
Beneficial In Vitro Effects of Polysaccharide and Non-Polysaccharide Components of Dendrobium huoshanense on Gut Microbiota of Rats with Type 1 Diabetes as Opposed to Metformin

Abstract
:1. Introduction
2. Results
2.1. Main Content of DHWE, DHP, and NDHP
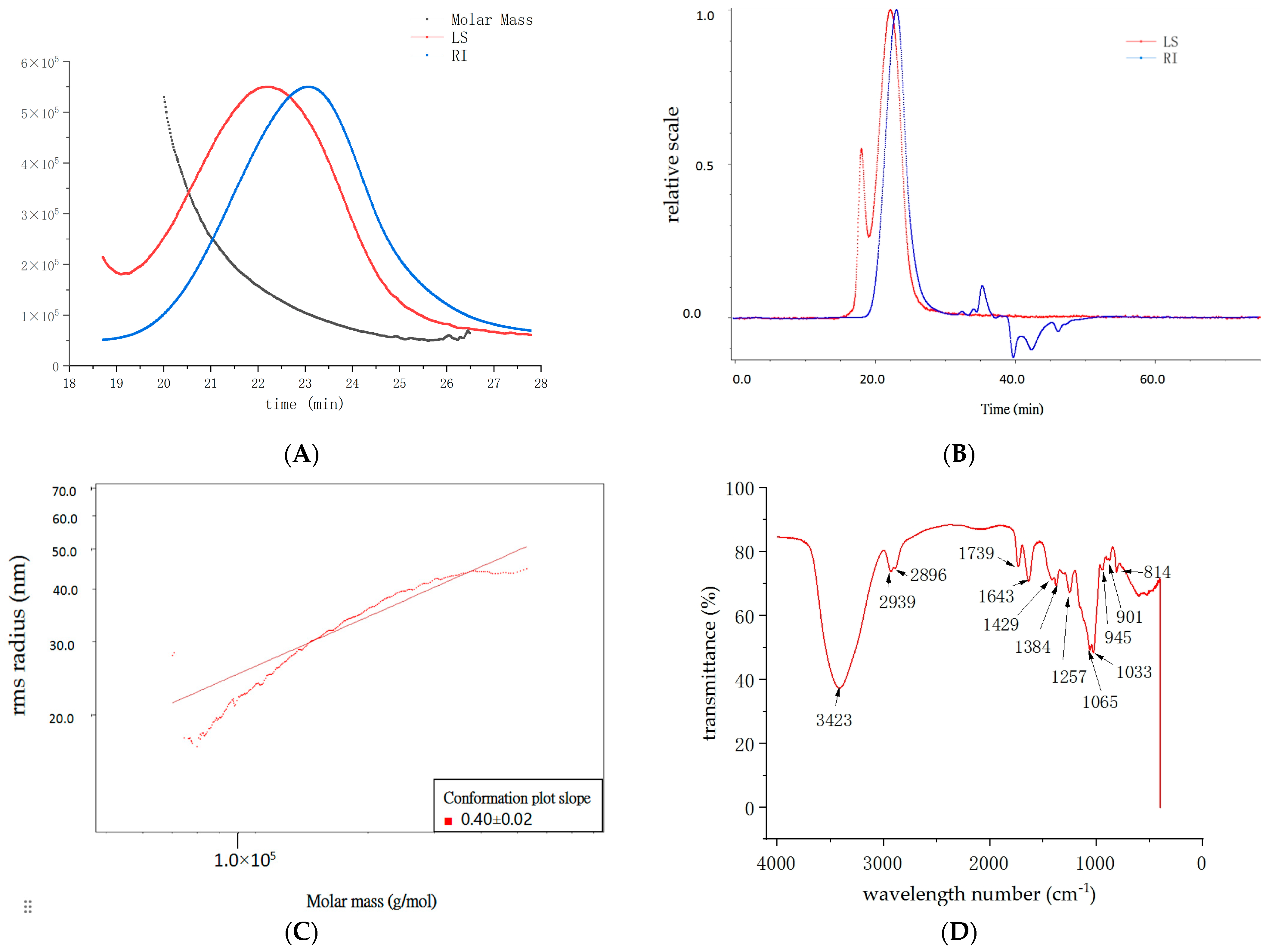
2.2. Analysis of the Structural Characteristics of DHP
2.3. Analysis of the Components of DHWE and NDHP with UPLC-Orbitrap-MS System
2.4. Effect of DHP, DHWE, NDHP, and Metformin (MET) on pH Change during Fermentation
2.5. Effect of DHP, DHWE, NDHP, and Metformin (MET) on Bacterial Growth during Fermentation
2.6. Effect of DHP, DHWE, NDHP, and Metformin (MET) on Short-Chain Fatty Acids (SCFAs) during Fermentation
2.7. Structure of the Fecal Microbiota
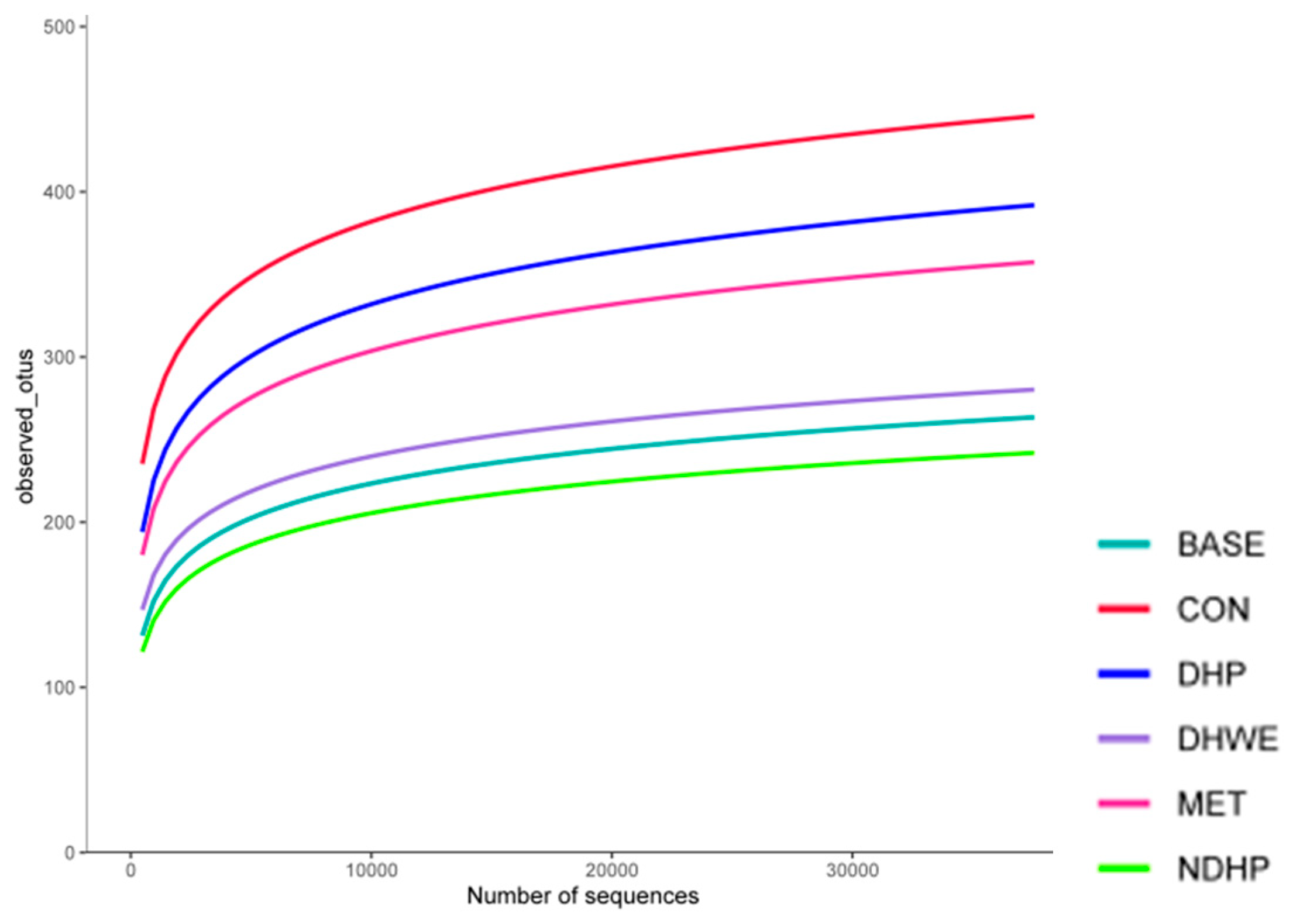
2.7.1. Alpha-Diversity Analysis
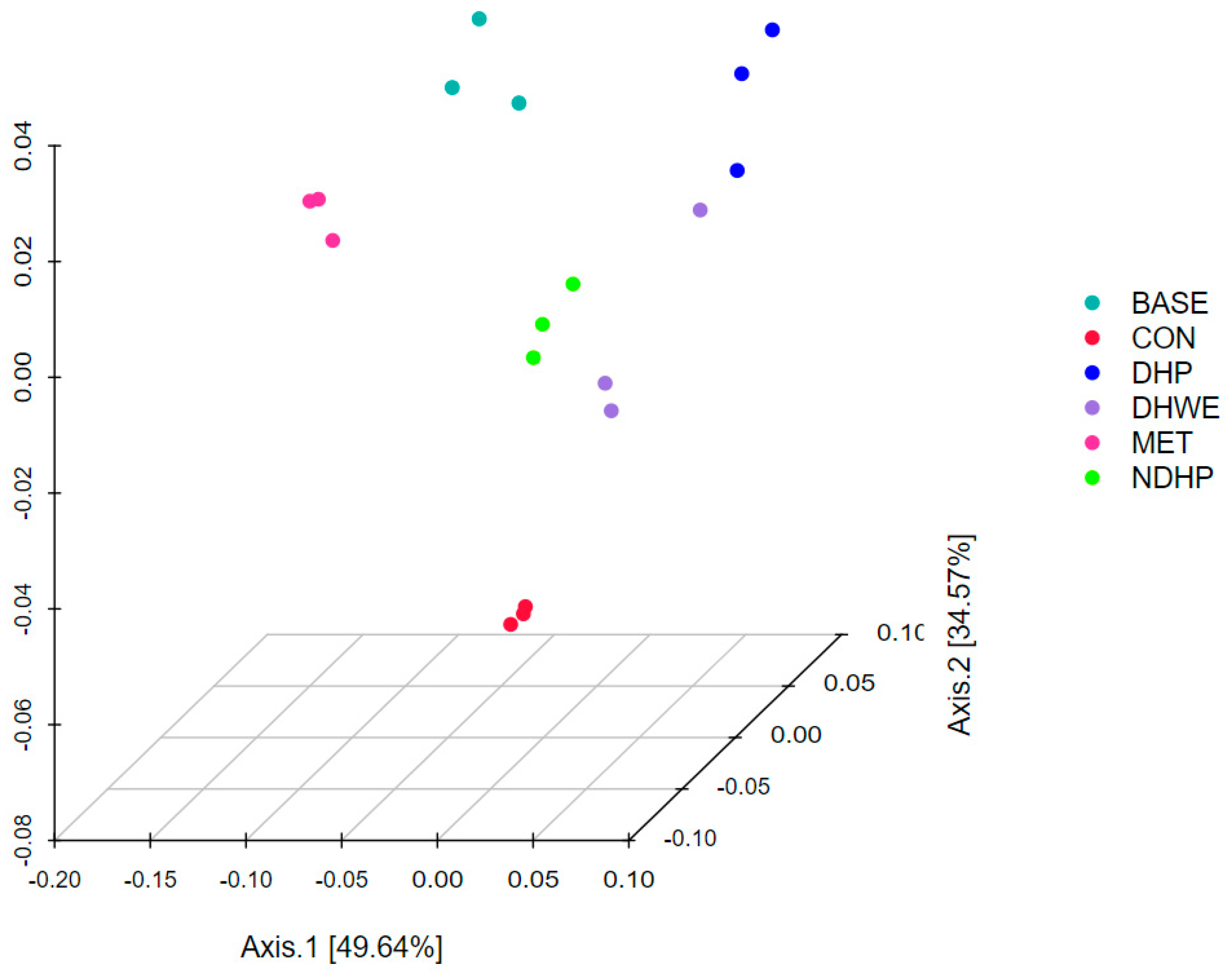
2.7.2. β-Diversity Analysis
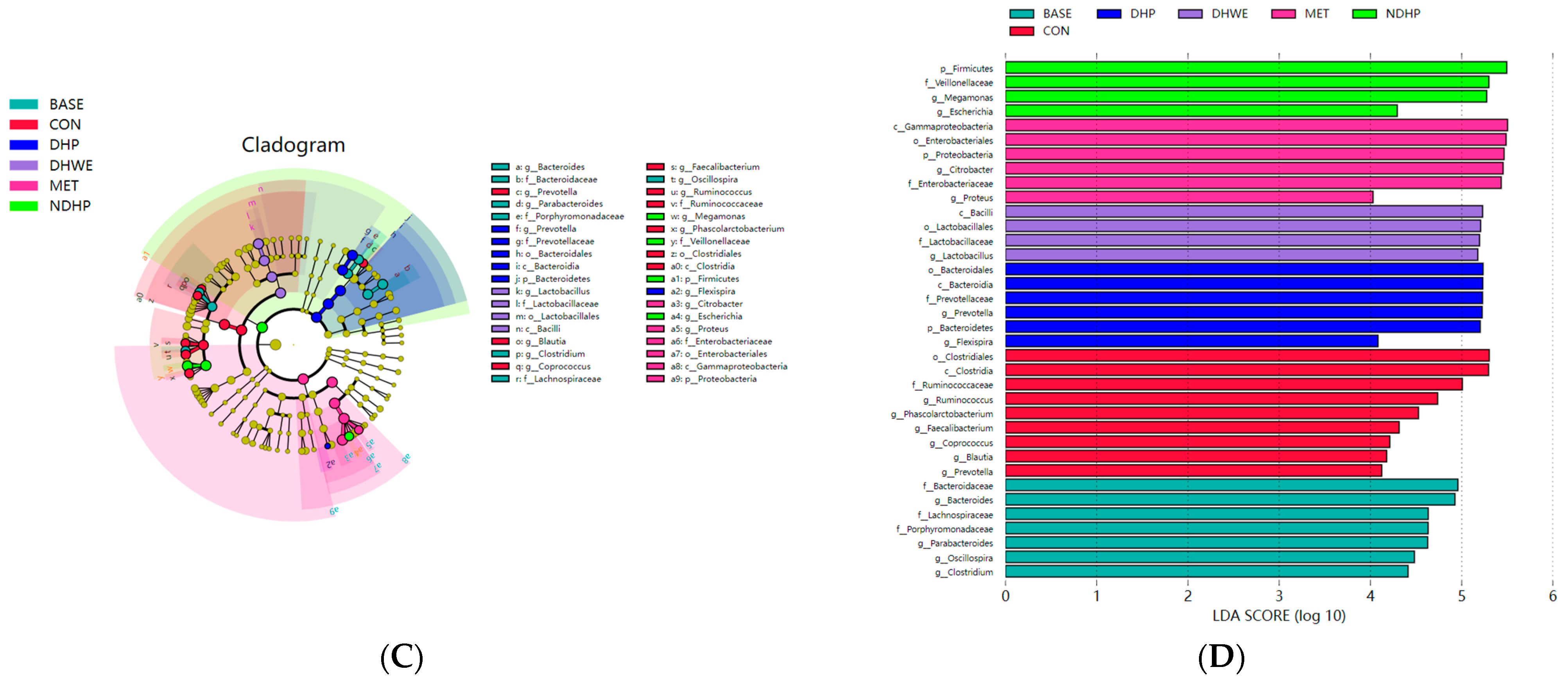
2.7.3. Key Phylotypes of the Fecal Microbiota
2.7.4. Correlation Analysis between the Gut Microbiota, pH of Fermentation, and ΔOD600 of Fermentation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Preparation of DHWE
4.2.2. Preparation of DHP
4.2.3. Preparation of NDHP
4.2.4. Determination of Carbohydrates, Total Polyphenols, and Protein Content in DHWE, DHP, and NDHP
4.2.5. Determination of the Homogeneity and Molecular Weight of DHP
4.2.6. Determination of the Structure of Characteristic Functional Groups of DHP
4.2.7. Analysis of the Components of DHWE and NDHP by UPLC-Orbitrap-MS (Ultra-Performance Liquid Chromatography–Orbitrap–Mass Spectrometry)
4.2.8. Preparation of Metformin Solution
4.2.9. Preparation of Rats with T1D
4.2.10. Preparation of Medium for In Vitro Fermentation
4.2.11. Preparation of a Fecal Slurry for Fermentation
4.2.12. Determination of the pH and Bacterial Count in the Fermentation Broth
4.2.13. Determination of Short-Chain Fatty Acid Content in Fermentation Broth
4.2.14. Bioinformatics Analysis of High-Throughput Sequencing Data from Fecal Microbiota
4.2.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toren, E.; Burnette, K.S.; Banerjee, R.R.; Hunter, C.S.; Tse, H.M. Partners in crime: Beta-cells and autoimmune responses complicit in type 1 diabetes pathogenesis. Front. Immunol. 2021, 12, 756548. [Google Scholar] [CrossRef] [PubMed]
- Rewers, M.; Ludvigsson, J. Environmental risk factors for type 1 diabetes. Lancet 2016, 387, 2340–2348. [Google Scholar] [CrossRef] [PubMed]
- Pociot, F.; Lernmark, Å. Genetic risk factors for type 1 diabetes. Lancet 2016, 387, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.; Wong, F.S.; Szot, G.L.; Bluestone, J.A.; et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef]
- Han, H.; Li, Y.; Fang, J.; Liu, G.; Yin, J.; Li, T.; Yin, Y. Gut microbiota and type 1 diabetes. Int. J. Mol. Sci. 2018, 19, 995. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Huang, J.; Zhou, Z. Host gene effects on gut microbiota in type 1 diabetes. Biochem. Soc. Trans. 2022, 50, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.R.C.; Francozo, M.C.S.; de Oliveira, G.G.; Ignacio, A.; Castoldi, A.; Zamboni, D.S.; Ramos, S.G.; Camara, N.O.; de Zoete, M.R.; Palm, N.W.; et al. Gut microbiota translocation to the pancreatic lymph nodes triggers NOD2 activation and contributes to t1d onset. J. Exp. Med. 2016, 213, 1223–1239. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Trapika, I.G.S.C.; Tang, S.Y.S.; Cho, J.L.; Qi, Y.; Li, C.G.; Li, Y.; Yao, M.; Yang, D.; Liu, B.; et al. Mechanisms and active compounds polysaccharides and bibenzyls of medicinal Dendrobiums for diabetes management. Front. Nutr. 2022, 8, 811870. [Google Scholar] [CrossRef]
- Gao, L.; Wang, F.; Hou, T.; Geng, C.; Xu, T.; Han, B.; Liu, D. Dendrobium huoshanense c.z.tang et s.j.cheng: A review of its traditional uses, phytochemistry, and pharmacology. Front. Pharmacol. 2022, 13, 920823. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, C. The linkage of gut microbiota and the property theory of traditional chinese medicine (TCM): Cold-natured and sweet-flavored TCMs as an example. J. Ethnopharmacol. 2023, 306, 116167. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.Z.; Liu, B.; Ye, H.Y.; Li, Q.M.; Pan, L.H.; Zha, X.Q.; Liu, J.; Duan, J.; Luo, J.P. Dendrobium huoshanense polysaccharide regionally regulates intestinal mucosal barrier function and intestinal microbiota in mice. Carbohydr. Polym. 2019, 206, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.W. Isolation and Purification of Polyphenols from Dendrobium huoshanense and the Effect on Probiotics and Phathogenic Bacteria. Master’s Thesis, Anhui Polytechnic University, Hefei, China, 2020. [Google Scholar]
- Vella, S.; Buetow, L.; Royle, P.; Livingstone, S.; Colhoun, H.M.; Petrie, J.R. The use of metformin in type 1 diabetes: A systematic review of efficacy. Diabetologia 2010, 53, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Foretz, M.; Guigas, B.; Viollet, B. Metformin: Update on mechanisms of action and repurposing potential. Nat. Rev. Endocrinol. 2023, 19, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Foretz, M.; Hébrard, S.; Leclerc, J.; Zarrinpashneh, E.; Soty, M.; Mithieux, G.; Sakamoto, K.; Andreelli, F.; Viollet, B. Metformin inhibits hepatic gluconeogenesis in mice independently of the LKB1/AMPK pathway via a decrease in hepatic energy state. J. Clin. Investig. 2010, 120, 2355–2369. [Google Scholar] [CrossRef]
- Li, S.S.; Qi, Y.L.; Chen, L.X.; Qu, D.; Li, Z.M.; Gao, K.; Chen, J.B.; Sun, Y.S. Effects of panax ginseng polysaccharides on the gut microbiota in mice with antibiotic-associated diarrhea. Int. J. Biol. Macromol. 2019, 124, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Cardona Mena, D.; Román López, P. New perspectives in health: Gut microbiota. Int. J. Environ. Res. Public Health 2022, 19, 5828. [Google Scholar] [CrossRef]
- Hajiagha, M.N.; Taghizadeh, S.; Asgharzadeh, M.; Dao, S.; Ganbarov, K.; Köse, Ş.; Kafil, H.S. Gut microbiota and human body interactions: Its impact on health: A review. Curr. Pharm. Biotechnol. 2022, 23, 4–14. [Google Scholar] [CrossRef]
- Meng, G.; Wei, M.; Zhao, S.G.; Zhang, K.; Yu, M.Y. The effects of polysaccharides from Dendrobium huoshanense on physiological characteristics of Lactobacillus acidophilus. Food Sci. Technol. 2017, 42, 184–187. [Google Scholar]
- Alves-Santos, A.M.; Sugizaki, C.; Lima, G.C.; Naves, M. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Xiao, L.; Li, X.P.; Yu, J.H.; Liu, C. Megamonas Funiformis and Its Application. CN Patent CN109504615A, 22 March 2019. [Google Scholar]
- Gorvitovskaia, A.; Holmes, S.P.; Huse, S.M. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Duncan, S.H.; Mcwilliam Leitch, E.C.; Child, M.W.; Flint, H.J. pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, M.; Ferro, A.; Gruden, G. Gastrointestinal microbiota and type 1 diabetes mellitus: The state of art. J. Clin. Med. 2019, 8, 1843. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. Type 1 Diabetes (T1D) in NOD Mouse Models: The Role of Toll-Like Receptor 7 and an Enteric Bacterial Pathogen in Accelerating the Development of T1D. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2009. [Google Scholar]
- Zhang, J.N. Effect of Polysaccharide from Dendrobium officinale on Human Intestinal Flora In Vitro Fermentation. Master’s Thesis, Xiamen University, Xiamen, China, 2018. [Google Scholar]
- Mccreight, L.J.; Bailey, C.J.; Pearson, E.R. Metformin and the gastrointestinal tract. Diabetologia 2016, 59, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Mariño, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.; Macdougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Lassenius, M.I.; Fogarty, C.L.; Blaut, M.; Haimila, L.; Riittinen, L.; Paju, A.; Kirveskari, J.; Jäerveläe, J.; Järvelä, J.; Ahola, A.J.; et al. Intestinal alkaline phosphatase at the crossroad of intestinal health and disease—A putative role in type 1 diabetes. J. Intern. Med. 2017, 281, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Z.; Zhang, H.H.; Meng, X.Y.; Yi, Y.T. Short chain fatty acids reduced insulin resistance and pancreatic damage in type 2 diabetic mice. Mod. Food Sci. Technol. 2020, 36, 1–7. [Google Scholar]
- Tirosh, A.; Calay, E.S.; Tuncman, G.; Claiborn, K.C.; Inouye, K.E.; Eguchi, K.; Alcala, M.; Rathaus, M.; Hollander, K.S.; Ron, I.; et al. The short-chain fatty acid propionate increases glucagon and fabp4 production, impairing insulin action in mice and humans. Sci. Transl. Med. 2019, 11, eaav0120. [Google Scholar] [CrossRef]
- Xu, H.J.; Hao, X.L.; Qiao, D.L.; Xia, L.B.; Chen, R.; He, X.M.; Gu, F.L.; Wei, C.B. Effects of Dendrobium huoshanense polysaccharides on antioxidant capacity, mucosal barrier integrity and inflammatory responses in an aging rat ileal model. Biotechnol. Biotechnol. Equip. 2019, 33, 1444–1452. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.J.; Zhang, X.F.; Wu, S.H.; Li, S.P.; Si, J.P. Study on quantification method of non-starch polysaccharides in Dendrobium catenatum. Chin. J. Chin. Mater. Med. 2019, 44, 3221–3225. [Google Scholar]
- Staub, A.M. Removal of proteins-sevag method. Methods Carbohydr. Chem. 1965, 5, 5–6. [Google Scholar]
- Xi, X.G.; Wei, X.L.; Wang, Y.F.; Chu, Q.J.; Xiao, J.B. Determination of tea polysaccharides in camellia sinensis by a modified phenol-sulfuric acid method. Arch. Biol. Sci. 2010, 62, 669–672. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. Curr. Protoc. Food Analyt. Chem. 2002, 6, I1.1.1–I1.1.8. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using coomassie brilliant blue g250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Marques, T.M.; O’Sullivan, O.; Fitzgerald, P.; Fitzgerald, G.F.; Cotter, P.D.; Dinan, T.G.; Cryan, J.F.; Stanton, C.; Ross, R.P. Streptozotocin-induced type-1-diabetes disease onset in sprague-dawley rats is associated with an altered intestinal microbiota composition and decreased diversity. Microbiology 2015, 161, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Xie, M.H.; Wan, P.; Chen, D.; Ye, H.; Chen, L.G.; Zeng, X.X.; Liu, Z.H. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from fuzhuan brick tea. Food Chem. 2018, 244, 331–339. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Z.; Huang, F.H.; Yang, C.; Huang, Q.D. In vitro digestion and fermentation by human fecal microbiota of polysaccharides from flax seed. Molecules 2020, 25, 4354. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component 1 | DHWE | DHP | NDHP |
|---|---|---|---|
| Carbohydrates (%) | 82.93 ± 1.10 | 100 ± 1.40 | 61.55 ± 0.81 |
| Total polyphenols (%) | 0.51 ± 0.03 | Not detected | 0.83 ± 0.05 |
| Protein (%) | 0.38 ± 0.06 | Not detected | Not detected |
| Metabolite Types | Peak Area Ratio in DHWE (%) | Peak Area Ratio in NDHP (%) |
|---|---|---|
| Phenylpropanoids and polyketides | 39.3086 | 39.7503 |
| Lipids and lipid-like molecules | 10.8831 | 10.5477 |
| Nucleosides, nucleotides, and analogues | 10.5435 | 11.5194 |
| Organic oxygen compounds | 10.4791 | 9.5488 |
| Organic acids and derivatives | 9.2488 | 8.7881 |
| Organoheterocyclic compounds | 6.9007 | 7.1450 |
| Unclassfied | 6.6155 | 6.3789 |
| Benzenoids | 3.7180 | 4.3710 |
| Lignans, neolignans, and related compounds | 0.7227 | 0.5817 |
| Alkaloids and derivatives | 0.6941 | 0.6642 |
| Hydrocarbon derivatives | 0.0146 | 0.0121 |
| Organic nitrogen compounds | 0.0033 | 0.0033 |
| Group 1 | BASE Group | DHWE Group | DHP Group | NDHP Group | MET Group | |
|---|---|---|---|---|---|---|
| Time (h) | ||||||
| 0 | 7.63 ± 0.06 B | 7.06 ± 0.05 A | 6.84 ± 0.13 A | 7.55 ± 0.16 A | 7.56 ± 0.06 b | |
| 12 | 7.90 ± 0.06 A | 4.65 ± 0.01 B | 4.84 ± 0.09 B | 5.70 ± 0.01 B | 7.71 ± 0.12 ab | |
| 24 | 7.84 ± 0.03 A | 4.64 ± 0.01 B | 4.27 ± 0.01 C | 6.24 ± 0.04 C | 7.80 ± 0.03 a | |
| Group | Fermentation Time (h) | SCFAs Content (μg/mL) 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| Acetic Acid | Propionic Acid | Butyrate | Isobutyric Acid | Valeric Acid | Isovaleric Acid | Total Fatty Acids | ||
| BASE group | 0 | 0.00 ± 0.00 gG | 3.50 ± 0.60 hG | 1.17 ± 0.12 gH | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 1.35 ± 0.05 eE | 6.02 ± 0.60 jI |
| 12 | 17.45 ± 1.25 gG | 41.57 ± 6.38 gF | 6.90 ± 1.41 eDE | 0.03 ± 0.05 eC | 0.00 ± 0.00 B | 0.33 ± 0.06 efE | 66.28 ± 7.81 iH | |
| 24 | 82.45 ± 0.45 fF | 70.60 ± 5.20 fE | 33.37 ± 1.86 bB | 0.60 ± 0.10 dC | 0.00 ± 0.00 B | 29.47 ± 1.70 aA | 216.48 ± 8.35 gF | |
| DHWE group | 0 | 118.90 ± 8.16 eE | 2.07 ± 0.21 hG | 1.27 ± 0.06 gH | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 1.37 ± 0.12 eE | 123.60 ± 8.28 hG |
| 12 | 164.95 ± 2.05 dD | 159.97 ± 16.74 dC | 3.90 ± 0.53 fFGH | 2.10 ± 0.10 cB | 0.00 ± 0.00 B | 3.67 ± 0.40 dD | 334.58 ± 19.47 eE | |
| 24 | 247.87 ± 12.74 bB | 230.33 ± 3.95 bB | 4.80 ± 0.53 fEF | 3.33 ± 0.21 bA | 0.00 ± 0.00 B | 4.55 ± 0.55 dD | 490.88 ± 9.88 cC | |
| DHP group | 0 | 197.95 ± 22.25 cC | 3.53 ± 0.81 hG | 1.25 ± 0.15 gH | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 1.23 ± 0.25 efE | 203.97 ± 23.33 gF |
| 12 | 247.10 ± 23.30 bB | 114.93 ± 11.45 eD | 9.47 ± 1.11 dD | 0.02 ± 0.03 eC | 0.00 ± 0.00 B | 8.37 ± 0.87 cC | 379.88 ± 23.35 dD | |
| 24 | 756.70 ± 30.20 aA | 149.67 ± 6.16 dC | 18.47 ± 1.55 cC | 0.01 ± 0.01 eC | 0.00 ± 0.00 B | 16.37 ± 1.46 bB | 941.21 ± 35.51 aA | |
| NDHP group | 0 | 22.00 ± 1.80 gG | 3.90 ± 0.80 hG | 1.30 ± 0.36 gH | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 0.00 ± 0.00 fE | 27.20 ± 2.86 jHI |
| 12 | 76.90 ± 11.30 fF | 224.83 ± 31.91 bcB | 8.40 ± 0.89 deD | 3.30 ± 0.61 bA | 0.00 ± 0.00 B | 7.40 ± 0.46 cC | 320.83 ± 43.58 efE | |
| 24 | 119.70 ± 3.30 eE | 325.23 ± 13.98 aA | 88.67 ± 3.30 aA | 3.73 ± 0.64 aA | 4.15 ± 0.15 A | 4.80 ± 0.70 dD | 546.28 ± 12.37 bB | |
| MET group | 0 | 0.00 ± 0.00 gG | 2.25 ± 0.25 hG | 1.15 ± 0.05 gH | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 1.15 ± 0.15 efE | 4.55 ± 0.05 jI |
| 12 | 9.50 ± 0.10 gG | 14.70 ± 1.00 hFG | 1.60 ± 0.20 gGH | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 0.02 ± 0.02 fE | 25.82 ± 0.84 jHI | |
| 24 | 78.80 ± 5.80 fF | 207.85 ± 22.65 cB | 4.40 ± 0.10 fEFG | 0.00 ± 0.00 eC | 0.00 ± 0.00 B | 0.08 ± 0.01 efE | 291.13 ± 16.91 fE | |
| Group | CON | BASE Group | DHWE Group | DHP Group | NDHP Group | MET Group |
|---|---|---|---|---|---|---|
| Phylum level | ||||||
| Proteobacteria | 5.49 ± 0.68 dD | 31.5 ± 5.7 bB | 8.4 ± 1.7 dD | 7.1 ± 1.0 dD | 15.9 ± 0.3 cC | 59.9 ± 2.9 aA |
| Firmicutes | 58.66 ± 1.18 bcBC | 34.97 ± 3.26 dD | 66.74 ± 7.91 abAB | 51.37 ± 8.03 cC | 72.29 ± 1.53 aA | 20.97 ± 1.99 eE |
| Bacteroidetes | 35.66 ± 1.80 aAB | 33.41 ± 3.81 abAB | 24.74 ± 9.32 bcBC | 41.19 ± 7.70 aA | 11.71 ± 1.77 dC | 19.0 ± 1.12 cdC |
| Genus level | ||||||
| Lactobacillus | 10.20 ± 1.56 bB | 10.7 ± 0.9 bB | 36.54 ± 5.70 aA | 32.32 ± 4.97 aA | 30.26 ± 4.29 aA | 5.67 ± 0.32 bB |
| Ruminococcus | 6.53 ± 0.64 aA | 0.27 ± 0.15 dC | 0.56 ± 0.16 cdBC | 0.97 ± 0.17 bcBC | 0.29 ± 0.03 dC | 1.34 ± 0.32 bB |
| Megamonas | 7.16 ± 0.43 cC | 3.36 ± 0.25 cdC | 21.72 ± 4.63 bB | 4.00 ± 0.23 cdC | 36.57 ± 2.98 aA | 1.63 ± 0.13 dC |
| Prevotellaceae_Prevotella | 6.99 ± 0.28 cBC | 1.79 ± 2.41 cC | 15.07 ± 8.67 bB | 32.58 ± 5.95 aA | 1.55 ± 0.27 cC | 0.53 ± 0.12 cC |
| Bacteroides | 5.44 ± 0.63 bcB | 20.22 ± 2.38 aA | 4.14 ± 1.4 cB | 5.07 ± 1.75 bcB | 4.78 ± 0.73 bcB | 7.19 ± 1.29 bB |
| Oscillospira | 3.85 ± 0.42 bB | 6.10 ± 0.32 aA | 1.14 ± 0.41 cCD | 1.14 ± 0.40 cCD | 0.41 ± 0.06 dD | 1.65 ± 0.47 cC |
| Phascolarctobacterium | 4.39 ± 0.70 aA | 1.01 ± 0.08 bcB | 0.88 ± 0.12 bcB | 1.20 ± 0.31 bB | 0.71 ± 0.07 bcB | 0.41 ± 0.06 cB |
| Faecalibacterium | 2.58 ± 0.25 aA | 0.16 ± 0.27 cB | 0.48 ± 0.29 bcB | 1.15 ± 1.06 bB | 0.15 ± 0.01 cB | 0.35 ± 0.08 bcB |
| Coprococcus | 2.0 ± 0.09 aA | 0.75 ± 0.44 bB | 0.39 ± 0.26 bcBC | 0.68 ± 0.16 bBC | 0.09 ± 0.01 cC | 0.48 ± 0.12 bcBC |
| Blautia | 1.90 ± 0.12 aA | 0.85 ± 0.06 bcBC | 0.51 ± 0.07 cdBC | 1.17 ± 0.73 bAB | 0.14 ± 0.02 dC | 0.42 ± 0.11 cdBC |
| Paraprevotellaceae-Prevotella | 1.65 ± 0.24 aA | 0.30 ± 0.46 bB | 0.58 ± 0.69 bB | 0.21 ± 0.03 bB | 0.13 ± 0.01 bB | 0.13 ± 0.03 bB |
| Parabacteroides | 1.45 ± 0.13 cdB | 8.64 ± 1.08 aA | 1.4 ± 0.7 cdB | 0.67 ± 0.14 dB | 2.55 ± 0.56 cB | 7.14 ± 1.26 bA |
| Citrobacter | 1.64 ± 0.53 cC | 10.13 ± 1.9 bB | 0.49 ± 0.24 cC | 0.43 ± 0.20 cC | 0.39 ± 0.11 cC | 52.11 ± 4.01 aA |
| Escherichia | 0.48 ± 0.05 cBC | 3.16 ± 0.95 aA | 1.60 ± 0.27 bB | 0.90 ± 0.29 bcBC | 3.72 ± 0.28 aA | 0.25 ± 0.03 cC |
| Klebsiella | 0.35 ± 0.07 cB | 0.96 ± 0.29 bB | 0.30 ± 0.17 cB | 0.77 ± 0.36 bcB | 0.33 ± 0.06 cB | 1.90 ± 0.35 aA |
| Clostridium | 0.36 ± 0.01 aA | 0.04 ± 0.02 cB | 0.27 ± 0.11 abAB | 0.31 ± 0.06 abAB | 0.32 ± 0.24 abAB | 0.12 ± 0.03 bcAB |
| Sutterella | 0.26 ± 0.02 abAB | 0.41 ± 0.23 aA | 0.20 ± 0.12 bAB | 0.11 ± 0.02 bB | 0.06 ± 0.03 bB | 0.08 ± 0.03 bB |
| Dorea | 0.17 ± 0.03 bB | 1.74 ± 0.05 aA | 0.22 ± 0.08 bB | 0.56 ± 0.79 bB | 0.09 ± 0.02 bB | 0.14 ± 0.02 bB |
| Megasphaera | 0.09 ± 0.02 dD | 0.14 ± 0.01 cdBCD | 0.20 ± 0.03 bcBC | 0.21 ± 0.01 bB | 0.80 ± 0.09 aA | 0.09 ± 0.02 dCD |
| Enterocloster | 0.21 ± 0.14 cC | 5.20 ± 0.27 aA | 0.73 ± 0.37 cC | 0.16 ± 0.09 cC | 0.30 ± 0.03 cC | 1.93 ± 1.00 bB |
| Proteus | 0.06 ± 0.01 cB | 9.20 ± 1.84 bA | 0.73 ± 0.21 cB | 1.17 ± 0.12 cB | 0.90 ± 0.19 cB | 7.18 ± 2.19 aA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Liu, Z.; Xu, W.; Zhang, Y. Beneficial In Vitro Effects of Polysaccharide and Non-Polysaccharide Components of Dendrobium huoshanense on Gut Microbiota of Rats with Type 1 Diabetes as Opposed to Metformin. Molecules 2024, 29, 2791. https://doi.org/10.3390/molecules29122791
Xu H, Liu Z, Xu W, Zhang Y. Beneficial In Vitro Effects of Polysaccharide and Non-Polysaccharide Components of Dendrobium huoshanense on Gut Microbiota of Rats with Type 1 Diabetes as Opposed to Metformin. Molecules. 2024; 29(12):2791. https://doi.org/10.3390/molecules29122791
Chicago/Turabian StyleXu, Haijun, Zhu Liu, Wen Xu, and Yafei Zhang. 2024. "Beneficial In Vitro Effects of Polysaccharide and Non-Polysaccharide Components of Dendrobium huoshanense on Gut Microbiota of Rats with Type 1 Diabetes as Opposed to Metformin" Molecules 29, no. 12: 2791. https://doi.org/10.3390/molecules29122791