
Transformation of Ginsenosides by Lactiplantibacillus plantarum MB11 Fermentation: Minor Ginsenosides Conversion and Enhancement of Anti-Colorectal Cancer Activity
 , , , , , , and
, , , , , , and
Abstract
:
1. Introduction
2. Results
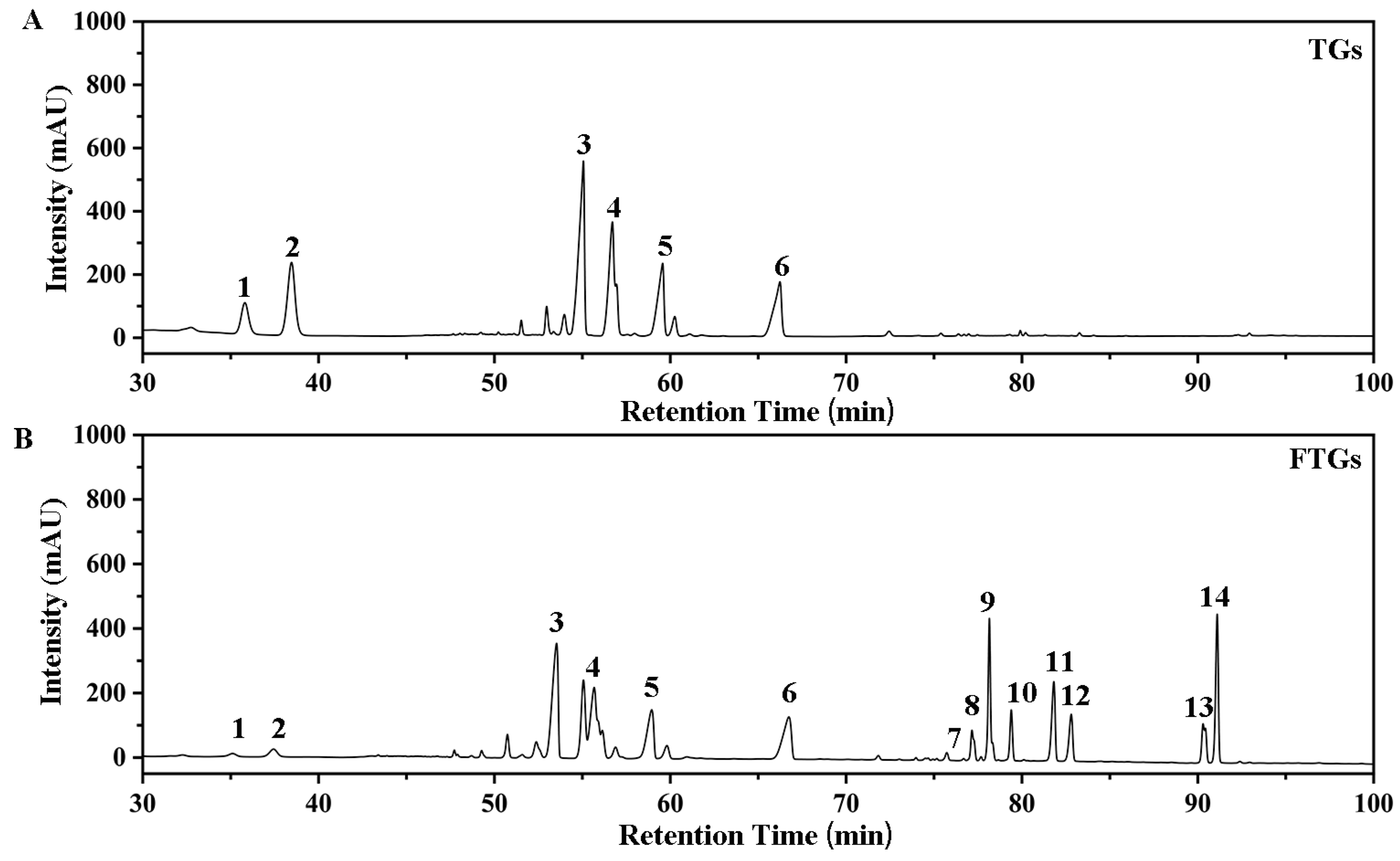
2.1. Transformation of Ginsenosides by L. plantarum MB11 Fermentation
2.2. FTGs Inhibit Tumor Growth in CT26-Bearing Mice
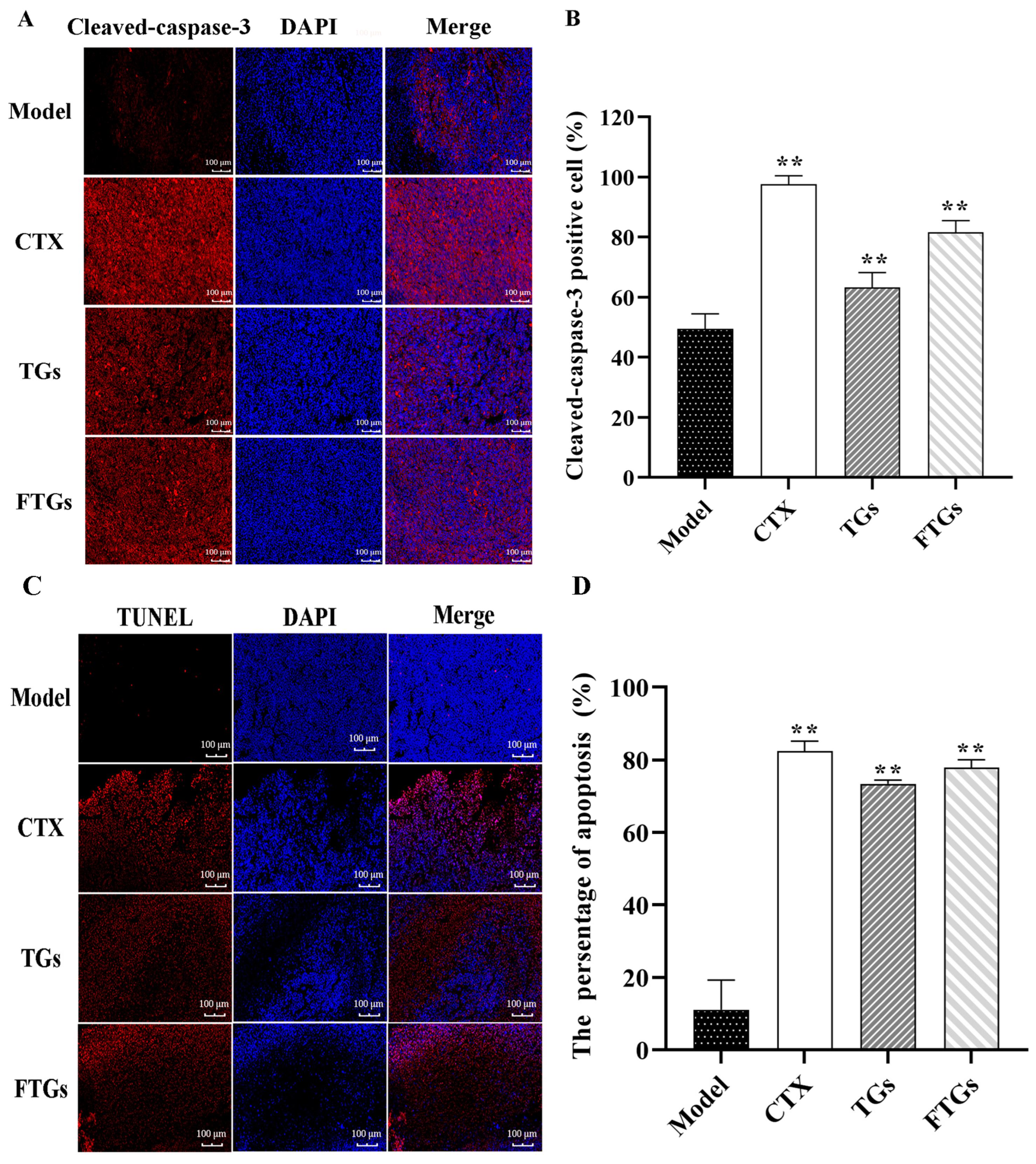
2.3. Effect of FTGs on the Expression of Caspase-3 in Tumor Tissue
2.4. In Vivo Apoptosis Induction by FTGs CT26-Bearing Mice
2.5. Effect of FTGs on AMPK/mTOR Signaling Pathway in CT26-Bearing Mice
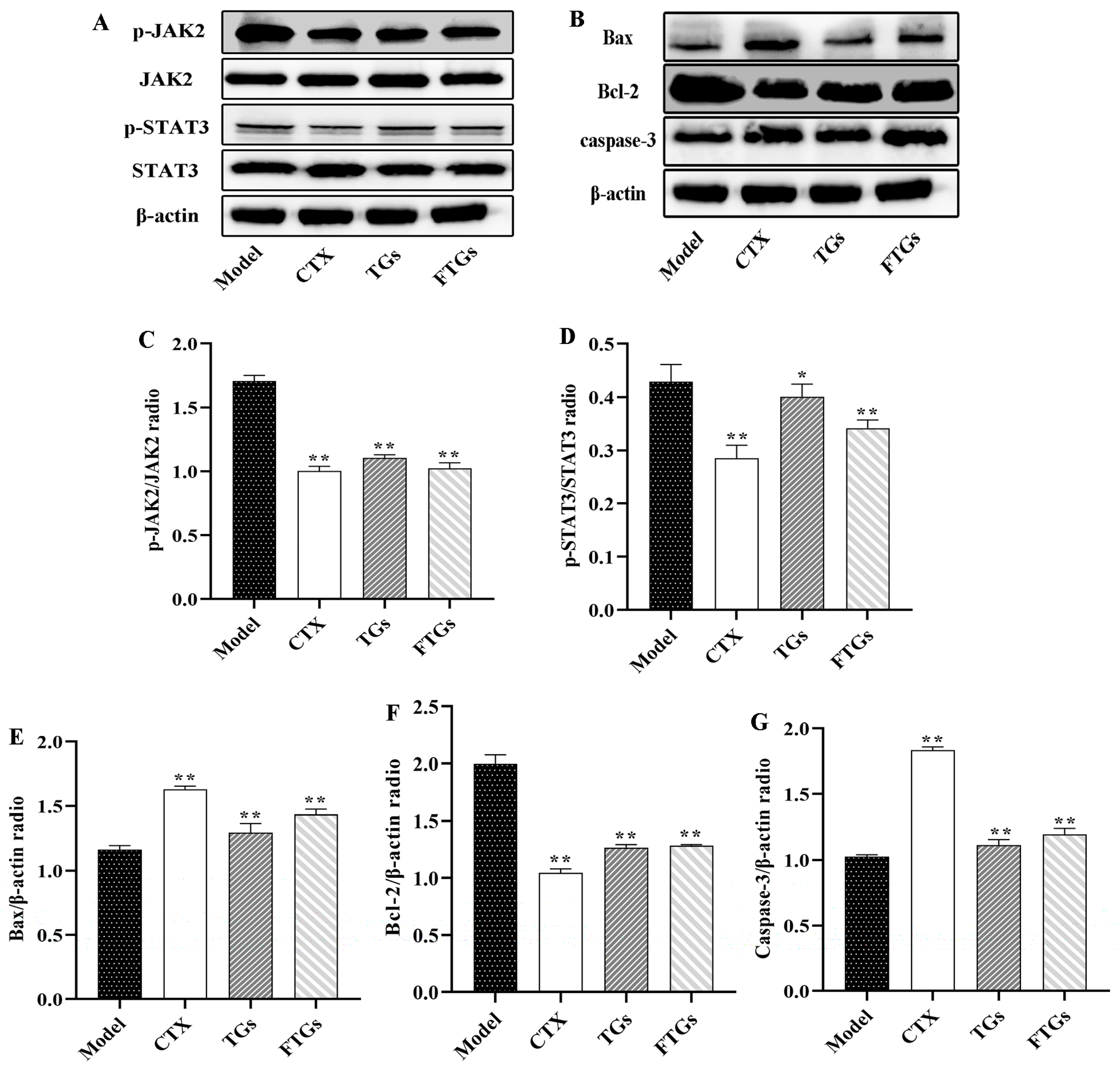
2.6. FTGs Exert Anticancer Activity via Inhibition of the JAK2/STAT3 Signaling Pathway
2.7. Effects of FTGs Treatment on Caspase-3, Bax, and Bcl-2expression in CT26-Bearing Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Lactiplantibacillus Strain and Culture Conditions
4.3. Fermentation of Total Ginsenoside by L. plantarum MB11
4.4. High-Performance Liquid Chromatography (HPLC) Analysis
4.5. Cell Line and Cell Culture
4.6. Animal Model and Treatment
4.7. Immunofluorescence Staining of Caspase-3
4.8. TUNEL Assay
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, X.Y.; Liu, G.C.; Zhang, J.X.; Wang, L.H.; Xu, C.; Yan, Z.A.; Wang, A.; Su, Y.F.; Lee, J.J.; Piao, G.C.; et al. Pharmacological Properties of Ginsenoside Re. Front. Pharmacol. 2022, 13, 754191. [Google Scholar] [CrossRef]
- Song, X.; Wang, L.; Fan, D. Insights into Recent Studies on Biotransformation and Pharmacological Activities of Ginsenoside Rd. Biomolecules 2022, 12, 512. [Google Scholar] [CrossRef]
- Kim, S.; Kim, N.; Jeong, J.; Lee, S.; Kim, W.; Ko, S.-G.; Kim, B. Anti-Cancer Effect of Panax Ginseng and Its Metabolites: From Traditional Medicine to Modern Drug Discovery. Processes 2021, 9, 1344. [Google Scholar] [CrossRef]
- Xu, J.; Pan, Y.; Liu, Y.; Na, S.; Zhou, H.; Li, L.; Chen, F.; Song, H. A review of anti-tumour effects of ginsenoside in gastrointestinal cancer. J. Pharm. Pharmacol. 2021, 73, 1292–1301. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, Z.; Zhang, S.; Li, X.; To, S.K.Y.; Peng, Y.; Liu, J.; Chen, S.; Hu, H.; Wong, A.S.T.; et al. The Ginsenoside Compound K Suppresses Stem-Cell-like Properties and Colorectal Cancer Metastasis by Targeting Hypoxia-Driven Nur77-Akt Feed-Forward Signaling. Cancers 2022, 15, 24. [Google Scholar] [CrossRef]
- Zhang, H.; Yi, J.K.; Huang, H.; Park, S.; Kwon, W.; Kim, E.; Jang, S.; Kim, S.Y.; Choi, S.K.; Yoon, D.; et al. 20 (S)-ginsenoside Rh2 inhibits colorectal cancer cell growth by suppressing the Axl signaling pathway in vitro and in vivo. J. Ginseng Res. 2022, 46, 396–407. [Google Scholar] [CrossRef]
- Quan, L.H.; Kim, Y.J.; Li, G.H.; Choi, K.T.; Yang, D.C. Microbial transformation of ginsenoside Rb1 to compound K by Lactobacillus paralimentarius. World J. Microbiol. Biotechnol. 2013, 29, 1001–1007. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, Y.; Kim, M.G. Changes in the ginsenoside content during the fermentation process using microbial strains. J. Ginseng Res. 2015, 39, 392–397. [Google Scholar] [CrossRef]
- Bae, E.A.; Han, M.J.; Choo, M.K.; Park, S.Y.; Kim, D.H. Metabolism of 20(S)- and 20(R)-ginsenoside Rg3 by human intestinal bacteria and its relation to in vitro biological activities. Biol. Pharm Bull. 2002, 25, 58–63. [Google Scholar] [CrossRef]
- Zheng, M.M.; Xu, F.X.; Li, Y.J.; Xi, X.Z.; Cui, X.W.; Han, C.C.; Zhang, X.L. Study on Transformation of Ginsenosides in Different Methods. BioMed Res. Int. 2017, 2017, 8601027. [Google Scholar] [CrossRef]
- Jin, X.; Zhu, L.Y.; Shen, H.; Xu, J.; Li, S.L.; Jia, X.B.; Cai, H.; Cai, B.C.; Yan, R. Influence of sulphur-fumigation on the quality of white ginseng: A quantitative evaluation of major ginsenosides by high performance liquid chromatography. Food Chem. 2012, 135, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.H.; Park, S.H.; Jee, H.S.; Kim, K.T.; Paik, H.D.; Lee, J.Y. Enhancement of the cytotoxicities and antioxidative activities of white ginseng extract by hydrolysis under mild acidic conditions. Food Sci. Biotechnol. 2014, 23, 173–178. [Google Scholar] [CrossRef]
- Chu, L.L.; Huy, N.Q.; Tung, N.H. Microorganisms for Ginsenosides Biosynthesis: Recent Progress, Challenges, and Perspectives. Molecules 2023, 28, 1437. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, Y.-H.; Park, J.-M.; Bai, D.-H.; Jang, J.K.; Park, Y.-S. Bioconversion of ginsenosides from red ginseng extract using candida allociferrii JNO301 isolated from Meju. Mycobiology 2014, 42, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, Z.; Liu, J.; Qi, H.; Li, J.; Chen, J.; Huang, Q.; Liu, Q.; Mi, J.; Li, X. Gut Microbiota: Therapeutic Targets of Ginseng Against Multiple Disorders and Ginsenoside Transformation. Front. Cell. Infect. Microbiol. 2022, 12, 853981. [Google Scholar] [CrossRef] [PubMed]
- Kumar Bajaj, B.; Claes, I.J.; Lebeer, S. Functional mechanisms of probiotics. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 321–327. [Google Scholar] [CrossRef]
- Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. Role of Probiotics and Diet in the Management of Neurological Diseases and Mood States: A Review. Microorganisms 2022, 10, 2268. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Na, C.S.; Yoo, S.A.; Seo, S.H.; Son, H.S. Biotransformation of major ginsenosides in ginsenoside model culture by lactic acid bacteria. J. Ginseng Res. 2017, 41, 36–42. [Google Scholar] [CrossRef]
- Renchinkhand, G.; Park, Y.W.; Cho, S.H.; Song, G.Y.; Bae, H.C.; Choi, S.J.; Nam, M.S. Identification of β-Glucosidase Activity of Lactobacillus plantarum CRNB22 in Kimchi and Its Potential to Convert Ginsenoside Rb1 from Panax Ginseng. J. Food Biochem. 2015, 39, 155–163. [Google Scholar] [CrossRef]
- Eom, S.J.; Kim, K.T.; Paik, H.D. Microbial bioconversion of ginsenosides in Panax ginseng and their improved bioactivities. Food Rev. Int. 2018, 34, 698–712. [Google Scholar] [CrossRef]
- Kim, B.G. Changes of ginsenosides in Korean red ginseng (Panax ginseng) fermented by Lactobacillus plantarum M1. Process Biochem. 2010, 45, 1319–1324. [Google Scholar] [CrossRef]
- Wu, L.; Jin, Y.; Yin, C.; Bai, L. Co-transformation of Panax major ginsenosides Rb₁ and Rg₁ to minor ginsenosides C-K and F₁ by Cladosporium cladosporioides. J. Ind. Microbiol. Biotechnol. 2012, 39, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhang, R.; Huang, Z.; Zhou, J. Progress in the Conversion of Ginsenoside Rb1 into Minor Ginsenosides Using β-Glucosidases. Foods 2023, 12, 397. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, S.J.; Liang, Y.L.; Sun, Y.; Cao, H.J.; Han, Y. Identification of the protopanaxatriol synthase gene CYP6H for ginsenoside biosynthesis in Panax quinquefolius. Funct. Integr. Genom. 2014, 14, 559–570. [Google Scholar] [CrossRef]
- Li, G.H.; Shen, Y.M.; Liu, Y.; Zhang, K.Q. Production of saponin in fermentation process of Sanchi (Panax notoginseng) and biotransformation of saponin by Bacillus subtilis. Ann. Microbiol. 2006, 56, 151–153. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yamabe, N.; Choi, P.; Lee, J.W.; Ham, J.; Kang, K.S. Efficient thermal deglycosylation of ginsenoside Rd and its contribution to the improved anticancer activity of ginseng. J. Agric. Food Chem. 2013, 61, 9185–9191. [Google Scholar] [CrossRef]
- Quan, L.H.; Min, J.W.; Jin, Y.; Wang, C.; Kim, Y.J.; Yang, D.C. Enzymatic biotransformation of ginsenoside Rb1 to compound K by recombinant β-glucosidase from Microbacterium esteraromaticum. J. Agric. Food Chem. 2012, 60, 3776–3781. [Google Scholar] [CrossRef]
- Gong, L.; Yin, J.; Zhang, Y.; Huang, R.; Lou, Y.; Jiang, H.; Sun, L.; Jia, J.; Zeng, X. Neuroprotective Mechanisms of Ginsenoside Rb1 in Central Nervous System Diseases. Front. Pharmacol. 2022, 13, 914352. [Google Scholar] [CrossRef]
- Nakhjavani, M.; Smith, E.; Townsend, A.R.; Price, T.J.; Hardingham, J.E. Anti-Angiogenic Properties of Ginsenoside Rg3. Molecules 2020, 25, 4905. [Google Scholar] [CrossRef]
- Tang, W.; Zhang, Y.; Gao, J.; Ding, X.; Gao, S. The anti-fatigue effect of 20(R)-ginsenoside Rg3 in mice by intranasally administration. Biol. Pharm. Bull. 2008, 31, 2024–2027. [Google Scholar] [CrossRef]
- Li, H.; Huang, N.; Zhu, W.; Wu, J.; Yang, X.; Teng, W.; Tian, J.; Fang, Z.; Luo, Y.; Chen, M.; et al. Modulation the crosstalk between tumor-associated macrophages and non-small cell lung cancer to inhibit tumor migration and invasion by ginsenoside Rh2. BMC Cancer 2018, 18, 579. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Mou, Y.C.; Su, C.C.; Chiang, B.H. Antihepatocarcinoma activity of lactic acid bacteria fermented Panax notoginseng. J. Agric. Food Chem. 2010, 58, 8528–8534. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Liu, D.; Xia, J.; Xu, J.F.; Zhang, L.; Yang, Y.; Wu, J.J.; Ao, H. Ginsenoside CK, rather than Rb1, possesses potential chemopreventive activities in human gastric cancer via regulating PI3K/AKT/NF-κB signal pathway. Front. Pharmacol. 2022, 13, 977539. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.K.; Dong, H.; Liang, X.; Bai, L.P.; Jiang, Z.H.; Guo, Y.; Kong, A.N.; Wang, R.; Kam, R.K.; Law, B.Y.; et al. Rh2E2, a novel metabolic suppressor, specifically inhibits energy-based metabolism of tumor cells. Oncotarget 2016, 7, 9907–9924. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Jiang, B.; Liu, W.; Wang, P.; Mou, Y.; Liu, Y.; Zhao, Y.; Bi, X. Anti-tumor activity of three novel derivatives of ginsenoside on colorectal cancer cells. Steroids 2014, 80, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Dong, Y.; Wang, L.; Xu, G.; Yang, Q.; Tang, X.; Qiao, Y.; Cong, Z. Ginsenoside metabolite compound K induces apoptosis and autophagy in non-small cell lung cancer cells via AMPK-mTOR and JNK pathways. Biochem. Cell Biol. 2019, 97, 406–414. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, P.; Duan, Z.; Liu, Y.; Mi, Y.; Fan, D. Ginsenoside Rh4 Suppressed Metastasis of Lung Adenocarcinoma via Inhibiting JAK2/STAT3 Signaling. Int. J. Mol. Sci. 2022, 23, 2018. [Google Scholar] [CrossRef]
- Han, S.; Jeong, A.J.; Yang, H.; Bin Kang, K.; Lee, H.; Yi, E.H.; Kim, B.H.; Cho, C.H.; Chung, J.W.; Sung, S.H.; et al. Ginsenoside 20(S)-Rh2 exerts anti-cancer activity through targeting IL-6-induced JAK2/STAT3 pathway in human colorectal cancer cells. J. Ethnopharmacol. 2016, 194, 83–90. [Google Scholar] [CrossRef]
- Yue, W.; Zheng, X.; Lin, Y.; Yang, C.S.; Xu, Q.; Carpizo, D.; Huang, H.; DiPaola, R.S.; Tan, X.L. Metformin combined with aspirin significantly inhibit pancreatic cancer cell growth in vitro and in vivo by suppressing anti-apoptotic proteins Mcl-1 and Bcl-2. Oncotarget 2015, 6, 21208–21224. [Google Scholar] [CrossRef]
- Ooi, K.L.; Tengku Muhammad, T.S.; Lam, L.Y.; Sulaiman, S.F. Cytotoxic and Apoptotic Effects of Ethyl Acetate Extract of Elephantopus mollis Kunth. in Human Liver Carcinoma HepG2 Cells Through Caspase-3 Activation. Integr. Cancer Ther. 2014, 13, NP1–NP9. [Google Scholar] [CrossRef]
- Hong, S.; Cai, W.; Huang, Z.; Wang, Y.; Mi, X.; Huang, Y.; Lin, Z.; Chen, X. Ginsenoside Rg3 enhances the anticancer effect of 5-FU in colon cancer cells via the PI3K/AKT pathway. Oncol. Rep. 2020, 44, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Yang, J.H.; Yang, H.J.; Cho, C.K.; Choi, J.G.; Chung, H.S. Antitumor Effect of Korean Red Ginseng through Blockade of PD-1/PD-L1 Interaction in a Humanized PD-L1 Knock-In MC38 Cancer Mouse Model. Int. J. Mol. Sci. 2023, 24, 1894. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Zhao, C.; Yang, C.; Zhou, Q.; Liu, X.; Yang, P.; Yang, F.; Shi, X. Limosilactobacillus fermentum-fermented ginseng improved antibiotic-induced diarrhoea and the gut microbiota profiles of rats. J. Appl. Microbiol. 2022, 133, 3476–3489. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Liao, H.F.; Wang, T.E.; Lin, S.C.; Shih, S.C.; Chang, W.H.; Yang, Y.C.; Lin, C.C.; Chen, Y.J. Etoposide sensitizes CT26 colorectal adenocarcinoma to radiation therapy in BALB/c mice. World J. Gastroenterol. 2005, 11, 4895–4898. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Retention Time (min) | Ginsenoside | TGs Content (mg/g) | FTGs Content (mg/g) |
|---|---|---|---|---|
| 1 | 35.81 | Rg1 | 77.87 ± 0.23 | 9.56 ± 0.36 |
| 2 | 38.46 | Re | 135.74 ± 0.31 | 15.10 ± 0.42 |
| 3 | 55.06 | Rb1 | 238.15 ± 0.45 | 161.50 ± 0.74 |
| 4 | 56.71 | Rb2 | 103.43 ± 0.25 | 74.21 ± 0.52 |
| 5 | 59.56 | Rc | 35.86 ± 0.54 | 27.32 ± 0.49 |
| 6 | 66.24 | Rd | 80.63 ± 0.68 | 66.72 ± 0.47 |
| 7 | 76.34 | Rg6 | 0 | 2.56 ± 0.16 |
| 8 | 77.84 | F4 | 0.52 ± 0.12 | 5.51 ± 0.23 |
| 9 | 78.86 | Rk3 | 0.50 ± 0.11 | 17.89 ± 0.53 |
| 10 | 80.08 | Rh4 | 0 | 9.08 ± 0.14 |
| 11 | 82.47 | 20(R)-Rg3 | 0 | 61.82 ± 0.26 |
| 12 | 83.42 | 20(S)-Rg3 | 0 | 37.96 ± 0.48 |
| 13 | 90.08 | CK | 0 | 14.73 ± 0.38 |
| 14 | 92.90 | Rh2 | 1.1 ± 0.05 | 62.37 ± 0.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Gao, Y.; Yang, G.; Zhao, Z.; Zhao, Y.; Gao, L.; Zhao, L.; Li, S. Transformation of Ginsenosides by Lactiplantibacillus plantarum MB11 Fermentation: Minor Ginsenosides Conversion and Enhancement of Anti-Colorectal Cancer Activity. Molecules 2024, 29, 27. https://doi.org/10.3390/molecules29010027
Shen Y, Gao Y, Yang G, Zhao Z, Zhao Y, Gao L, Zhao L, Li S. Transformation of Ginsenosides by Lactiplantibacillus plantarum MB11 Fermentation: Minor Ginsenosides Conversion and Enhancement of Anti-Colorectal Cancer Activity. Molecules. 2024; 29(1):27. https://doi.org/10.3390/molecules29010027
Chicago/Turabian StyleShen, Yunjiao, Yansong Gao, Ge Yang, Zijian Zhao, Yujuan Zhao, Lei Gao, Lei Zhao, and Shengyu Li. 2024. "Transformation of Ginsenosides by Lactiplantibacillus plantarum MB11 Fermentation: Minor Ginsenosides Conversion and Enhancement of Anti-Colorectal Cancer Activity" Molecules 29, no. 1: 27. https://doi.org/10.3390/molecules29010027