
Rapid Screening of Phenolic Compounds with Anti-Enteritis Activity from Camellia oleifera Oil Using a Smurf Drosophila Model and Molecular Docking Methods

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Anti-Enteritis Activity of PECS
2.2. Screening of Phenolic Compounds in PECS with Anti-Enteritis Activity
2.2.1. Metabolomics Analysis Based on UPLC-QTRAP/MS
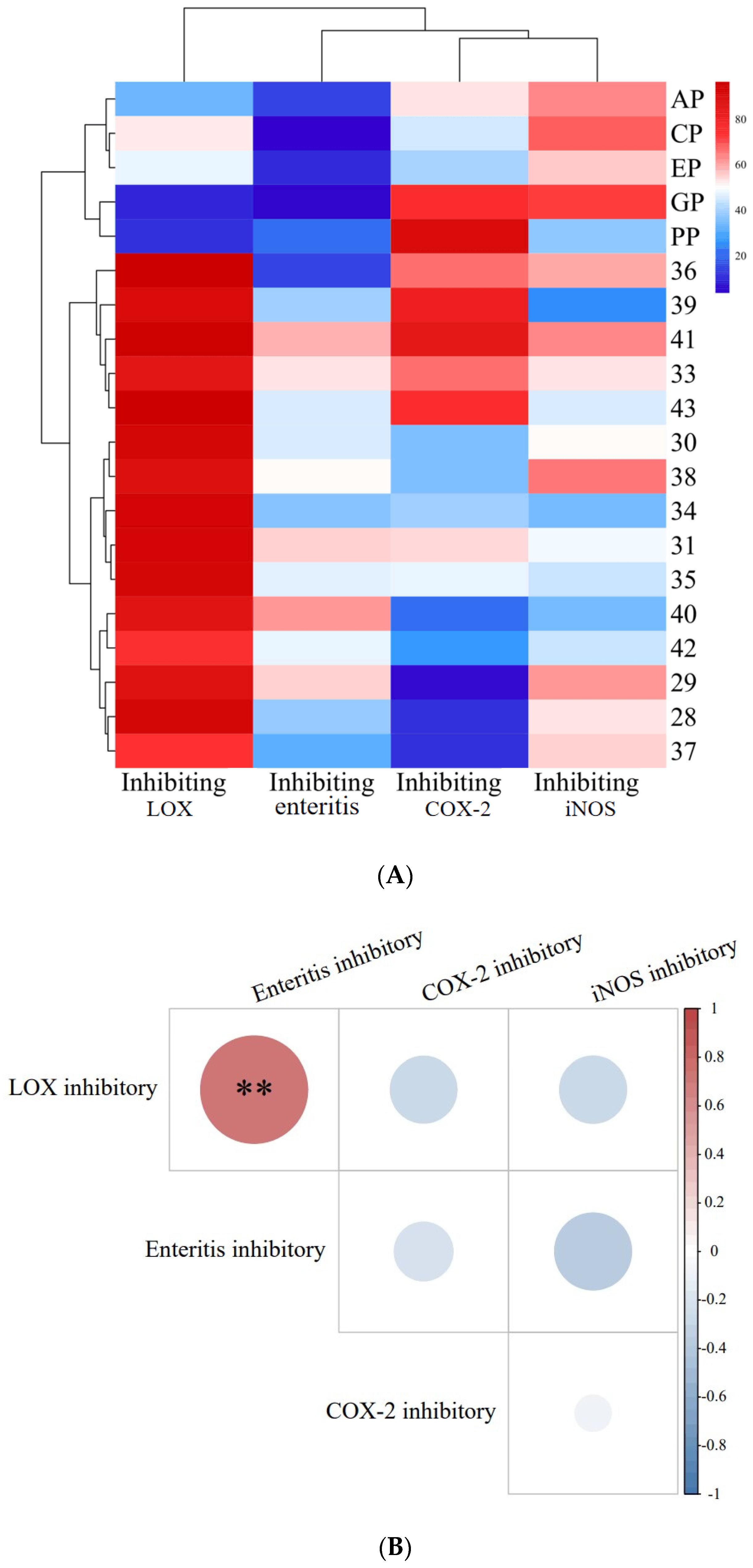
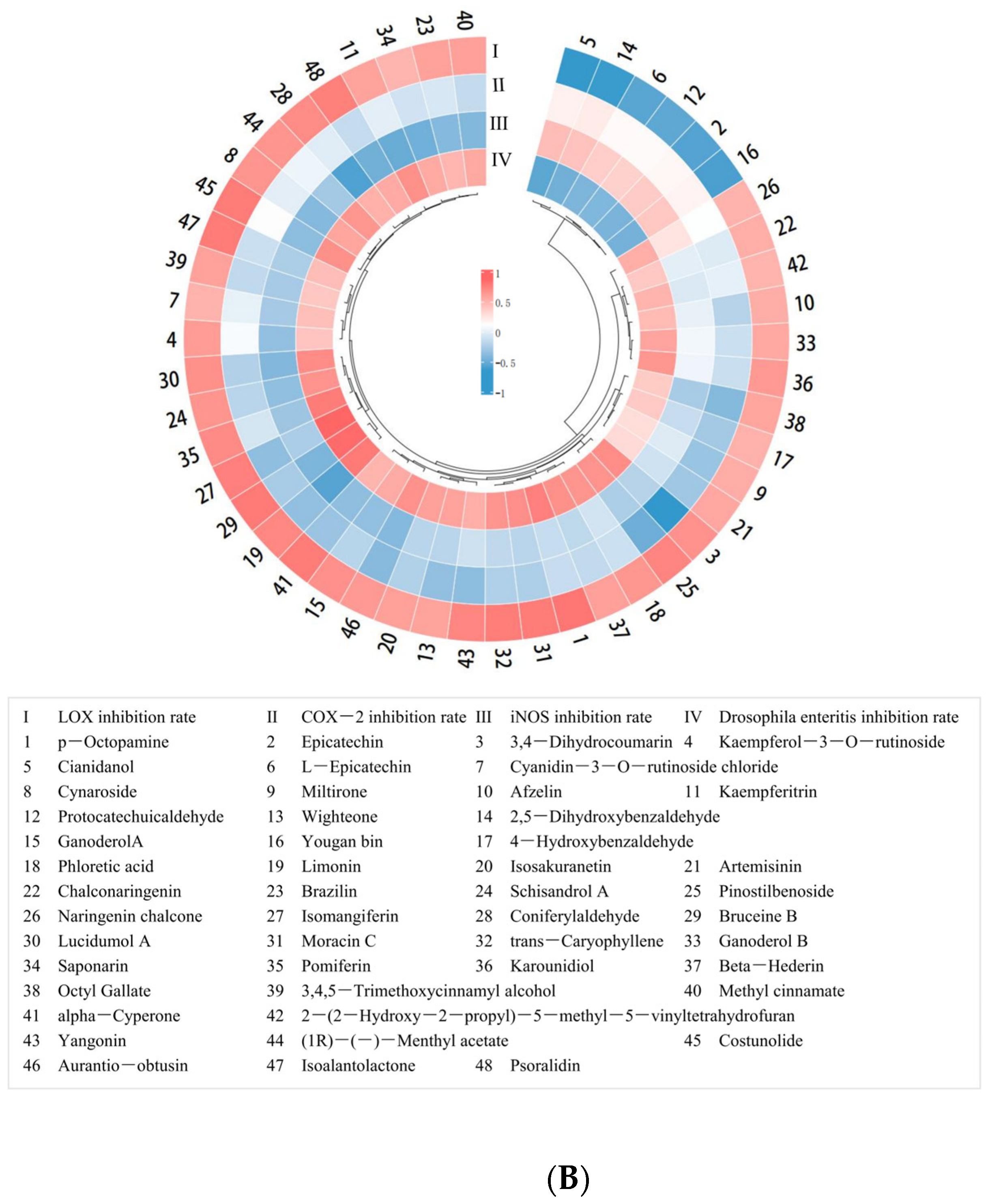
2.2.2. Correlation Analysis between Small-Molecule Organic Compounds’ Content and Anti-Enteritis Activity of PECS
2.2.3. Molecular Docking Analysis
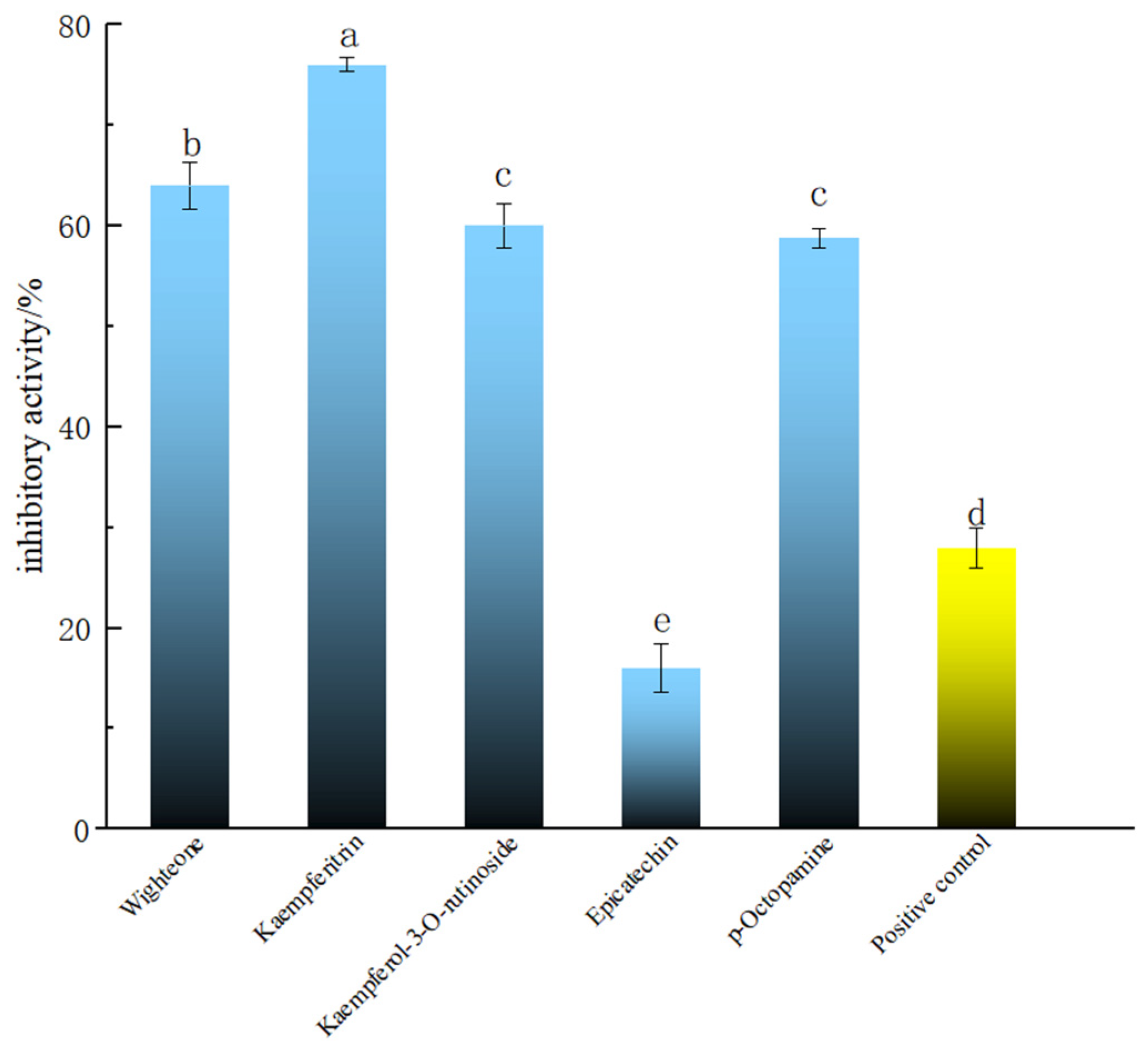
2.3. Screening of Polyphenolic Compounds in PECS with Anti-Enteritis Activity Using the Smurf Drosophila Assay
2.4. Inhibition of COX-2, LOX, and iNOS Activity by Phenolic Compounds
3. Materials and Methods
3.1. Materials
3.2. Preparation of Polyphenol Extracts from C. oleifera Oil
3.3. Determination of Anti-Inflammatory Activity In Vivo
3.4. Determination of the Inhibitory of 5-Lipoxygenase (LOX), COX-2, and iNOS
3.5. Metabolomics Analysis Based on UPLC-QTRAP/MS
3.6. Molecular Docking Analysis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lei, X.; Liu, Q.; Liu, Q.; Cao, Z.; Zhang, J.; Kuang, T.; Fang, Y.; Liu, G.; Qian, K.; Fu, J. Camellia oil (Camellia oleifera Abel.) attenuates CCl4 -induced liver fibrosis via suppressing hepatocyte apoptosis in mice. Food Funct. 2020, 11, 4582–4590. [Google Scholar] [CrossRef]
- Xiao, X.; He, L.; Chen, Y.; Wu, L.; Wang, L.; Liu, Z. Anti-inflammatory and antioxidative effects of Camellia oleifera Abel components. Future Med. Chem. 2017, 9, 2069–2079. [Google Scholar] [CrossRef]
- Tung, Y.T.; Tu, P.S.; Lee, W.T.; Yen, G.C. Protective effect of Camellia Oil (Camellia oleifera Abel.) against ethanol-induced acute oxidative injury of the gastric mucosa in mice. J. Agric. Food Chem. 2017, 65, 4932–4941. [Google Scholar] [CrossRef]
- Chang, M.; Qiu, F.; Lan, N.; Zhang, T.; Guo, X.; Jin, Q.; Liu, R.; Wang, X. Analysis of phytochemical composition of Camellia oleifera oil and evaluation of its anti-inflammatory effect in lipopolysaccharide- stimulated RAW 2647 macrophages. Lipids 2020, 55, 353–363. [Google Scholar] [CrossRef]
- Zhang, T.; Qiu, F.; Chen, L.; Liu, R.; Wang, X. Identification and in vitro anti-inflammatory activity of different forms of phenolic compounds in Camellia oleifera oil. Food Chem. 2020, 344, 128660. [Google Scholar] [CrossRef]
- Salmaso, V.; Moro, S. Bridging molecular docking to molecular dynamics in exploring ligand-protein recognition process: An overview. Front. Pharmacol. 2018, 9, 923. [Google Scholar] [CrossRef]
- Rajesh, S.V.; Gnanendra, T.S.; Kumar, T.S. Anti- inflammatory and molecular docking analysis of leucas vestita wall. Ex. Benth. extract against cyclo-oxygenase-2. Prostaglandins Other Lipid Mediat. 2021, 156, 106581. [Google Scholar] [CrossRef]
- Che, Y.H.; Xu, Z.R.; Ni, L.L.; Dong, X.X.; Yang, Z.Z.; Yang, Z.B. Isolation and identification of the components in Cybister chinensis Motschulsky against inflammation and their mechanisms of action based on network pharmacology and molecular docking. J. Ethnopharmacol. 2022, 285, 114851. [Google Scholar] [CrossRef]
- Apidianakis, Y.; Rahme, L.G. Drosophila melanogaster as a model for human intestinal infection and pathology. Dis. Models Mech. 2011, 4, 21–30. [Google Scholar] [CrossRef]
- Lee, K.A.; Lee, W.J. Drosophila as a model for intestinal dysbiosis and chronic inflammatory diseases. Dev. Comp. Immunol. 2014, 42, 102–110. [Google Scholar] [CrossRef]
- Keshav, N.; Ammankallu, R.; Shashidhar; Paithankar, J.G.; Baliga, M.S.; Patil, R.K.; Kudva, A.K.; Raghu, S.V. Dextran sodium sulfate alters antioxidant status in the gut affecting the survival of Drosophila melanogaster. 3 Biotech 2022, 12, 280. [Google Scholar] [CrossRef]
- Toda, K.; Tsukayama, I.; Nagasaki, Y.; Konoike, Y.; Tamenobu, A.; Ganeko, N.; Ito, H.; Kawakami, Y.; Takahashi, Y.; Miki, Y. Red-kerneled rice proanthocyanidin inhibits arachidonate 5-lipoxygenase and decreases psoriasis-like skin inflammation. Arch. Biochem. Biophys. 2020, 689, 108307. [Google Scholar] [CrossRef]
- Richard, W.; Piper, M.D.W.; Bregje, W.; Linda, P.; Raine, N.E. Quantification of food intake in Drosophila. PLoS ONE 2009, 4, e6063. [Google Scholar] [CrossRef]
- Rera, M.; Bahadorani, S.; Cho, J.; Koehler, C.L.; Ulgherait, M.; Hur, J.H.; Ansari, W.S.; Lo, T.; Jones, D.L.; Walker, D.W. Modulation of longevity and tissue homeostasis by the Drosophila PGC-1 homolog. Cell Metab. 2011, 14, 623–634. [Google Scholar] [CrossRef]
- Tsuge, A.S.; Inazumi, T.; Shimamoto, A.; Sugimoto, Y. Molecular mechanisms underlying prostaglandin E-2-exacerbated inflammation and immune diseases. Int. Immunol. 2019, 31, 597–606. [Google Scholar] [CrossRef]
- Yao, C.; Narumiya, S. Prostaglandin-cytokine crosstalk in chronic inflammation. Br. J. Pharmacol. 2019, 176, 337–354. [Google Scholar] [CrossRef]
- Desai, S.J.; Prickril, B.; Rasooly, A. Mechanisms of phytonutrient modulation of cyclooxygenase-2 (COX-2) and inflammation related to cancer. Nutr. Cancer 2018, 70, 350–375. [Google Scholar] [CrossRef]
- Araújo, F.F.D.; Farias, D.D.P.; Neri-Numa, I.A.; Pastore, G.M. Polyphenols and their applications: An approach in food chemistry and innovation potential. Food Chem. 2020, 338, 127535. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, Q.; del Mar Contreras, M.; Wang, L. Profiling and quantification of phenolic compounds in Camellia seed oils: Natural tea polyphenols in vegetable oil. Food Res. Int. 2017, 102, 184–194. [Google Scholar] [CrossRef]
- Hong, C.; Chang, C.; Zhang, H.; Jin, Q.; Wu, G.; Wang, X. Identification and characterization of polyphenols in different varieties of Camellia oleifera seed cakes by UPLC-QTOF-MS. Food Res. Int. 2019, 126, 108614. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, M.; Kim, J.M.; Lee, M.; Seo, S.J.; Park, K.Y. Afzelin suppresses proinflammatory responses in particulate matter-exposed human keratinocytes. Int. J. Mol. Med. 2019, 43, 2516–2522. [Google Scholar] [CrossRef]
- Akter, M.; Parvin, M.S.; Hasan, M.M.; Rahman, M.A.A.; Islam, M.E. Anti-tumor and antioxidant activity of kaempferol-3-O-alpha-L-rhamnoside (Afzelin) isolated from Pithecellobium dulce leaves. BMC Complement. Med. Ther. 2022, 22, 1–11. [Google Scholar] [CrossRef]
- Šarić, A.; Sobočanec, S.; Balog, T.; Kušić, B.; Šverko, V.; Dragović-Uzelac, V.; Levaj, B.; Čosić, Z.; Marotti, Ž.M.Š. Improved antioxidant and anti-inflammatory potential in mice consuming sour cherry juice (Prunus Cerasus cv. Maraska). Plant Foods Hum. Nutr. 2009, 64, 231–237. [Google Scholar] [CrossRef]
- Akkarachiyasit, S.; Yibchok-Anun, S.; Wacharasindhu, S.; Adisakwattana, S. In vitro inhibitory effects of Cyandin-3-rutinoside on pancreatic α-amylase and its combined effect with acarbose. Molecules 2011, 16, 2075–2083. [Google Scholar] [CrossRef]
- Sun, P.; Qu, Y.; Wang, Y.; Wang, J.; Wang, X.; Sheng, J. Wighteone exhibits an antitumor effect against EGFR L858R/T790M mutation non-small cell lung cancer. J. Cancer 2021, 12, 3900–3908. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, Y.; Zhang, Y.; Zhang, H.; Xu, L.; Zou, Z.; Yang, W.; Cheng, J.; Zhou, Y. Genome-wide analysis of the expression profile of Saccharomyces cerevisiae in response to treatment with the plant isoflavone, wighteone, as a potential antifungal agent. Biotechnol. Lett. 2006, 28, 99–105. [Google Scholar] [CrossRef]
- Uckoo, R.M.; Jayaprakasha, G.K.; Nelson, S.D.; Patil, B.S. Rapid simultaneous determination of amines and organic acids in citrus using high-performance liquid chromatography. Talanta 2011, 83, 948–954. [Google Scholar] [CrossRef]
- Stohs, S.J. Physiological functions and pharmacological and toxicological effects of p-octopamine. Drug Chem. Toxicol. 2015, 38, 106–112. [Google Scholar] [CrossRef]
- Guo, X.H.; Ye, M.Z.; Wu, L.H. Molecular docking study on the anti-osteoarthritis mechanism of fisetin. J. Gannan Med. Univ. 2021, 41, 832–837. [Google Scholar] [CrossRef]
- Aier, I.; Varadwaj, P.K.; Raj, U. Structural insights into conformational stability of both wild-type and mutant EZH2 receptor. Sci. Rep. 2016, 6, 34984. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Q. Kaempferitrin inhibits proliferation, induces apoptosis, and ameliorates inflammation in human rheumatoid arthritis fibroblast-like synoviocytes. Phytother. Res. 2019, 33, 1726–1735. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, X.; Liu, L.; Shen, J. Polyphenol-rich extracts from Oiltea camellia prevent weight gain in obese mice fed a high-fat diet and slowed the accumulation of triacylglycerols in 3T3-L1 adipocytes. J. Funct. Foods 2014, 9, 148–155. [Google Scholar] [CrossRef]
- Yu, J.J.; Yang, S.; Pan, L. Smurf assay to detect Drosophila intestine leakage. Bio-101 2019, e1010267. [Google Scholar] [CrossRef]
- Huang, Y.; Hua, Y.; Qiu, A. Soybean protein aggregation induced by lipoxygenase catalyzed linoleic acid oxidation. Food Res. Int. 2006, 39, 240–249. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds | Docking Binding Energy (kcal mol−1) | RMSD | ||||
|---|---|---|---|---|---|---|
| LOX | COX-2 | iNOS | LOX | COX-2 | iNOS | |
| p-Octopamine | −5.1 | −5.31 | −3.3 | 1.760 | 1.770 | 1.731 |
| 3-(2-hydroxyphenyl)propanoic acid | −4.82 | −4.05 | −4.75 | 2.522 | 2.638 | 2.677 |
| cyanidin 3-rutinoside | −5.77 | −5.20 | −3.26 | 2.772 | 1.974 | 1.987 |
| (1R)-(−)-Menthyl acetate | −5.25 | −5.04 | −4.8 | 0.000 | 0.000 | 0.000 |
| Ganoderol B | −9.47 | −11.16 | −9.59 | 0.001 | 0.527 | 0.001 |
| Karounidiol | −12.23 | −12.11 | −10.7 | 0.000 | 0.000 | 0.000 |
| Wighteone | −5.56 | −5.56 | −5.03 | 0.000 | 0.229 | 0.001 |
| Lucidumol A | −10.06 | −12.83 | −10.83 | 0.000 | 0.000 | 0.294 |
| β-Hederin | −8.68 | −13.62 | −11.41 | 0.624 | 0.767 | 0.521 |
| Limonin | −6.40 | −7.04 | −6.33 | 0.000 | 0.000 | 0.000 |
| Isomangiferin | −2.96 | −4.41 | −4.92 | 0.001 | 0.008 | 0.001 |
| Chalconaringenin | −2.17 | −0.84 | −2.09 | 1.918 | 0.431 | 1.004 |
| Bruceine B | −6.25 | −5.87 | −5.62 | 0.001 | 0.001 | 0.000 |
| Afzelin | −3.62 | −3.54 | −3.15 | 0.484 | 0.522 | 0.601 |
| Cynaroside | −3.60 | −6.00 | −3.04 | 1.335 | 0.784 | 1.318 |
| Kaempferitrin | −9.27 | −13.05 | −9.51 | 0.323 | 0.352 | 0.588 |
| Kaempferol-3-O-rutinoside | −10.49 | −14.49 | −10.62 | 0.596 | 0.650 | 0.460 |
| Cyanidin 3-rutinoside | −5.77 | −5.20 | −3.26 | 2.772 | 1.974 | 1.987 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Li, Y.; Lin, X.; Fu, X.; Zhong, H.; Ren, K.; Liu, C.; Yao, W. Rapid Screening of Phenolic Compounds with Anti-Enteritis Activity from Camellia oleifera Oil Using a Smurf Drosophila Model and Molecular Docking Methods. Molecules 2024, 29, 76. https://doi.org/10.3390/molecules29010076
Wang S, Li Y, Lin X, Fu X, Zhong H, Ren K, Liu C, Yao W. Rapid Screening of Phenolic Compounds with Anti-Enteritis Activity from Camellia oleifera Oil Using a Smurf Drosophila Model and Molecular Docking Methods. Molecules. 2024; 29(1):76. https://doi.org/10.3390/molecules29010076
Chicago/Turabian StyleWang, Shuhao, Yang Li, Xin Lin, Xiangjin Fu, Haiyan Zhong, Kangzi Ren, Cheng Liu, and Wen Yao. 2024. "Rapid Screening of Phenolic Compounds with Anti-Enteritis Activity from Camellia oleifera Oil Using a Smurf Drosophila Model and Molecular Docking Methods" Molecules 29, no. 1: 76. https://doi.org/10.3390/molecules29010076
APA StyleWang, S., Li, Y., Lin, X., Fu, X., Zhong, H., Ren, K., Liu, C., & Yao, W. (2024). Rapid Screening of Phenolic Compounds with Anti-Enteritis Activity from Camellia oleifera Oil Using a Smurf Drosophila Model and Molecular Docking Methods. Molecules, 29(1), 76. https://doi.org/10.3390/molecules29010076