
Cytotoxicity and Chemotaxonomic Significance of Saponins from Wild and Cultured Asparagus Shoots
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Moisture Content
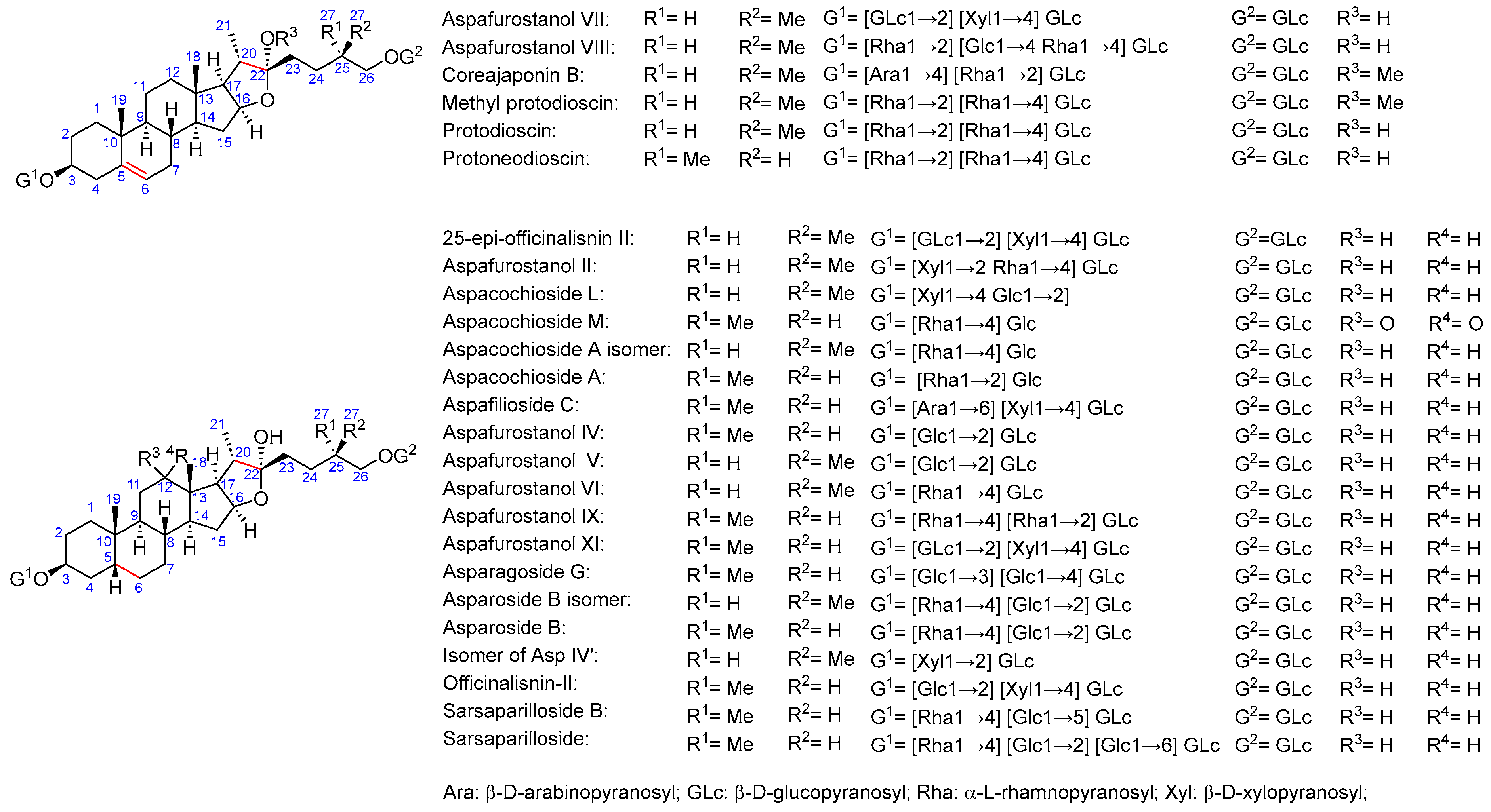
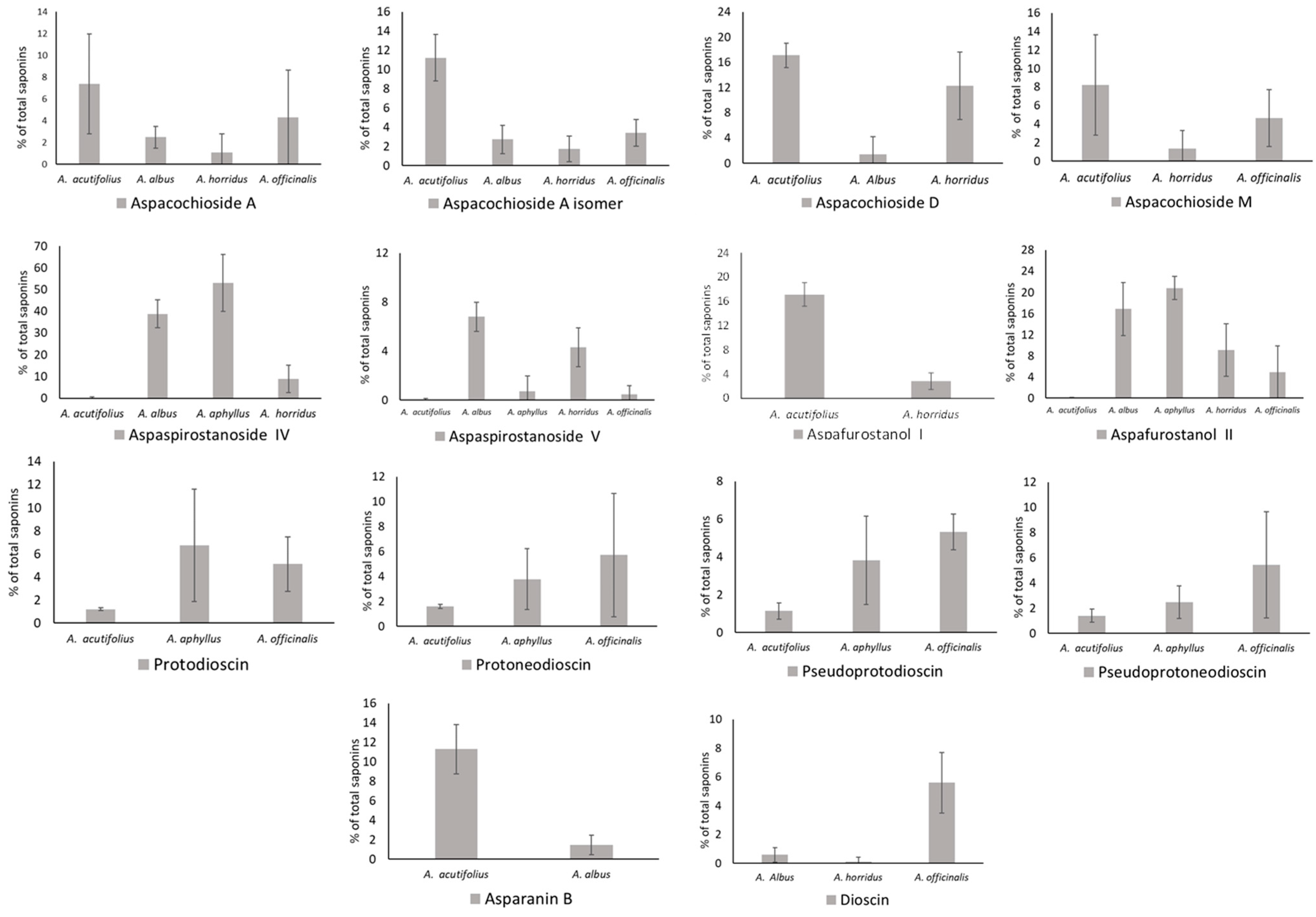
2.2. Total Saponins and Saponin Profiles
2.3. Cluster Analysis
2.4. Antiproliferative Activity
3. Discussion
3.1. Saponin Profiles of the Various Asparagus Species Analyzed in This Work
3.2. Multivariable Analyses for Assessing Chemotaxonomy
3.3. Antiproliferative Activity of the Saponins Extracts of Asparagus Shoots on HT-29 Cancer Cells
4. Materials and Methods
4.1. Samples
4.2. Extraction of Saponins
4.3. Total Saponin Content
4.4. Characterization of Saponins by LC-MS
4.5. Antitumor Assays
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johns, T.; Eyzaguirre, P.B. Linking biodiversity, diet and health in policy and practice. Proc. Nutr. Soc. 2006, 65, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, A.; Price, L.L. Eating and Healing; Haworth Press: London, UK; New York, NY, USA, 2006; pp. 1–384. [Google Scholar]
- Swiderska, K.; Argumedo, A.; Wekesa, C.; Ndalilo, L.; Song, Y.; Rastogi, A.; Ryan, P. Indigenous peoples’ food systems and biocultural heritage: Addressing indigenous priorities using decolonial and interdisciplinary research approaches. Sustainability 2022, 14, 11311. [Google Scholar] [CrossRef]
- Kuhnlein, H.V.; Erasmus, B.; Spigelski, D. Indigenous Peoples’ Food Systems: The Many Dimensions of Culture, Diversity and Environment for Nutrition and Health; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; pp. 1–339. [Google Scholar]
- Adouni, K.; Chahdoura, H.; Mosbah, H.; Santos-Buelga, C.; González-Paramás, A.M.; Ciudad-Mulero, M.; Fernandes, A.; Calhelha, R.C.; Morales, P.; Flamini, G.; et al. Revalorization of wild Asparagus stipularis Forssk. as a traditional vegetable with nutritional and functional properties. Food Funct. 2018, 9, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 471–491. [Google Scholar] [CrossRef]
- Lee, E.J.; Yoo, K.S.; Patil, B.S. Development of a rapid HPLC-UV method for simultaneous quantification of protodioscin and rutin in white and green asparagus spears. J. Food Sci. 2010, 75, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tadmor, Y.; Wu, Q.L.; Chin, C.K.; Garrison, S.A.; Simon, J.E. Quantification of protodioscin and rutin in asparagus shoots by LC/MS and HPLC methods. J. Agric. Food Chem. 2003, 51, 6132–6136. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, A.; Jaramillo-Carmona, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Lachaal, M.; Karray-Bouraoui, N.; Guillén-Bejarano, R. Phytochemical Characterization and Bioactivity of Asparagus acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities. Molecules 2021, 26, 3328. [Google Scholar] [CrossRef] [PubMed]
- Kamat, J.P.; Boloor, K.K.; Devasagayam, T.P.; Venkatachalam, S.R. Antioxidant properties of Asparagus racemosus against damage induced by gamma-radiation in rat liver mitochondria. J. Ethnopharmacol. 2000, 71, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Valdes, B.; Stoycheva, C. Edible Wild Relatives of Cultivated Plants of Popular Use in the Iberian Peninsula. Preprints 2024, 2024061286. [Google Scholar] [CrossRef]
- Marrelli, M.; Conforti, F.; Araniti, F.; Statti, G.A. Effects of saponins on lipid metabolism: A review of potential health benefits in the treatment of obesity. Molecules 2016, 21, 1404. [Google Scholar] [CrossRef]
- Vázquez-Castilla, S.; De la Puerta, R.; Garcia-Gimenez, M.; Fernández-Arche, M.; Guillén-Bejarano, R. Bioactive constituents from “Triguero” asparagus improve the plasma lipid profile and liver antioxidant status in hypercholesterolemic rats. Int. J. Mol. Sci. 2013, 14, 21227–21239. [Google Scholar] [CrossRef] [PubMed]
- Chileh Chelh, T.; Rincon-Cervera, M.A.; Gomez-Mercado, F.; Lopez-Ruiz, R.; Gallon-Bedoya, M.; Ezzaitouni, M.; Guil-Guerrero, J.L. Wild Asparagus Shoots Constitute a Healthy Source of Bioactive Compounds. Molecules 2023, 28, 5786. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Poobrasert, O.; Kennelly, E.J.; Chin, C.; Ho, C.; Huang, M.; Garrison, S.A.; Cordell, G.A. Cytotoxic activity of steroidal saponins from Asparagus officinalis. In Proceedings of the IX International Asparagus Symposium 479, Pasco, WA, USA, 15–17 July 1997; pp. 277–282. [Google Scholar]
- Vázquez-Castilla, S.; Jaramillo-Carmona, S.; Fuentes-Alventosa, J.M.; Jiménez-Araujo, A.; Rodriguez-Arcos, R.; Cermeño-Sacristán, P.; Espejo-Calvo, J.A.; Guillén-Bejarano, R. Optimization of a method for the profiling and quantification of saponins in different green asparagus genotypes. J. Agric. Food Chem. 2013, 61, 6250–6258. [Google Scholar] [CrossRef] [PubMed]
- Pegiou, E.; Mumm, R.; Acharya, P.; de Vos, R.C.H.; Hall, R.D. Green and White Asparagus (Asparagus officinalis): A Source of Developmental, Chemical and Urinary Intrigue. Metabolites 2020, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Norup, M.F.; Petersen, G.; Burrows, S.; Bouchenak-Khelladi, Y.; Leebens-Mack, J.; Pires, J.C.; Linder, H.P.; Seberg, O. Evolution of Asparagus L. (Asparagaceae): Out-of-South-Africa and multiple origins of sexual dimorphism. Mol. Phylogenet. Evol. 2015, 92, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.M.; Nahar, L.; Mannan, A.; Arfan, M.; Khan, G.A.; Al-Groshi, A.; Evans, A.; Dempster, N.M.; Ismail, F.M.D.; Sarker, S.D. Liquid chromatography mass spectrometry analysis and cytotoxicity of Asparagus adscendens roots against human cancer cell lines. Pharmacogn. Mag. 2017, 13, S890–S894. [Google Scholar]
- Liu, W.; Ning, R.; Chen, R.N.; Huang, X.F.; Dai, Q.S.; Hu, J.H.; Wang, Y.W.; Wu, L.L.; Xiong, J.; Hu, G.; et al. Aspafilioside B induces G2/M cell cycle arrest and apoptosis by up-regulating H-Ras and N-Ras via ERK and p38 MAPK signaling pathways in human hepatoma HepG2 cells. Mol. Carcinog. 2015, 55, 440–457. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, A.; Jaramillo-Carmona, S.; Srairi Beji, R.; Tej, R.; Zaoui, S.; Rodriguez-Arcos, R.; Guillen-Bejarano, R. The phytochemical and bioactivity profiles of wild Asparagus albus L. plant. Food Res. Int. 2017, 99, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, Y.-Y.; Sun, Y.-S.; Ma, R.-H.; Thakur, K.; Zhang, J.-G.; Wei, Z.-J. Asparanin A from Asparagus officinalis L. Induces G0/G1 Cell Cycle Arrest and Apoptosis in Human Endometrial Carcinoma Ishikawa Cells via Mitochondrial and PI3K/AKT Signaling Pathways. J. Agric. Food Chem. 2020, 68, 213–224. [Google Scholar] [CrossRef]
- Liu, W.; Huang, X.F.; Qi, Q.; Dai, Q.S.; Yang, L.; Nie, F.F.; Lu, N.; Gong, D.D.; Kong, L.Y.; Guo, Q.L. Asparanin A induces G2/M cell cycle arrest and apoptosis in human hepatocellular carcinoma HepG2 cells. Biochem. Biophys. Res. Commun. 2009, 381, 700–705. [Google Scholar] [CrossRef]
- Bousserouel, S.; Le Grandois, J.; Gosse, F.; Werner, D.; Barth, S.W.; Marchioni, E.; Raul, F. Methanolic extract of white asparagus shoots activates TRAIL apoptotic death pathway in human cancer cells and inhibits colon carcinogenesis in a preclinical model. Int. J. Oncol. 2013, 43, 394–404. [Google Scholar] [CrossRef]
- Jaramillo, S.; Muriana, F.; Guillen, R.; Jimenez-Araujo, A.; Rodriguez-Arcos, R.; Lopez, S. Saponins from edible spears of wild asparagus inhibit AKT, p70S6K, and ERK signalling, and induce apoptosis through G0/G1 cell cycle arrest in human colon cancer HCT-116 cells. J. Funct. Foods 2016, 26, 1–10. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Zhao, J.; Zhang, W.; Pang, X. Saponins extracted from by-product of Asparagus officinalis L. suppress tumour cell migration and invasion through targeting Rho GTPase signalling pathway. J. Sci. Food Agric. 2013, 93, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Sunb, Q.Y.; Ge, Z.J. Therapeutic effects of ginseng and ginsenosides on colorectal cancer. Food Funct. 2022, 13, 6450. [Google Scholar] [CrossRef]
- Jaramillo-Carmona, S.; Guillen-Bejarano, R.; Jimenez-Araujo, A.; Rodriguez-Arcos, R.; Lopez, S. In Vitro Toxicity of Asparagus Saponins in Distinct Multidrug-Resistant Colon Cancer Cells. Chem. Biodivers. 2018, 15, e1800282. [Google Scholar] [CrossRef]
- Yang, L.; Ren, S.; Xu, F.; Ma, Z.; Liu, X.; Wang, L. Recent advances in the pharmacological activities of dioscin. Hindawi BioMed. Res. Int. 2019, 2019, 5763602. [Google Scholar] [CrossRef]
- Mitra, S.K.; Prakash, N.S.; Sundaram, R. Shatavarins (containing Shatavarin IV) with anticancer activity from the roots of Asparagus racemosus. Indian J. Pharmacol. 2012, 44, 732. [Google Scholar] [PubMed]
- Boik, J.C. Natural Compounds in Cancer Therapy; Oregon Medical Press: Princeton, MN, USA, 2001; pp. 149–190. [Google Scholar]
- Stefanowicz-Hajduk, J.; Król-Kogus, B.; Sparzak-Stefanowska, B.; Kimel, K.; Ochocka, J.R.; Krauze-Baranowska, M. Cytotoxic activity of standardized extracts, a fraction, and individual secondary metabolites from fenugreek seeds against SKOV-3, HeLa, and MOLT-4 cell lines. Pharm. Biol. 2021, 59, 424–437. [Google Scholar] [CrossRef]
- Suffness, M.; Pezzuto, J.M. Assays related to cancer drug discovery. In Methods in Plant Biochemistry: Assays for Bioactivity; Hostettmann, K., Ed.; Academic Press: London, UK, 1991; pp. 71–133. [Google Scholar]
- Xiang, J.; Xiang, Y.; Lin, S.; Xin, D.; Liu, X.; Weng, L.; Chen, T.; Zhang, M. Anticancer effects of deproteinized asparagus polysaccharide on hepatocellular carcinoma in vitro and in vivo. Tumour Biol. 2014, 35, 3517–3524. [Google Scholar] [CrossRef]
- Le Bot, M.; Thibault, J.; Pottier, Q.; Boisard, S.; Guilet, D. An accurate, cost-effective and simple colorimetric method for the quantification of total triterpenoid and steroidal saponins from plant materials. Food Chem. 2022, 383, 132597. [Google Scholar] [CrossRef]
- Ncube, B.; Ngunge, V.; Finnie, J.; Van Staden, J. A comparative study of the antimicrobial and phytochemical properties between outdoor grown and micropropagated Tulbaghia violacea Harv. plants. J. Ethnopharmacol. 2011, 134, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Lyashenko, S.; González-Fernández, M.J.; Gómez-Mercado, F.; Yunusova, S.; Denisenko, O.; Guil-Guerrero, J.L. Ribes taxa: A promising source of γ-linolenic acid-rich functional oils. Food Chem. 2019, 301, 125309. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Bueno, R.P.; Romero-González, R.; González-Fernández, M.J.; Guil-Guerrero, J.L. Phytochemical composition and in vitro anti-tumour activities of selected tomato varieties. J. Sci. Food Agric. 2017, 97, 488–496. [Google Scholar] [CrossRef]
- Vichitsakul, K.; Laowichuwakonnukul, K.; Soontornworajit, B.; Poomipark, N.; Itharat, A.; Rotkrua, P. Anti-proliferation and induction of mitochondria-mediated apoptosis by Garcinia hanburyi resin in colorectal cancer cells. Heliyon 2023, 9, e16411. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Samples | Moisture (g/100 g) | Saponins (mg/100 g Dry Weight) |
|---|---|---|
| A. acutifolius | ||
| AC1 | 81.4 ± 0.0 h | 1094.9 ± 21.2 def |
| AC2 | 84.6 ± 0.3 fg | 1529.3 ± 107.8 a |
| AC3 | 85.2 ± 0.3 ef | 1151.5 ± 7.9 de |
| Mean ± SD | 84.5 ± 2.3 B | 1258.6 ± 236.2 A |
| A. albus | ||
| AL1 | 88.0 ± 0.2 bc | 1405.2 ± 176.3 ab |
| AL2 | 88.1 ± 0.2 bc | 996.9 ± 39.1 efg |
| AL3 | 86.4 ± 1.2 de | 1387.4 ± 107.9 abc |
| AL4 | 89.0 ± 0.5 b | 930.5 ± 98.5 fgh |
| Mean ± SD | 87.9 ± 1.1 AB | 1180.0 ± 251.3 AB |
| A. aphyllus | ||
| AP1 | 83.7 ± 0.9 g | 907.8 ± 27.3 fgh |
| AP2 | 85.3 ± 0.5 ef | 860.8 ± 115.0 gh |
| AP3 | 86.5 ± 1.2 de | 909.3 ± 7.7 fgh |
| Mean ± SD | 85.2 ± 1.4 B | 892.6 ± 27.6 AB |
| A. horridus | ||
| H1 | 80.7 ± 1.4 h | 1211.5 ± 60.3 cd |
| H2 | 84.0 ± 0.7 fg | 1422.9 ± 70.8 ab |
| H3 | 83.3 ± 1.1 g | 838.3 ± 174.1 ghi |
| H4 | 87.5 ± 0.1 cd | 838.1 ± 55.5 ghi |
| H5 | 87.3 ± 0.5 cd | 794.9 ± 31.2 hi |
| Mean ± SD | 84.6 ± 2.9 B | 1021.1 ± 281.0 AB |
| A. officinalis | ||
| O1 | 91.1 ± 0.2 a | 750.9 ± 112.0 hi |
| O2 | 90.9 ± 0.3 a | 669.1 ± 101.0 i |
| Mean ± SD | 91.0 ± 0.1 A | 710.0 ± 57.8 B |
| Species/Location | Code | Geographical Coordinates | Date |
|---|---|---|---|
| A. acutifolius (Raviscanina) | |||
| Punta Entinas, El Ejido, Almería | AC1 | 36.690831, −2.7732501 | 04 April 2023 |
| Algámitas, Sevilla | AC2 | 37.018445, −5.161181 | 07 March 2023 |
| Fonelas, Granada | AC3 | 37.409918, −3.200831 | 10 May 2023 |
| A. albus (White asparagus) | |||
| Darrícal, Almería | AL1 | 36.917674, −3.028230 | 17 March 2023 |
| Sierra Cabrera, Almería | AL2 | 37.134984, −1.868005 | 05 January 2023 |
| Barranco de las Lastras, Adra, Almería | AL3 | 36.790886, −3.100039 | 03 March 2023 |
| El Toyo, Almería | AL4 | 36.847975, −2.332920 | 03 February 2023 |
| A. aphyllus (Prickly asparagus) | |||
| Zahara de la Sierra, Cádiz | AP1 | 36.841757, −5.395525 | 20 March 2023 |
| Mijas, Málaga | AP2 | 36.591142, −4.606242 | 23 March 2023 |
| Torremolinos, Málaga | AP3 | 36.605964, −4.526782 | 20 March 2023 |
| A. horridus (Esparraguera) | |||
| Cabo de Gata, Almería | H1 | 36.723495, −2.183220 | 19 March 2023 |
| Vícar, Almería | H2 | 36.813587, −2.60462 | 11 March 2023 |
| Enix, Almería | H3 | 36.875594, −2.609560 | 12 March 2023 |
| Las Amoladeras Almería | H4 | 36.817729, −2.253485 | 07 March 2023 |
| Rodalquilar, Níjar | H5 | 36.849231, −2.043093 | 14 February 2023 |
| A. officinalis (Garden asparagus) | |||
| Láchar, Granada | O1 | Purchased | 01 April 2023 |
| Loja, Granada | O2 | Purchased | 05 October 2023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chileh-Chelh, T.; López-Ruiz, R.; García-Cervantes, A.M.; Rodríguez-García, I.; Rincón-Cervera, M.A.; Ezzaitouni, M.; Guil-Guerrero, J.L. Cytotoxicity and Chemotaxonomic Significance of Saponins from Wild and Cultured Asparagus Shoots. Molecules 2024, 29, 3367. https://doi.org/10.3390/molecules29143367
Chileh-Chelh T, López-Ruiz R, García-Cervantes AM, Rodríguez-García I, Rincón-Cervera MA, Ezzaitouni M, Guil-Guerrero JL. Cytotoxicity and Chemotaxonomic Significance of Saponins from Wild and Cultured Asparagus Shoots. Molecules. 2024; 29(14):3367. https://doi.org/10.3390/molecules29143367
Chicago/Turabian StyleChileh-Chelh, Tarik, Rosalía López-Ruiz, Ana M. García-Cervantes, Ignacio Rodríguez-García, Miguel A. Rincón-Cervera, Mohamed Ezzaitouni, and José L. Guil-Guerrero. 2024. "Cytotoxicity and Chemotaxonomic Significance of Saponins from Wild and Cultured Asparagus Shoots" Molecules 29, no. 14: 3367. https://doi.org/10.3390/molecules29143367