

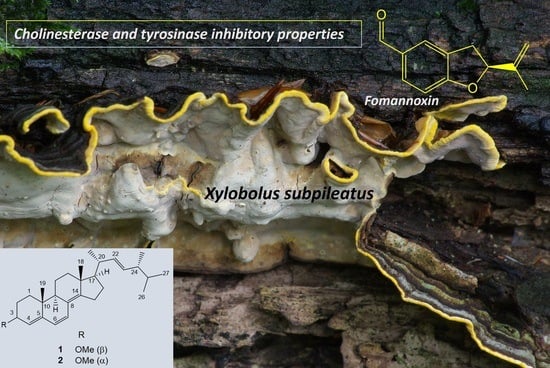
Anticholinesterase and Antityrosinase Secondary Metabolites from the Fungus Xylobolus subpileatus


 , , , , ,
, , , , ,  ,
,  and
and
Abstract
:
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Mushroom Material
3.3. Extraction and Isolation
3.4. Anti-Tyrosinase Activity
3.5. Acetylcholinesterase (AChE) and Butyrylcholinesterase (BChE) Inhibitory Activity
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, Y.-F.; He, S.-H. Xylobolus austrosinensis sp. nov. (Stereaceae, Russulales) and notes on the genus. Phytotaxa 2020, 452, 200–208. [Google Scholar] [CrossRef]
- Taudiere, A.; Bellanger, J.-M.; Moreau, P.-A.; Carcaillet, C.; Christophe, A.; Læssøe, T.; Panaïotis, C.; Richard, F. Xylobolus subpileatus, a specialized basidiomycete functionally linked to old canopy gaps. Can. J. For. Res. 2017, 47, 965–973. [Google Scholar] [CrossRef]
- Tura, D.; Zmitrovich, I.; Wasser, S.P.; Nevo, E. The genus Stereum in Israel. Mycotaxon 2008, 106, 109–126. [Google Scholar]
- Ryvarden, L. A note on the genus Xylobolus P. Karst. Synop. Fungorum 2012, 30, 17–24. [Google Scholar]
- Jahn, H.T. Stereoide Pilze in Europa (Stereaceae Pil. Emend. Parm. u. a., Hymenochaete), Mit Besonderer Berücksichtigung ihres Vorkommens in der Bundesrepublik Deutschland. In Westfälische Pilzbriefe; Pilzkundlichen Arbeitsgemeinschaft in Westfalen: Detmold, Germany, 1971; Volume 8, pp. 69–176. [Google Scholar]
- Chamuris, G.P. The non-stipitate steroid fungi in the northeastern United States and adjacent Canada. In Mycologia Memoirs; Schweizerbart Science Publishers: Stuttgart, Germany, 1988; Volume 14, 247p. [Google Scholar]
- Berkeley, M.J.; Curtis, M.A. Decades of fungi. Decades XXIII and XXIV. North and South Carolina Fungi. Hooker’s J. Bot. Kew Gard. Misc. 1849, 1, 234–239. [Google Scholar]
- Bernicchia, A.; Gorjón, S. Polypores of the Mediterranean Region; Romar: Segrate, Italy, 2020. [Google Scholar]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Yu, C.J.; Zhou, L.W. Species diversity and utilization of medicinal mushrooms and fungi in China (review). Int. J. Med. Mushrooms 2009, 11, 287–302. [Google Scholar] [CrossRef]
- Yasin, H.; Zahoor, M.; Yousaf, Z.; Aftab, A.; Saleh, N.; Riaz, N.; Shamsheer, B. Ethnopharmacological exploration of medicinal mushroom from Pakistan. Phytomedicine 2019, 54, 43–55. [Google Scholar] [CrossRef]
- Birkinshaw, J.H.; Chaplen, P.; Findlay, W.P. Biochemistry of the wood-rotting fungi. IX. Volatile metabolic products of Stereum subpileatum Berk. & Curt. Biochem. J. 1957, 66, 188–192. [Google Scholar] [CrossRef]
- Bu’Lock, J.D.; Kaye, B.; Hudson, A.T. New benzofurans from stereum subpileatum, their biosynthesis, and related processes of aromatic aminoacid metabolism in a basidiomycete. Phytochemistry 1971, 10, 1037–1046. [Google Scholar] [CrossRef]
- Tian, M.Q.; Wu, Q.L.; Wang, X.; Zhang, K.Q.; Li, G.H. A new compound from Stereum insigne CGMCC5.57. Nat. Prod. Res. 2017, 31, 932–937. [Google Scholar] [CrossRef]
- Lee, S.; Kim, C.S.; Yu, J.S.; Kang, H.; Yoo, M.J.; Youn, U.J.; Ryoo, R.; Bae, H.Y.; Kim, K.H. Ergopyrone, a styrylpyrone-fused steroid with a hexacyclic 6/5/6/6/6/5 Skeleton from a mushroom Gymnopilus orientispectabilis. Org. Lett. 2021, 23, 3315–3319. [Google Scholar] [CrossRef]
- Mahé, C.; Patin, H.; Van Hulle, M.T.; Barton, D.H.R. Structure and chemistry of π-allyl palladium complexes from steroids. J. Chem. Soc. Perkin Trans. 1 1981, 2504–2508. [Google Scholar] [CrossRef]
- Chobot, V.; Opletal, L.; Jahodář, L.; Patel, A.V.; Dacke, C.G.; Blunden, G. Ergosta-4,6,8,22-tetraen-3-one from the edible fungus, Pleurotus ostreatus (oyster fungus). Phytochemistry 1997, 45, 1669–1671. [Google Scholar] [CrossRef]
- Tanemossu, S.A.F.; Franke, K.; Arnold, N.; Schmidt, J.; Wabo, H.K.; Tane, P.; Wessjohann, L.A. Rare biscoumarin derivatives and flavonoids from Hypericum riparium. Phytochemistry 2014, 105, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T.H.J.; Lindequist, U.; Mentel, R.; Gördes, D.; Schmidt, E.; Thurow, K.; Lalk, M. Antiviral terpenoid constituents of Ganoderma pfeifferi. J. Nat. Prod. 2005, 68, 1728–1731. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.K.; Kuo, Y.H.; Chiang, B.H.; Lo, J.M.; Sheen, L.Y. Cytotoxic activities of 9,11-dehydroergosteroi peroxide and ergosterol peroxide from the fermentation mycelia of Ganoderma lucidum cultivated in the medium containing leguminous plants on Hep 3B cells. J. Agric. Food Chem. 2009, 57, 5713–5719. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Ko, Y.; Pang, C.; Ko, Y.J.; Choi, Y.K.; Kim, K.H.; Kang, K.S. Estrogenic Activity of Mycoestrogen (3β,5α,22E)-Ergost-22-en-3-ol via Estrogen Receptor α-Dependent Signaling Pathways in MCF-7 Cells. Molecules 2022, 27, 36. [Google Scholar] [CrossRef]
- Hansson, D.; Menkis, A.; Olson, K.; Stenlid, J.; Broberg, A.; Karlsson, M. Biosynthesis of fomannoxin in the root rotting pathogen Heterobasidion occidentale. Phytochemistry 2012, 84, 31–39. [Google Scholar] [CrossRef]
- Hirotani, M.; O’Reilly, J.; Donnelly, D.M.X.; Polonsky, J. Fomannoxin—A toxic metabolite of fomes annosus. Tetrahedron Lett. 1977, 18, 651–652. [Google Scholar] [CrossRef]
- Chang, J.C.; Hsiao, G.; Lin, R.K.; Kuo, Y.H.; Ju, Y.M.; Lee, T.H. Bioactive Constituents from the Termite Nest-Derived Medicinal Fungus Xylaria nigripes. J. Nat. Prod. 2017, 80, 38–44. [Google Scholar] [CrossRef]
- Arnone, A.; Assante, G.; Nasini, G.; Pava, O. ChemInform Abstract: Secondary Mould Metabolites. Part 38. Isolation and Structure Elucidation of Dihydrobenzofurans Isolated from Laurilia taxodii. Cheminform 2010, 23, 261. [Google Scholar] [CrossRef]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Giménez, J.; Cuadros, T.; Bové, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepulveda, M.; Peñuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 973. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in Parkinson’s disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef] [PubMed]
- González-Ramírez, M.; Gavilán, J.; Silva-Grecchi, T.; Cajas-Madriaga, D.; Triviño, S.; Becerra, J.; Saez-Orellana, F.; Pérez, C.; Fuentealba, J. A Natural Benzofuran from the Patagonic Aleurodiscus vitellinus Fungus has Potent Neuroprotective Properties on a Cellular Model of Amyloid-β Peptide Toxicity. J. Alzheimers Dis. 2018, 61, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.Y.; Lim, Y.; Yule, C. Evaluation of antioxidant, antibacterial and anti-tyrosinase activities of four Macaranga species. Food Chem. 2009, 114, 594–599. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Zalewski, P.; Szwajgier, D.; Baranowska-Wójcik, E.; Żarowski, M.; Plech, T.; Cielecka-Piontek, J. Permeability of Hypogymnia physodes Extract Component-Physodic Acid through the Blood-Brain Barrier as an Important Argument for Its Anticancer and Neuroprotective Activity within the Central Nervous System. Cancers 2021, 13, 1717. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Studzińska-Sroka, E.; Majchrzak-Celińska, A.; Bańdurska, M.; Rosiak, N.; Szwajgier, D.; Baranowska-Wójcik, E.; Szymański, M.; Gruszka, W.; Cielecka-Piontek, J. Is Caperatic Acid the Only Compound Responsible for Activity of Lichen Platismatia glauca within the Nervous System? Antioxidants 2022, 11, 2069. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | ||||
|---|---|---|---|---|---|---|
| δ 13C | δ 1H | m, J | δ 13C | δ 1H | m, J | |
| 1 | 33.4 | 1.73 | m | 29.7 | 1.53 | m |
| 1.38 | m | 1.26 | m | |||
| 2 | 25.0 | 2.09 | m | 23.6 | 1.96 | m |
| 1.62 | m | 1.76 | m | |||
| 3 | 76.6 | 3.93 | ddd, 10.1, 6.5, 2.0 Hz | 72.5 | 3.73 | b t, 4.6 Hz |
| 4 | 123.1 | 5.49 | b s | 120.9 | 5.59 | d, 4.6 Hz |
| 5 | 145.5 | - | - | 147.4 | - | - |
| 6 | 125.8 | 5.88 | d, 9.6 Hz | 125.9 | 5.88 | d, 9.7 Hz |
| 7 | 126.0 | 6.17 | d, 9.6 Hz | 126.8 | 6.21 | d, 9.7 Hz |
| 8 | 124.8 | - | - | 124.9 | - | - |
| 9 | 45.5 | 1.97 | m | 44.9 | 2.05 | m |
| 10 | 35.9 | - | - | 36.0 | - | - |
| 11 | 19.1 | 1.60 | m | 19.4 | 1.64 | m |
| 1.52 | m | 1.52 | m | |||
| 12 | 36.3 | 2.02 | m | 36.4 | 2.00 | m |
| 1.26 | m | 1.25 | m | |||
| 13 | 43.5 | - | - | 43.6 | - | - |
| 14 | 149.7 | - | - | 149.7 | - | - |
| 15 | 25.0 | 2.40 | m | 25.0 | 2.40 | m |
| 2.29 | m | 2.28 | m | |||
| 16 | 27.9 | 1.77 | m | 27.9 | 1.77 | m |
| 1.44 | m | 1.44 | m | |||
| 17 | 55.9 | 1.21 | m | 55.8 | 1.22 | m |
| 18 | 19.2 | 0.92 | s | 19.1 | 0.92 | s |
| 19 | 18.2 | 0.89 | s | 17.1 | 0.82 | s |
| 20 | 39.4 | 2.11 | m | 39.4 | 2.11 | m |
| 21 | 21.2 | 1.04 | d, 6.7 Hz | 21.2 | 1.04 | d, 6.8 Hz |
| 22 | 135.3 | 5.21 | m | 135.4 | 5.22 | m |
| 23 | 132.2 | 5.22 | m | 132.1 | 5.22 | m |
| 24 | 42.8 | 1.86 | m | 42.8 | 1.87 | m |
| 25 | 33.1 | 1.48 | m | 33.1 | 1.47 | m |
| 26 | 19.7 | 0.83 | d, 6.8 Hz | 19.7 | 0.83 | d, 6.8 Hz |
| 27 | 20.0 | 0.84 | d, 6.8 Hz | 20.0 | 0.84 | d, 6.8 Hz |
| 28 | 17.6 | 0.92 | d, 6.8 Hz | 17.6 | 0.93 | d, 6.8 Hz |
| 3-OCH3 | 55.4 | 3.40 | s | 56.1 | 3.36 | s |
| Compound/ Extract (8 mg/mL) | Inhibition (%) | SD |
|---|---|---|
| Methanol extract | 25.41 c/d | 7.08 |
| Hexane extract | 15.51 d | 2.88 |
| Chloroform extract | 38.33 c | 1.16 |
| 2 | na | - |
| 3 | 51.62 b | 11.80 |
| 4 | na | - |
| 5 | na | - |
| 6 | na | - |
| 7 | na | - |
| Azelaic acid (2 mg/mL) * | 91.63 a | 0.58 |
| Compound/ Extract (20 mg/mL) | AChE Inhibition (%) | SD | BChE Inhibition (%) | SD |
|---|---|---|---|---|
| Methanol extract | 31.67 d | 0.54 | 45.93 b | 3.24 |
| Hexane extract | na | - | 22.18 c/d | 3.57 |
| Chloroform extract | 94.05 a | 6.44 | 86.75 a | 9.07 |
| 2 | na | - | na | - |
| 3 | 67.66 c | 5.92 | 83.86 a | 9.79 |
| 4 | 31.28 d | 5.08 | 32.99 c | 14.08 |
| 5 | na | - | 6.98 e/f | 1.24 |
| 6 | na | - | 18.28 d/e | 1.38 |
| 7 | na | - | 0.49 f | 1.67 |
| Galanthamine (0.2 mg/mL) * | 76.21 b | 1.57 | 58.38 b | 7.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Felegyi, K.; Garádi, Z.; Studzińska-Sroka, E.; Papp, V.; Boldizsár, I.; Dancsó, A.; Béni, S.; Zalewski, P.; Ványolós, A. Anticholinesterase and Antityrosinase Secondary Metabolites from the Fungus Xylobolus subpileatus. Molecules 2024, 29, 213. https://doi.org/10.3390/molecules29010213
Felegyi K, Garádi Z, Studzińska-Sroka E, Papp V, Boldizsár I, Dancsó A, Béni S, Zalewski P, Ványolós A. Anticholinesterase and Antityrosinase Secondary Metabolites from the Fungus Xylobolus subpileatus. Molecules. 2024; 29(1):213. https://doi.org/10.3390/molecules29010213
Chicago/Turabian StyleFelegyi, Kristóf, Zsófia Garádi, Elżbieta Studzińska-Sroka, Viktor Papp, Imre Boldizsár, András Dancsó, Szabolcs Béni, Przemysław Zalewski, and Attila Ványolós. 2024. "Anticholinesterase and Antityrosinase Secondary Metabolites from the Fungus Xylobolus subpileatus" Molecules 29, no. 1: 213. https://doi.org/10.3390/molecules29010213
APA StyleFelegyi, K., Garádi, Z., Studzińska-Sroka, E., Papp, V., Boldizsár, I., Dancsó, A., Béni, S., Zalewski, P., & Ványolós, A. (2024). Anticholinesterase and Antityrosinase Secondary Metabolites from the Fungus Xylobolus subpileatus. Molecules, 29(1), 213. https://doi.org/10.3390/molecules29010213