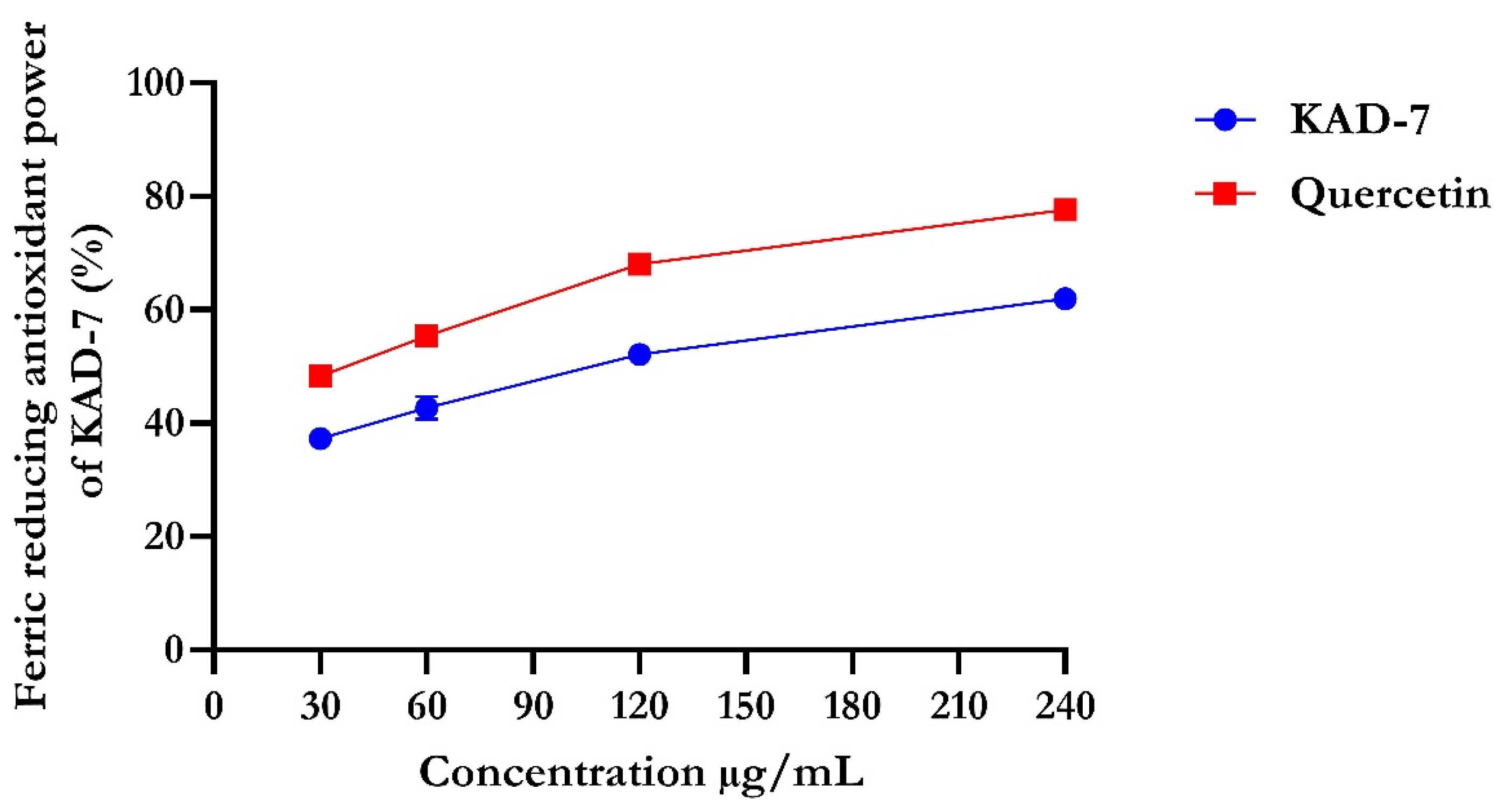
2.1. Antioxidant Activity
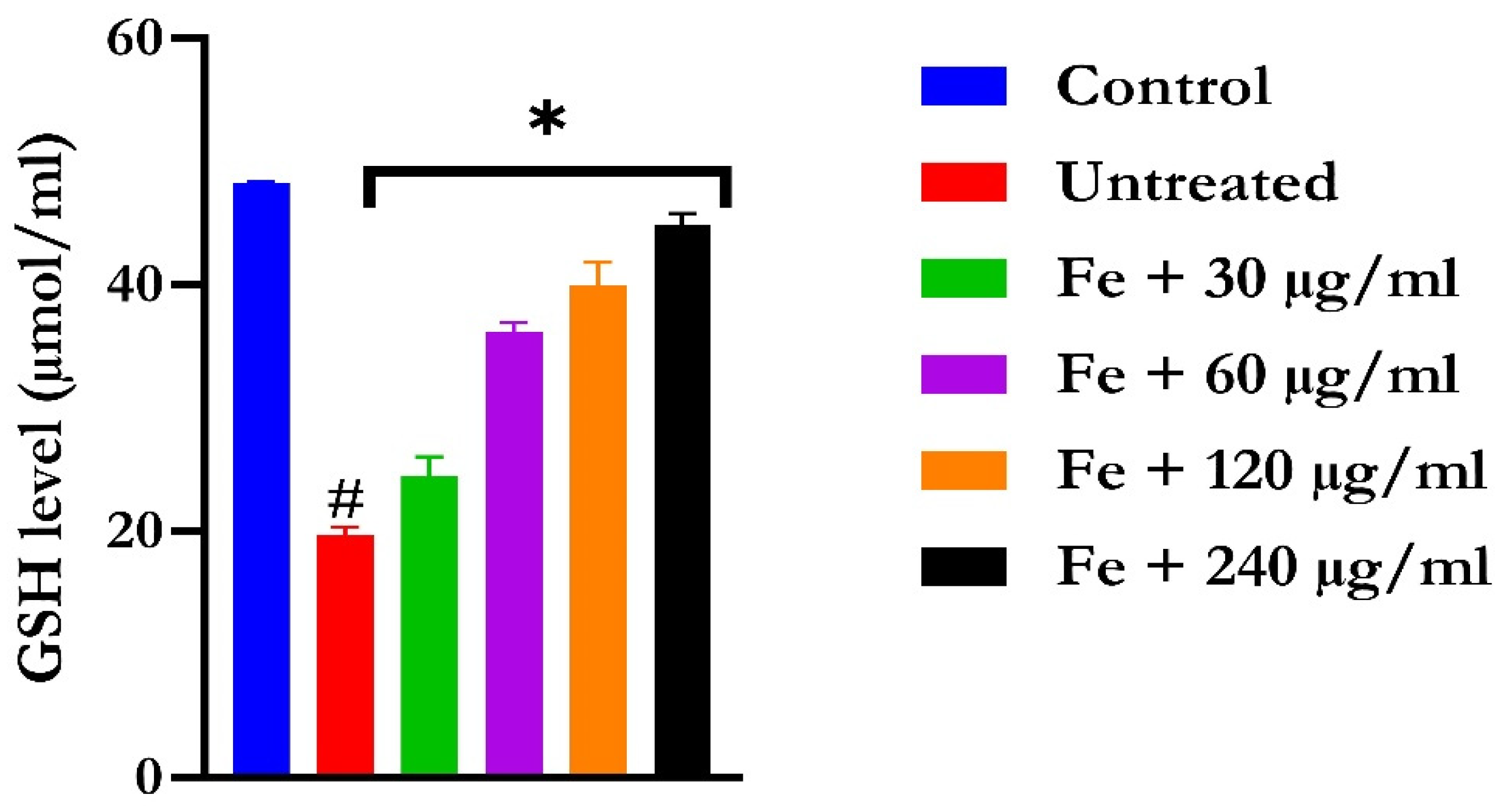
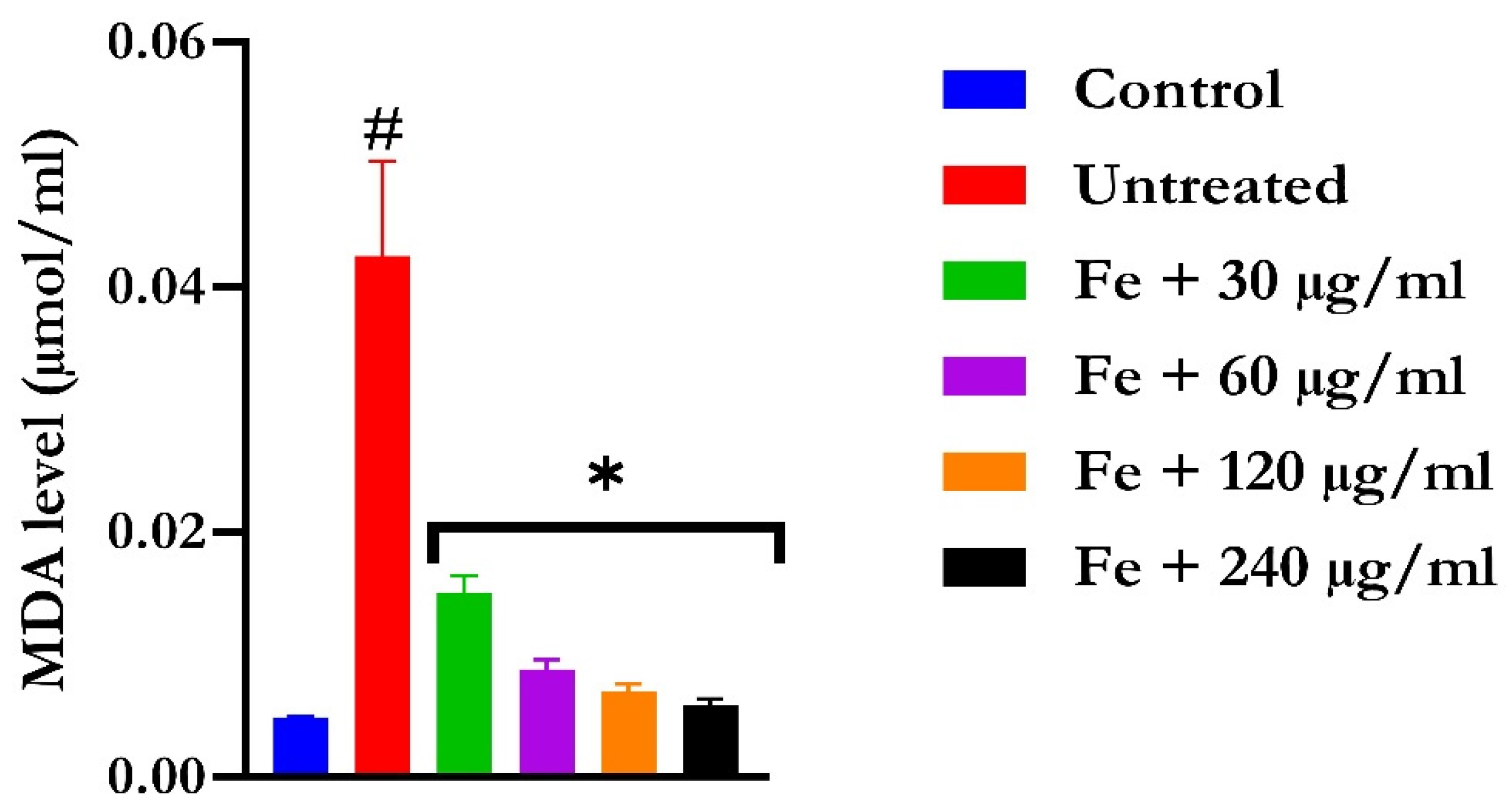
2.2. Ex Vivo Antioxidant Activity
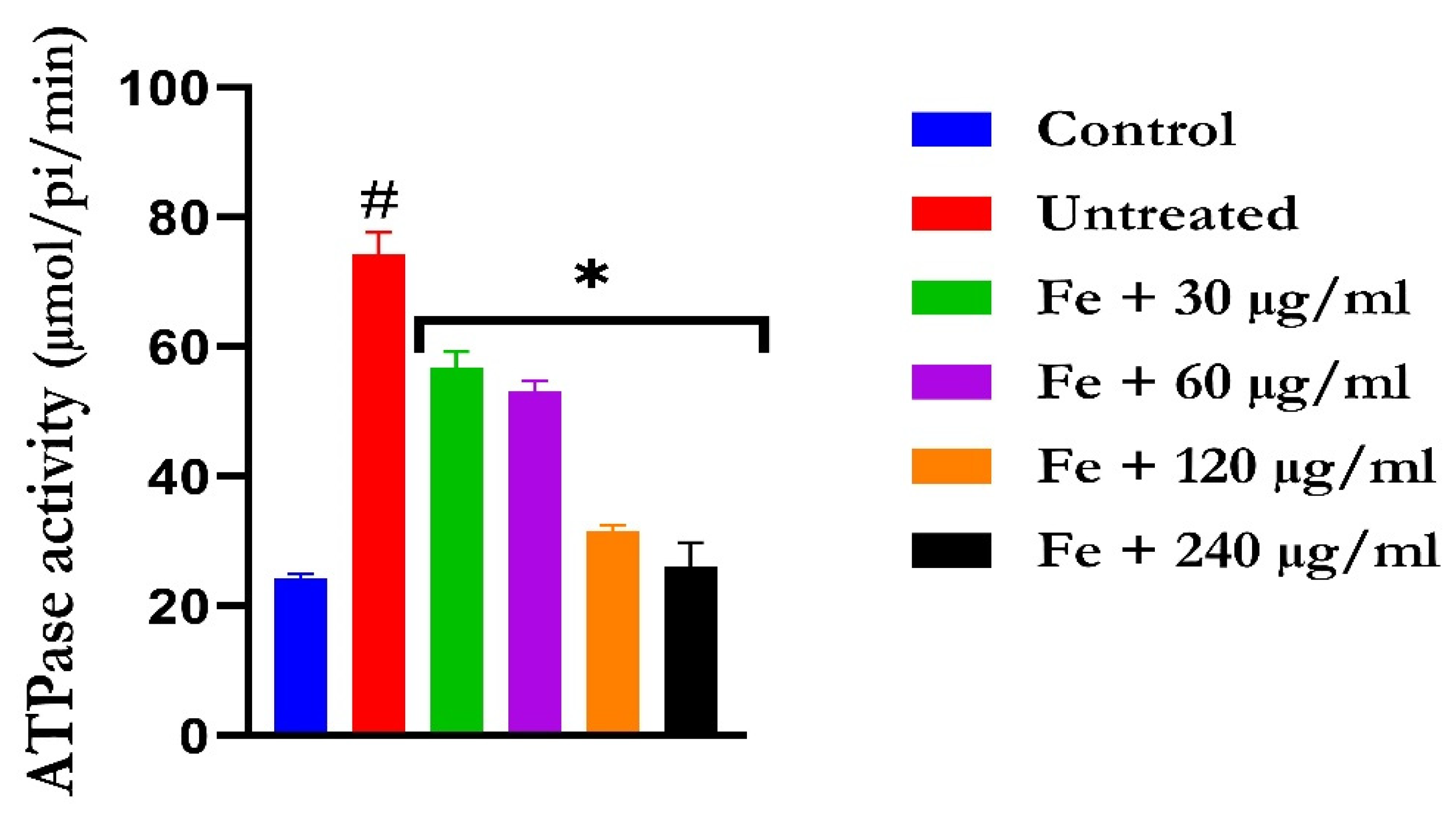
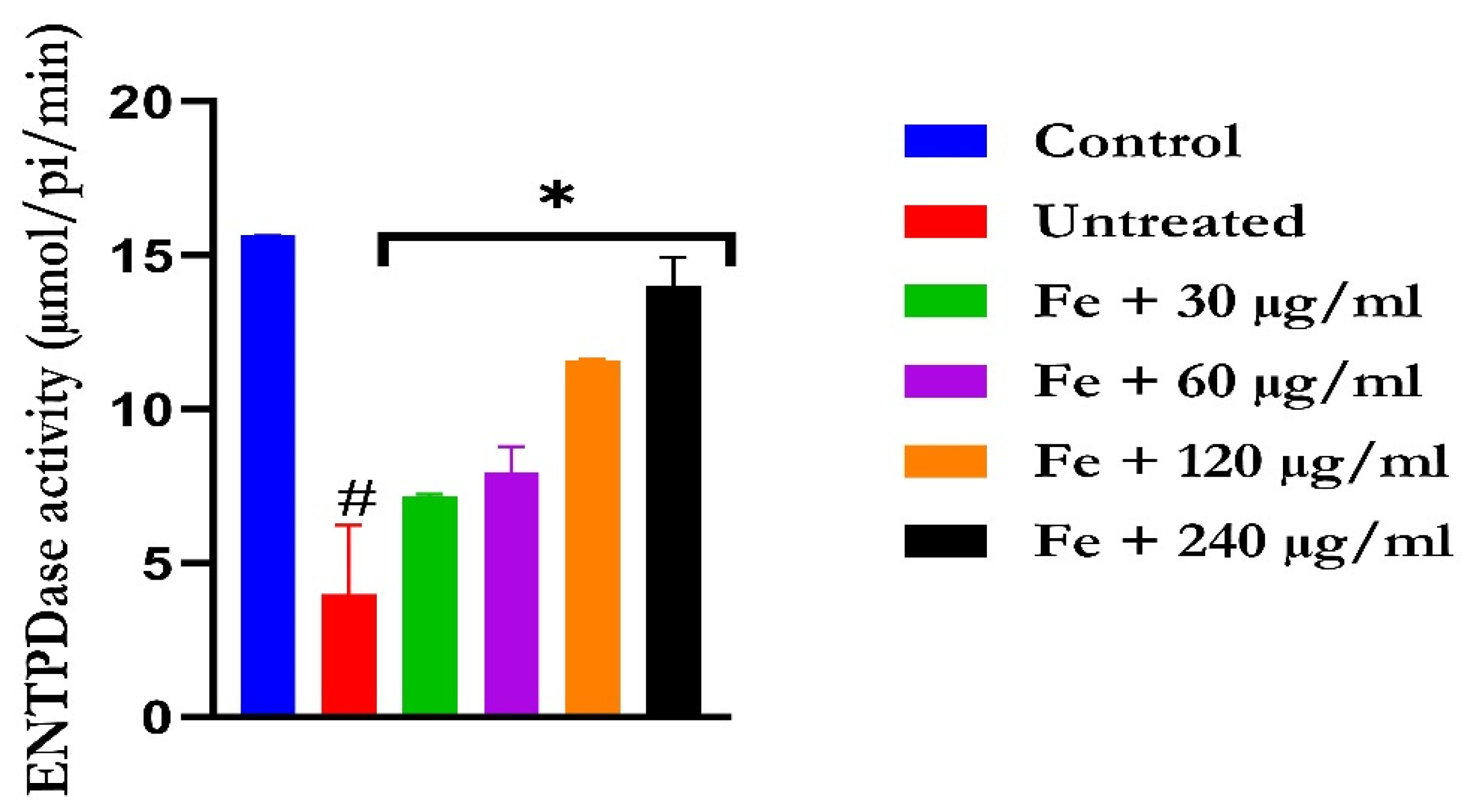
2.3. Purinergic Function
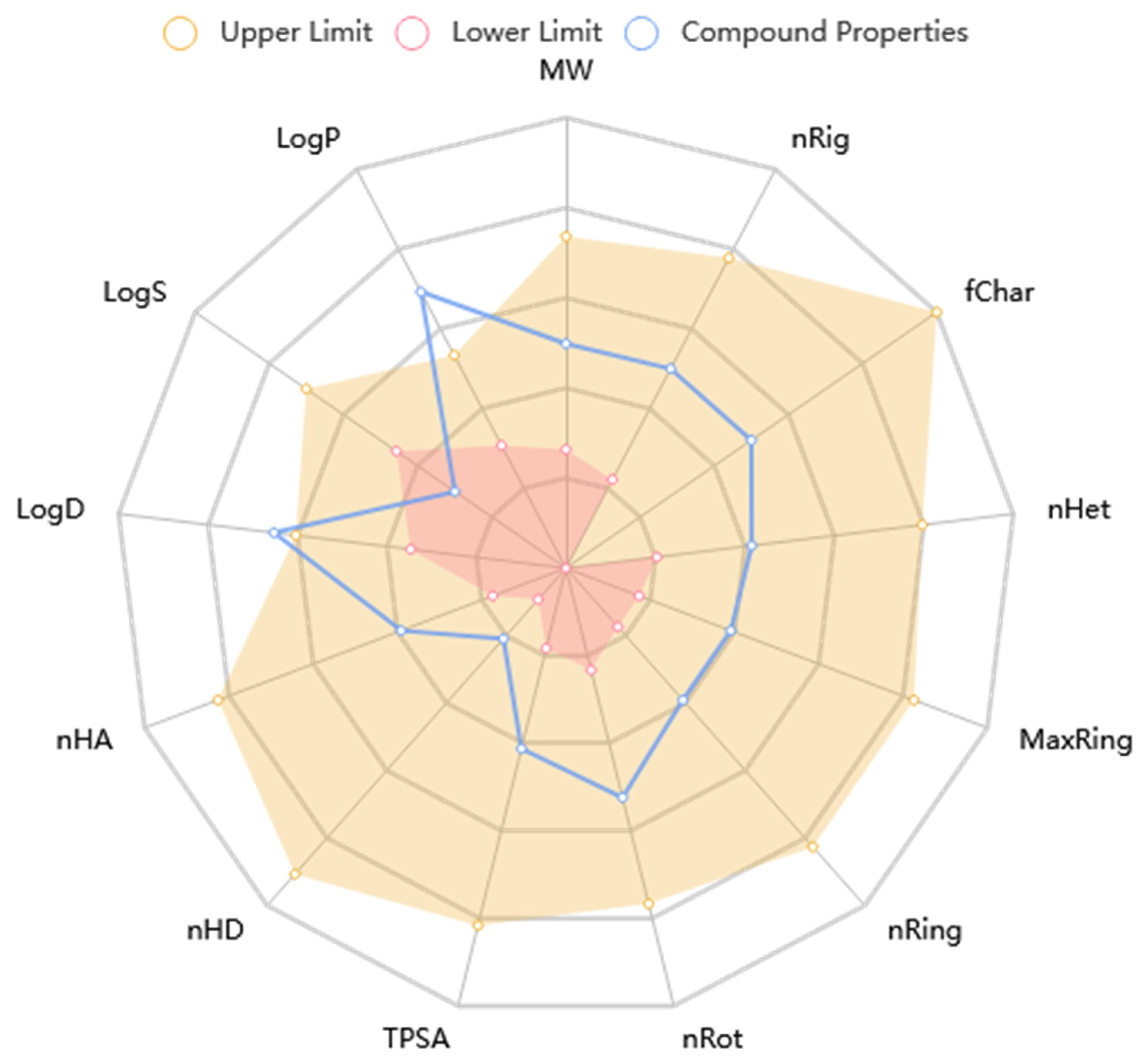
2.4. ADME/Toxicity Profiling of Ligands
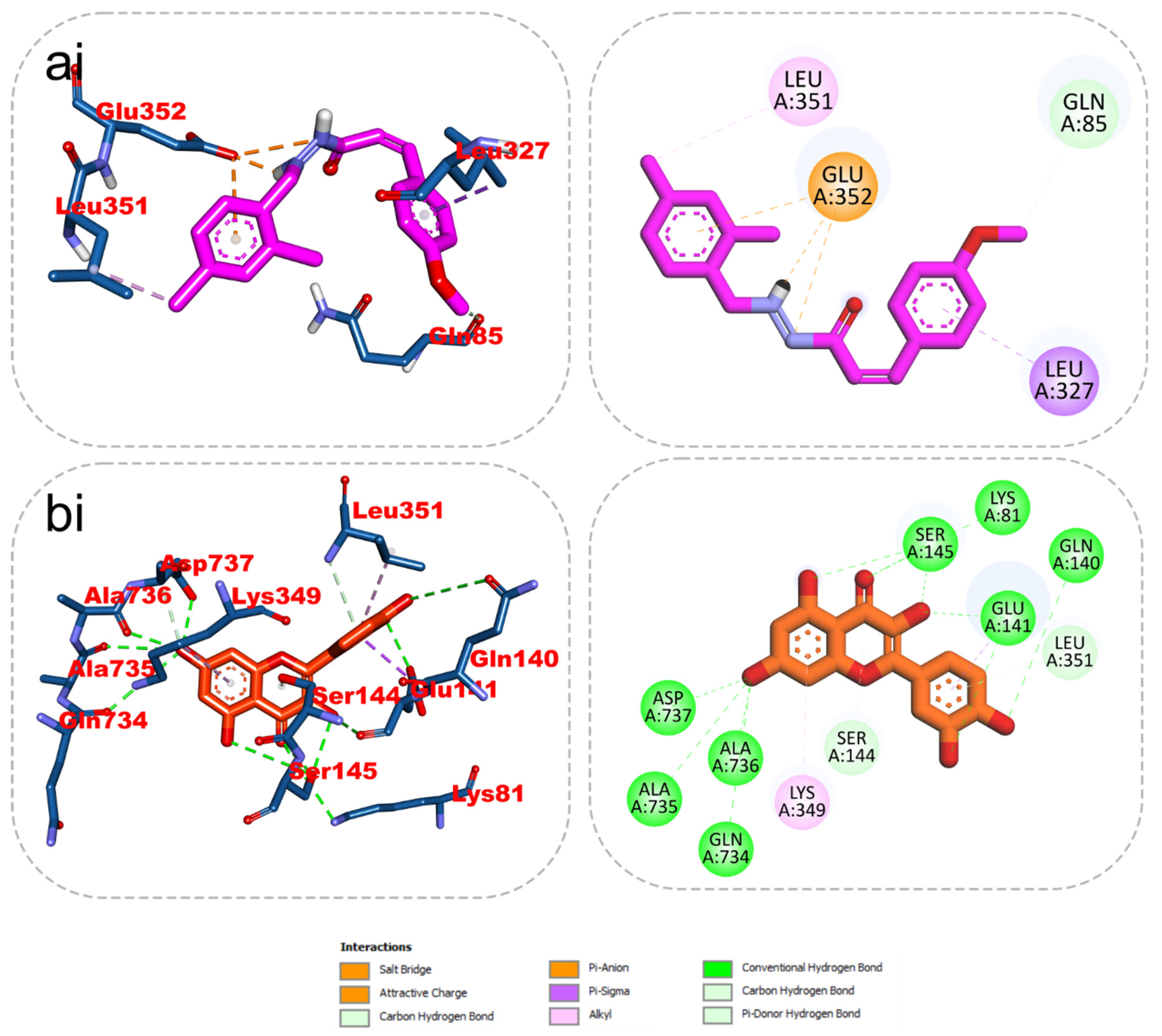
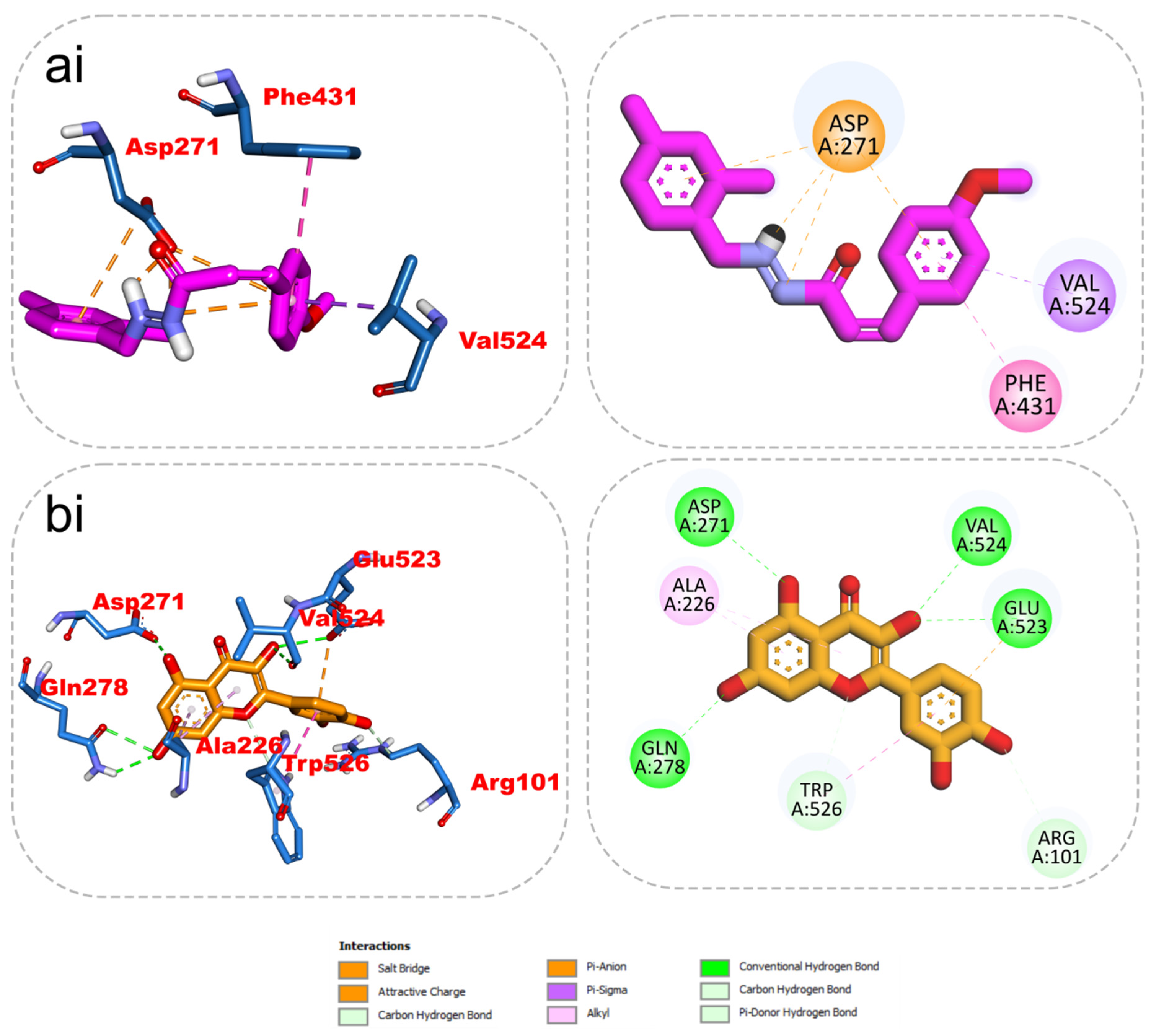
2.5. 2D/3D Interaction
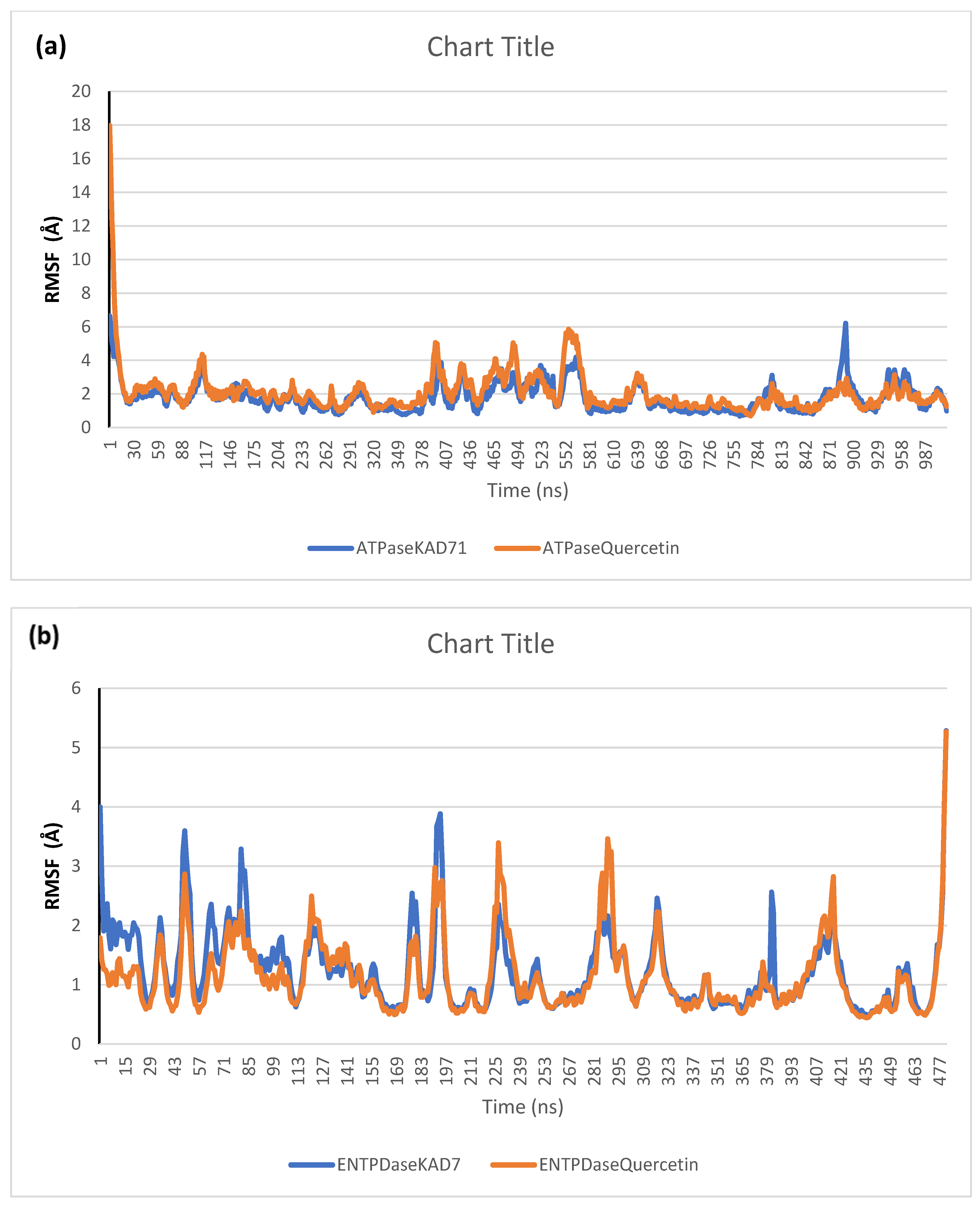
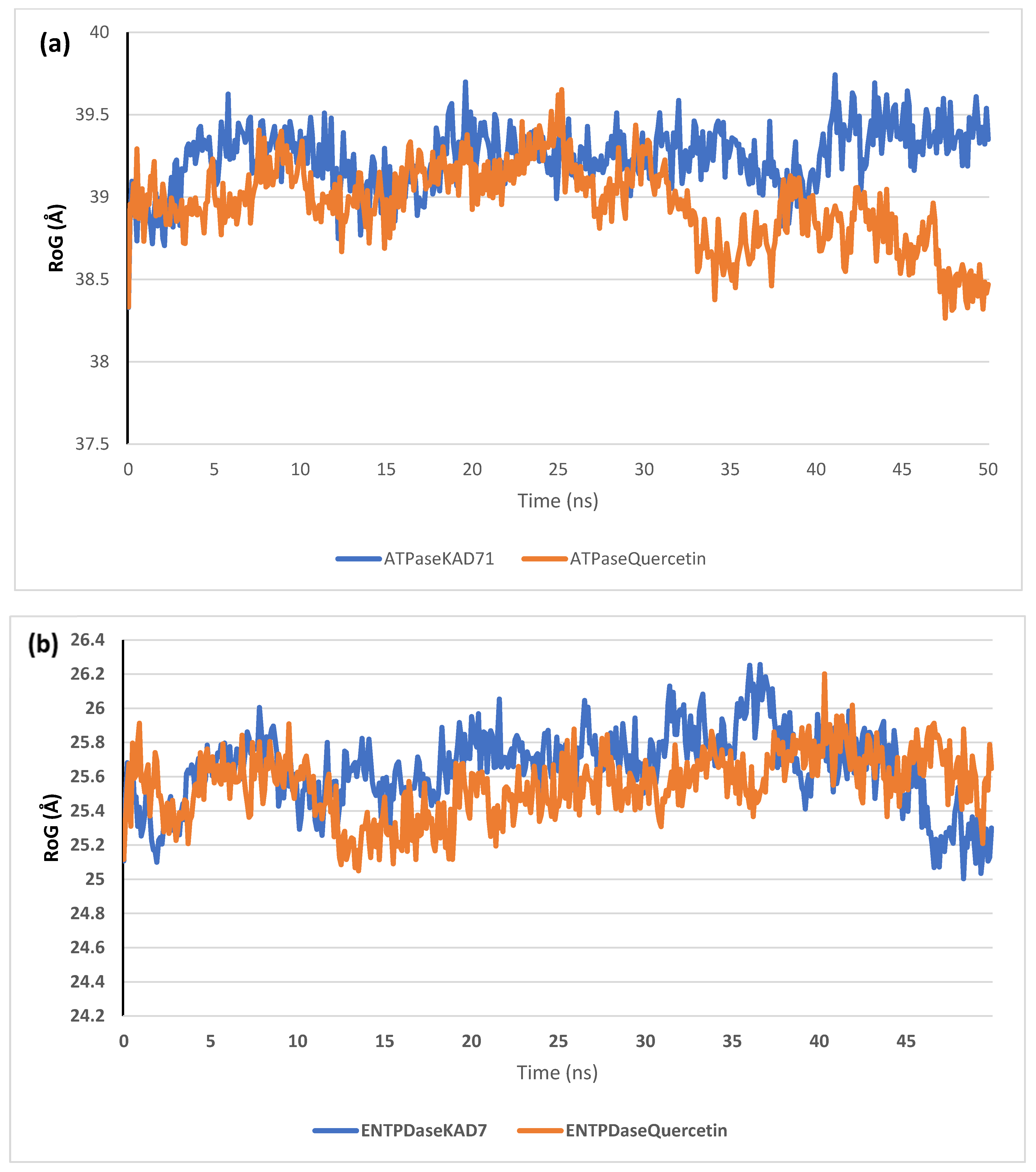
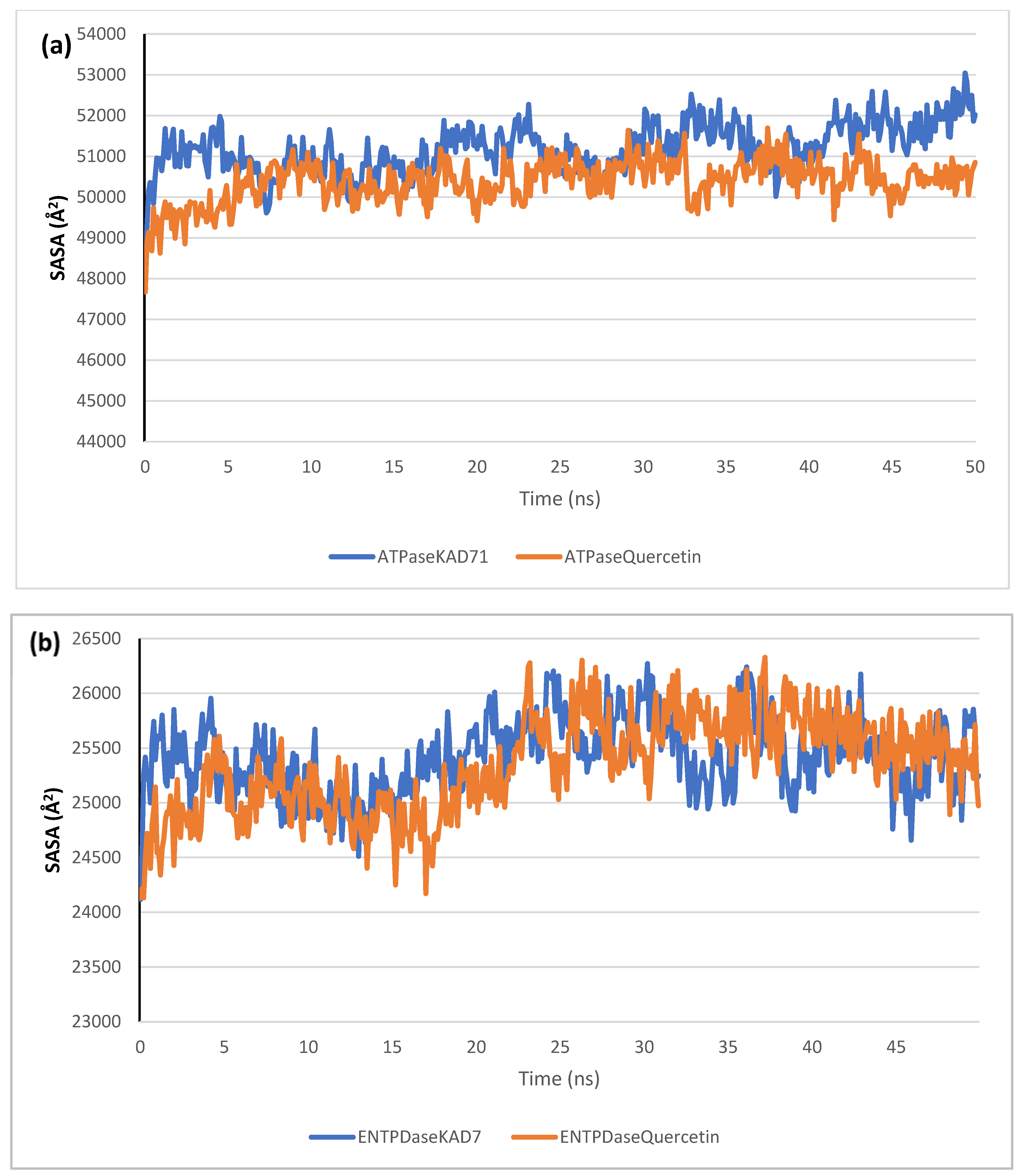
2.6. Molecular Dynamics Simulation
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Antioxidant Studies
4.2.1. DPPH Radical Scavenging Activity of KAD-7
4.2.2. Ferric-Reducing Antioxidant Power of KAD-7
4.2.3. Evaluation of Iron (Fe)-Chelating Activity of KAD-7
4.3. Experimental Rats and Organ Preparation
4.4. Induction of Liver Damage
4.5. Antioxidant Activities
4.5.1. Catalase (CAT) Activity of KAD-7
4.5.2. Reduced Glutathione Level
4.5.3. Lipid Peroxidation Level
4.6. Purinergic Activity
4.7. In Silico Studies
4.7.1. 3D Structure of Protein and Determination of Binding Pocket
4.7.2. Retrieval of N′-(2,4-Dichlorobenzylidene)-3-(4-methoxyphenyl) acrylohydrazide) and Quercetin
4.7.3. ADME/Toxicity Profiling of Ligands
4.7.4. Molecular Docking Simulation
4.7.5. Molecular Dynamics Simulation
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prakash, B.; Kumar, A.; Singh, P.P.; Songachan, L.J.F. Antimicrobial and antioxidant properties of phytochemicals: Current status and future perspective. Funct. Preserv. Prop. Phytochem. 2020, 1–45. [Google Scholar] [CrossRef]
- Ojo, O.A.; Oni, A.I.; Grant, S.; Amanze, J.; Ojo, A.B.; Taiwo, O.A.; Maimako, R.F.; Evbuomwan, I.O.; Iyobhebhe, M.; Nwonuma, C.O. Antidiabetic Activity of Elephant Grass (Cenchrus Purpureus (Schumach.) Morrone) via Activation of PI3K/AkT Signaling Pathway, Oxidative Stress Inhibition, and Apoptosis in Wistar Rats. Front. Pharmacol. 2022, 13, 845196. [Google Scholar] [CrossRef]
- Savych, A.; Marchyshyn, S.; Kyryliv, M.; Bekus, I. Cinnamic acid and its derivatives in the herbal mixtures and their antidiabetic activity. Farmacia 2021, 69, 595–601. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Z.; Pan, P.; Lao, Z.; Xu, J.; Li, Z.; Zhan, S.; Liu, X.; Wu, Y.; Wang, W.; et al. Cinnamic acid inhibits Zika virus by inhibiting RdRp activity. Antivir. Res. 2021, 192, 105117. [Google Scholar] [CrossRef] [PubMed]
- Ogunlakin, A.D.; Sonibare, M.A.; Yeye, O.E.; Jabeen, A.; Shah, S.F.; Ojo, O.A.; Gyebi, G.A.; Ayokunle, D.I. Design, Synthesis, and Characterization of Cinnamic Acid Derivatives with Two Novel Acrylohydrazones on HeLa and CHO-1 Cancer Cell Lines: The Experimental and Computational Perspective. Chem. Afr. 2023. [Google Scholar] [CrossRef]
- Rudrapal, M.; Satyanandam, R.S.; Swaroopini, T.; Lakshmi, T.N.; Jaha, S.K.; Zaheera, S. Synthesis and antibacterial activity of some new hydrazones. Med. Chem. Res. 2013, 22, 2840–2846. [Google Scholar] [CrossRef]
- Kumar, P.; Narasimhan, B. Hydrazides/hydrazones as antimicrobial and anticancer agents in the new millennium. Mini-Rev. Med. Chem. 2013, 13, 971–987. [Google Scholar] [CrossRef]
- Congiu, C.; Onnis, V. Synthesis and biological evaluation of novel acylhydrazone derivatives as potential antitumor agents. Bioorg. Med. Chem. 2013, 21, 6592–6599. [Google Scholar] [CrossRef]
- Patra, D.; Biswas, N.; Kumari, B.; Das, P.; Sepay, N.; Chatterjee, S.; Drew, M.G.; Ghosh, T. A family of mixed-ligand oxidovanadium (V) complexes with aroylhydrazone ligands: A combined experimental and computational study on the electronic effects of para substituents of hydrazone ligands on the electronic properties, DNA binding and nuclease activities. RSC Adv. 2015, 5, 92456–92472. [Google Scholar]
- Alagesan, M.; Bhuvanesh, N.S.; Dharmaraj, N. Potentially cytotoxic new copper (II) hydrazone complexes: Synthesis, crystal structure and biological properties. Dalton Trans. 2013, 42, 7210–7223. [Google Scholar] [PubMed]
- Nolasco-Quintana, N.Y.; Gonzalez-Maya, L.; Razo-Hernandez, R.S.; Alvarez, L. Exploring the gallic and cinnamic acids chimeric derivatives as Anticancer Agents over HeLa cell line: An in silico and in vitro study. Mol. Inf. 2023, 42, 2200016. [Google Scholar] [CrossRef] [PubMed]
- Şenkardeş, S.; Han, M.; Kulabaş, N.; Abbak, M.; Cevik, O.; Kucukguzel, İ.; Kucukguzel, Ş.G. Synthesis, molecular docking and evaluation of novel sulfonyl hydrazones as anticancer agents and COX-2 inhibitors. Mol. Divers. 2020, 24, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Nassar, I.F.; Farargy, A.F.; Abdelrazek, F.M.; Hamza, Z. Synthesis of new uracil derivatives and their sugar hydrazones with potent antimicrobial, antioxidant and anticancer activities. Nucleosides Nucleotides Nucleic Acids 2020, 39, 991–1010. [Google Scholar] [CrossRef]
- Ogunlakin, A.D.; Sonibare, M.A.; Jabeen, A.; Shah, S.F.; Shaheen, F. Antioxidant and anti-proliferative studies on Kigelia Africana (Lam.) Benth. And its constituents. Trop. J. Nat. Prod. Res. 2021, 5, 570–575. [Google Scholar]
- Moslemi, E.; Musazadeh, V.; Kavyani, Z.; Naghsh, N.; Shoura, S.M.S.; Dehghan, P. Efficacy of vitamin D supplementation as an adjunct therapy for improving inflammatory and oxidative stress biomarkers: An umbrella meta-analysis. Pharmacol. Res. 2022, 186, 106484. [Google Scholar] [CrossRef]
- Ahmad, S.; Alrouji, M.; Alhajlah, S.; Alomeir, O.; Pandey, R.P.; Ashraf, M.S.; Ahmad, S.; Khan, S. Secondary Metabolite Profiling, Antioxidant, Antidiabetic and Neuroprotective Activity of Cestrum nocturnum (Night Scented-Jasmine): Use of In Vitro and In Silico Approach in Determining the Potential Bioactive Compound. Plants 2023, 12, 1206. [Google Scholar] [CrossRef]
- Anderson, E.; Shah, Y. Iron homeostasis in the liver. Compr. Physiol. 2013, 3, 315–330. [Google Scholar] [PubMed]
- Erukainure, O.L.; Matsabisa, M.G.; Salau, V.F.; Oyedemi, S.O.; Oyenihi, O.R.; Ibeji, C.U.; Islam, M.S. Cannabis sativa L. (var. indica) Exhibits Hepatoprotective Effects by Modulating Hepatic Lipid Profile and Mitigating Gluconeogenesis and Cholinergic Dysfunction in Oxidative Hepatic Injury. Front. Pharmacol. 2021, 12, 705402. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Gan, R.-Y.; Li, S.; Zhou, Y.; Li, A.-N.; Xu, D.-P.; Li, H.-B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Chukwuma, C.I.; Matsabisa, M.G.; Javu, M.T.; Salau, V.F.; Koorbanally, N.A.; Islam, M.S. Turbina oblongata Protects against Oxidative Cardiotoxicity by Suppressing Lipid Dysmetabolism and Modulating Cardiometabolic Activities Linked to Cardiac Dysfunctions. Front. Pharmacol. 2021, 12, 610835. [Google Scholar] [CrossRef]
- Rychlicka, M.; Rot, A.; Gliszczyńska, A. Biological properties, health benefits and enzymatic modifications of dietary methoxylated derivatives of cinnamic acid. Foods 2021, 10, 1417. [Google Scholar] [CrossRef] [PubMed]
- Alevizopoulos, K.; Calogeropoulou, T.; Lang, F.; Stournaras, C. Na+/K+ ATPase inhibitors in cancer. Curr. Drug Targets 2014, 15, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H.; Zebisch, M.; Strater, N. Cellular Function and Molecular Structure of Ecto-Nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef]
- Afzal, S.; Al-Rashida, M.; Hameed, A.; Pelletier, J.; Sévigny, J.; Iqbal, J. Synthesis, In-Vitro Evaluation and Molecular Docking Studies of Oxoindolin Phenylhydrazine Carboxamides as Potent and Selective Inhibitors of Ectonucleoside Triphosphate Diphosphohydrolase (NTPDase). Bioorg. Chem. 2021, 112, 104957. [Google Scholar] [CrossRef] [PubMed]
- Srikanthan, K.; Shapiro, J.I.; Sodhi, K. The Role of Na/K-ATPase Signaling in Oxidative Stress Related to Obesity and Cardiovascular Disease. Molecules 2016, 21, 1172. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, J.-X.; Wu, X.-Y.; Cui, Y.; Zou, Z.-H.; Liu, Y.; Gao, J. Correlation Analysis of Plasma Myeloperoxidase Level with Global Registry of Acute Coronary Events Score and Prognosis in Patients with Acute Non-ST-Segment Elevation Myocardial Infarction. Front. Med. 2022, 9, 828174. [Google Scholar] [CrossRef]
- Liu, Q.; Kulak, M.V.; Borcherding, N.; Maina, P.K.; Zhang, W.; Weigel, R.J.; Qi, H.H. A novel HER2 gene body enhancer contributes to HER2 expression. Oncogene 2018, 37, 687–694. [Google Scholar] [CrossRef]
- Zhou, B.; Lin, W.; Long, Y.; Yang, Y.; Zhang, H.; Wu, K.; Chu, Q. Notch signaling pathway: Architecture, disease, and therapeutics. Sig. Transduct. Target Ther. 2022, 7, 95. [Google Scholar] [CrossRef]
- Al-Rashida, M.; Iqbal, J. Therapeutic Potentials of Ecto-Nucleoside Triphosphate Diphosphohydrolase, Ecto-Nucleotide Pyrophosphatase/Phosphodiesterase, Ecto-5’-Nucleotidase, and Alkaline Phosphatase Inhibitors. Med. Res. Rev. 2014, 34, 703–743. [Google Scholar] [CrossRef]
- Bashir, L.; Saidu, S.; Amos, S.O.; Yunusa, O.I.; Abdulhakeem, R.A.; Halimat, Y.L.; Femi, O.; Ali, A.J.; Batiha, G.E.L.-S.; Shukurat, B.B.; et al. Preclinical anti-inflammatory and antioxidant effects of Azanza garckeana in STZ-induced glycemic-impaired rats, and pharmacoinformatics of it major phytoconstituents. Biomed. Pharmacother. 2022, 152, 113196. [Google Scholar]
- Sliwoski, G.; Kothiwale, S.; Meiler, J.; Lowe, E.W. Computational methods in drug discovery. Pharmacol. Rev. 2014, 66, 334–395. [Google Scholar] [CrossRef]
- Lee, K.; Kim, D. In-silico molecular binding prediction for human drug targets using deep neural multi-task learning. Genes 2019, 10, 906. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ivanov, I. Molecular dynamics. In Computational Toxicology: Volume I; Humana Press: Totowa, NJ, USA, 2012; pp. 243–285. [Google Scholar]
- Dong, Y.-W.; Liao, M.-L.; Meng, X.-L.; Somero, G.N. Structural flexibility and protein adaptation to temperature: Molecular dynamics analysis of malate dehydrogenases of marine molluscs. Proc. Natl. Acad. Sci. USA 2018, 115, 1274–1279. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Wang, S.M. Classification of VUS and unclassified variants in BRCA1 BRCT repeats by molecular dynamics simulation. Comput. Struct. Biotechnol. J. 2020, 18, 723–736. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A boiled-egg to predict gastrointestinal absorption and brain penetration of small molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Drug-like properties and the causes of poor solubility and poor permeability. J. Pharmacol. Toxicol. Methods 2000, 44, 235–249. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.-Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Wang, N.-N.; Huang, C.; Dong, J.; Yao, Z.-J.; Zhu, M.-F.; Deng, Z.-K.; Lv, B.; Lu, A.-P.; Chen, A.F.; Cao, D.-S. Predicting human intestinal absorption with modified random forest approach: A comprehensive evaluation of molecular representation, unbalanced data, and applicability domain issues. RSC Adv. 2017, 7, 19007–19018. [Google Scholar]
- Kratz, J.M.; Grienke, U.; Scheel, O.; Mann, S.A.; Rollinger, J.M. Natural products modulating the hERG channel: Heartaches and hope. Nat. Prod. Rep. 2017, 34, 957–980. [Google Scholar] [CrossRef]
- Raschi, E.; Vasina, V.; Poluzzi, E.; De Ponti, F. The hERG K+ channel: Target and antitarget strategies in drug development. Pharmacol. Res. 2008, 57, 181–195. [Google Scholar] [CrossRef]
- Kalpana, A.; Vinodhini, T. In silico validation and comparison of antifungal competence and druglikeness of some natural xanthones–A step towards antimycotic therapeutics. J. Indian Chem. Soc. 2022, 99, 100577. [Google Scholar] [CrossRef]
- Sapkota, K.; Mao, Z.; Synowicki, P.; Lieber, D.; Liu, M.; Ikezu, T.; Gautam, V.; Monaghan, D.T. GluN2D N-methyl-d-aspartate receptor subunit contribution to the stimulation of brain activity and gamma oscillations by ketamine: Implications for schizophrenia. J. Pharmacol. Exp. Ther. 2016, 356, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Djoumbou Feunang, Y.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E. ClassyFire: Automated chemical classification with a comprehensive, computable taxonomy. J. Cheminform. 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Martin, T.M.; Ye, L.; Sedykh, A.; Young, D.M.; Tropsha, A. Quantitative structure− activity relationship modeling of rat acute toxicity by oral exposure. Chem. Res. Toxicol. 2009, 22, 1913–1921. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Cheng, F.; Chen, L.; Du, Z.; Li, W.; Liu, G.; Lee, P.W.; Tang, Y. In silico prediction of chemical Ames mutagenicity. J. Chem. Inf. Model. 2012, 52, 2840–2847. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Morales, P.; Cámara, M.; Alves, M.J.; Oliveira, M.B.P.; Ferreira, I.C. Wild Fragaria vesca L. fruits: A rich source of bioactive phytochemicals. Food Funct. 2016, 7, 4523–4532. [Google Scholar] [CrossRef]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Chukwuma, C.I.; Matsabisa, M.G.; Salau, V.F.; Koorbanally, N.A.; Islam, M.S. Buddleja saligna Willd (Loganiaceae) inhibits angiotensin-converting enzyme activity in oxidative cardiopathy with concomitant modulation of nucleotide hydrolyzing enzymatic activities and dysregulated lipid metabolic pathways. J. Ethnopharmacol. 2020, 248, 112358. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Islam, M. Caffeic acid protects against iron-induced cardiotoxicity by suppressing angiotensin-converting enzyme activity and modulating lipid spectrum, gluconeogenesis and nucleotide hydrolyzing enzyme activities. Biol. Trace Elem. Res. 2021, 199, 1052–1061. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Mopuri, R.; Oyebode, O.A.; Koorbanally, N.A.; Islam, M.S. Dacryodes edulis enhances antioxidant activities, suppresses DNA fragmentation in oxidative pancreatic and hepatic injuries; and inhibits carbohydrate digestive enzymes linked to type 2 diabetes. Biomed. Pharmacother. 2017, 96, 37–47. [Google Scholar] [CrossRef]
- Kozakov, D.; Grove, L.E.; Hall, D.R.; Bohnuud, T.; Mottarella, S.E.; Luo, L.; Xia, B.; Beglov, D.; Vajda, S. FTMap family of web servers for determining and characterizing ligand-binding hot spots of proteins. Nat. Protoc. 2015, 10, 733–755. [Google Scholar] [CrossRef] [PubMed]
- Schöning-Stierand, K.; Diedrich, K.; Fährrolfes, R.; Flachsenberg, F.; Meyder, A.; Nittinger, E.; Steinegger, R.; Rarey, M. ProteinsPlus: Interactive analysis of protein–ligand binding interfaces. Nucleic Acids Res. 2020, 48, W48–W53. [Google Scholar] [CrossRef] [PubMed]
- Garbett, N.C.; Chaires, J.B. Thermodynamic studies for drug design and screening. Expet. Opin. Drug Discov. 2012, 7, 299–314. [Google Scholar] [CrossRef]
- Adewole, K.E.; Gyebi, G.A.; Ibrahim, I.M. Amyloid β fibrils disruption by kolaviron: Molecular docking and extended molecular dynamics simulation studies. Comput. Biol. Chem. 2021, 94, 107557. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.A.; Ogunlakin, A.D.; Iyobhebhe, M.; Olowosoke, C.B.; Taiwo, O.A.; Akinola, A.; Fadiora, D.; Odugbemi, A.I.; Gyebi, G.A.; Nwonuma, C.O. Computer aided and experimental study of cinnamic acid analog for oxidative stress treatment: The therapeutic validations. Inform. Med. Unlocked 2022, 35, 101137. [Google Scholar] [CrossRef]
- Gyebi, G.A.; Ogunyemi, O.M.; Ibrahim, I.M.; Afolabi, S.O.; Adebayo, J.O. Dual targeting of cytokine storm and viral replication in COVID-19 by plant-derived steroidal pregnanes: An in silico perspective. Comput. Biol. Med. 2021, 134, 104406. [Google Scholar] [CrossRef] [PubMed]
- Ogunyemi, O.M.; Gyebi, G.A.; Ibrahim, I.M.; Olaiya, C.O.; Ocheje, J.O.; Fabusiwa, M.M.; Adebayo, J.O. Dietary stigmastane-type saponins as promising dual-target directed inhibitors of SARS-CoV-2 proteases: A structure-based screening. RSC Adv. 2021, 11, 33380–33398. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L.; Mackerell, A.D., 3rd; Nilsson, L., Jr.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef]
- Jo, S.; Cheng, X.; Islam, S.M.; Huang, L.; Rui, H.; Zhu, A.; Lee, H.S.; Qi, Y.; Han, W.; Vanommeslaeghe, K. CHARMM-GUI PDB manipulator for advanced modeling and simulations of proteins containing nonstandard residues. Adv. Protein Chem. Struct. Biol. 2014, 96, 235–265. [Google Scholar]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for charmm SUNHWAN. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Govindarajan, N.; Tiwari, A.; Ensing, B.; Meijer, E.J. Impact of the ligand flexibility and solvent on the O–O bond formation step in a highly active ruthenium water oxidation catalyst. Inorg. Chem. 2018, 57, 13063–13066. [Google Scholar] [CrossRef] [PubMed]
- Hess, B.; Bekker, H.; Berendsen, H.J.; Fraaije, J.G. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds/ ADMET-Profile | Properties | KAD-7 N′-(2,4-Dichlorobenzylidene)-3-(4-methoxyphenyl)acrylohydrazide N′-(2,4-Dichlorobenzylidene)-3-(4-methoxyphenyl)acrylohydrazide) |
|---|---|---|
| Physicochemical | Formula | C17H14Cl2N2O2 |
| Molecular weight (g/mol) | 349.21 | |
| H-bond acceptors | 3 | |
| H-bond donors | 1 | |
| Rotatable bond | 6 | |
| TPSA (Å2) | 50.69 | |
| AlogP | 4.17 | |
| Water solubility (LogS) | −4.325 | |
| Absorption | XLOGP3 | 4.48 |
| WLOGP | 4.06 | |
| SILICOS-IT | 4.87 | |
| Human intestinal absorption | +0.9927 | |
| Caco2 | +0.6696 | |
| Human oral bioavailability | +0.5857 | |
| P-gp inhibitor | −0.6796 | |
| P-gp substrate | −0.8773 | |
| Distribution | Volume distribution (L/Kg) | 0.668 |
| Plasma protein binding (100%) | 93.8 | |
| Blood–brain barrier | +0.9804 | |
| Metabolism | OATP2B1 inhibitor | −0.8521 |
| OATP1B1 inhibitor | +0.9248 | |
| OATP1B3 inhibitor | +0.9501 | |
| MATE1 inhibitor | −0.9400 | |
| OCT2 inhibitor | −0.6000 | |
| BSEP inhibitor | +0.9097 | |
| CYP3A4 substrate | +0.5923 | |
| CYP2C9 substrate | −0.6150 | |
| CYP2D6 substrate | −0.8428 | |
| CYP3A4 inhibition | +0.5625 | |
| CYP2C9 inhibition | +0.8115 | |
| CYP2C19 inhibition | +0.9337 | |
| CYP2D6 inhibition | −0.8913 | |
| CYP1A2 inhibition | +0.9318 | |
| Excretion | Clearance | 4.116 |
| Half-life | 0.112 | |
| Toxicity | Carcinogenicity (binary) | −0.6718 |
| Hepatotoxicity | +0.7250 | |
| Respiratory toxicity | −0.6333 | |
| Reproductive toxicity | −0.5778 | |
| Mitochondrial toxicity | −0.6750 | |
| Nephrotoxicity | +0.4681 | |
| Acute oral toxicity (c) | III 0.7012 | |
| Ames mutagenesis | −0.5200 | |
| Drug-likeness | Lipinski violation | Nil |
| Ghose violation | Nil | |
| Veber violation | Nil | |
| Egan violation | Nil | |
| Medicinal Chemistry | PAIN violation | 0 |
| BRENK violation | 2 | |
| Lead-likeness | No (XLOGP3 > 3.5) |
| Compounds | Protein | Hydrogen Bonds Bond Distance (Å) | Hydrophobic Interaction | |
|---|---|---|---|---|
| Binding Energies | Interacted Residues | Interacted Residues | ||
| KAD-7 | ATPase | −7.1 | Leu351, Glu352, Gln85, Leu327 | |
| Quercetin | −7.9 | Asp737, Ala735, Ala736, Gln734, Ser145, Lys81, Glu141, Gln140 | Lys349, Glu141 | |
| KAD-7 | ENTPDase | −7.4 | Asp271, Phe431, Val524 | |
| Quercetin | −7.8 | Asp271, Val524, Glu523, Gln278 | Ala226, Trp526, Glu523 | |
| RMSD | RMSF | RoG | SASA | H-Bonds | |
|---|---|---|---|---|---|
| Mean (Å) | Mean (Å) | Mean (Å) | Mean (Å) | Mean (Å) | |
| ATPase_KAD-7 | 4.48 ± 0.18 | 1.80 ± 0.83 | 39.24 ± 0.18 | 51,187.90 ± 598.62 | 250.07 ± 13.04 |
| ATPase_Quercetin | 4.19 ± 0.31 | 2.09 ± 1.26 | 38.95 ± 0.24 | 50,378.98 ± 514.39 | 252.57 ± 13.68 |
| ENTPDase_KAD-7 | 2.19 ± 0.64 | 1.23 ± 0.64 | 25.64 ± 0.22 | 25,420.13 ± 347.37 | 113.45 ± 8.23 |
| ENTPDase_Quercetin | 2.00 ± 0.29 | 1.14 ± 0.61 | 25.54 ± 0.18 | 25,341.01 ± 432.89 | 116.78 ± 8.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojo, O.A.; Ogunlakin, A.D.; Maimako, R.F.; Gyebi, G.A.; Olowosoke, C.B.; Taiwo, O.A.; Elebiyo, T.C.; Adeniyi, D.; David, B.; Iyobhebhe, M.; et al. Therapeutic Study of Cinnamic Acid Derivative for Oxidative Stress Ablation: The Computational and Experimental Answers. Molecules 2023, 28, 7425. https://doi.org/10.3390/molecules28217425
Ojo OA, Ogunlakin AD, Maimako RF, Gyebi GA, Olowosoke CB, Taiwo OA, Elebiyo TC, Adeniyi D, David B, Iyobhebhe M, et al. Therapeutic Study of Cinnamic Acid Derivative for Oxidative Stress Ablation: The Computational and Experimental Answers. Molecules. 2023; 28(21):7425. https://doi.org/10.3390/molecules28217425
Chicago/Turabian StyleOjo, Oluwafemi Adeleke, Akingbolabo Daniel Ogunlakin, Rotdelmwa Filibis Maimako, Gideon Ampoma Gyebi, Christopher Busayo Olowosoke, Odunayo Anthonia Taiwo, Tobiloba Christiana Elebiyo, David Adeniyi, Bolaji David, Matthew Iyobhebhe, and et al. 2023. "Therapeutic Study of Cinnamic Acid Derivative for Oxidative Stress Ablation: The Computational and Experimental Answers" Molecules 28, no. 21: 7425. https://doi.org/10.3390/molecules28217425
APA StyleOjo, O. A., Ogunlakin, A. D., Maimako, R. F., Gyebi, G. A., Olowosoke, C. B., Taiwo, O. A., Elebiyo, T. C., Adeniyi, D., David, B., Iyobhebhe, M., Adetunji, J. B., Ayokunle, D. I., Ojo, A. B., Mothana, R. A., & Alanzi, A. R. (2023). Therapeutic Study of Cinnamic Acid Derivative for Oxidative Stress Ablation: The Computational and Experimental Answers. Molecules, 28(21), 7425. https://doi.org/10.3390/molecules28217425