
Impact of Processing on the Phenolic Content and Antioxidant Activity of Sorghum bicolor L. Moench

 ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results and Discussion
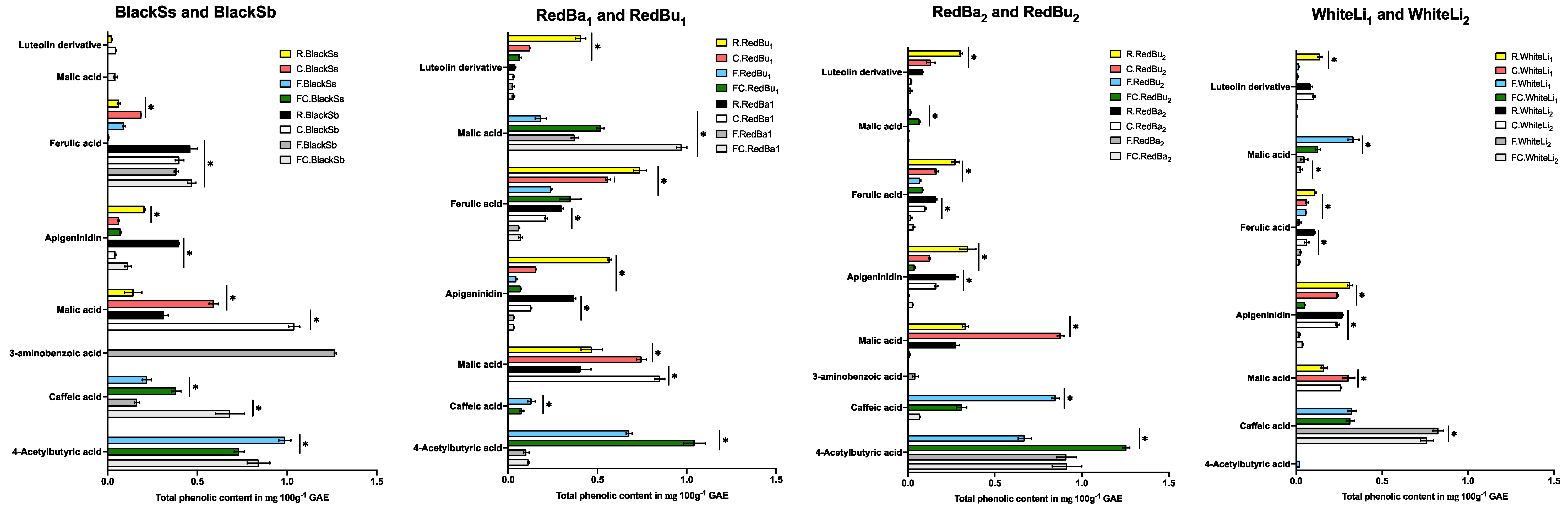
2.1. Impact of Processing on Total Phenolic Content (TPC)
2.2. Total Proanthocyanidin Content (TPAC)
2.3. Post Processing Variation in Antioxidant Activity
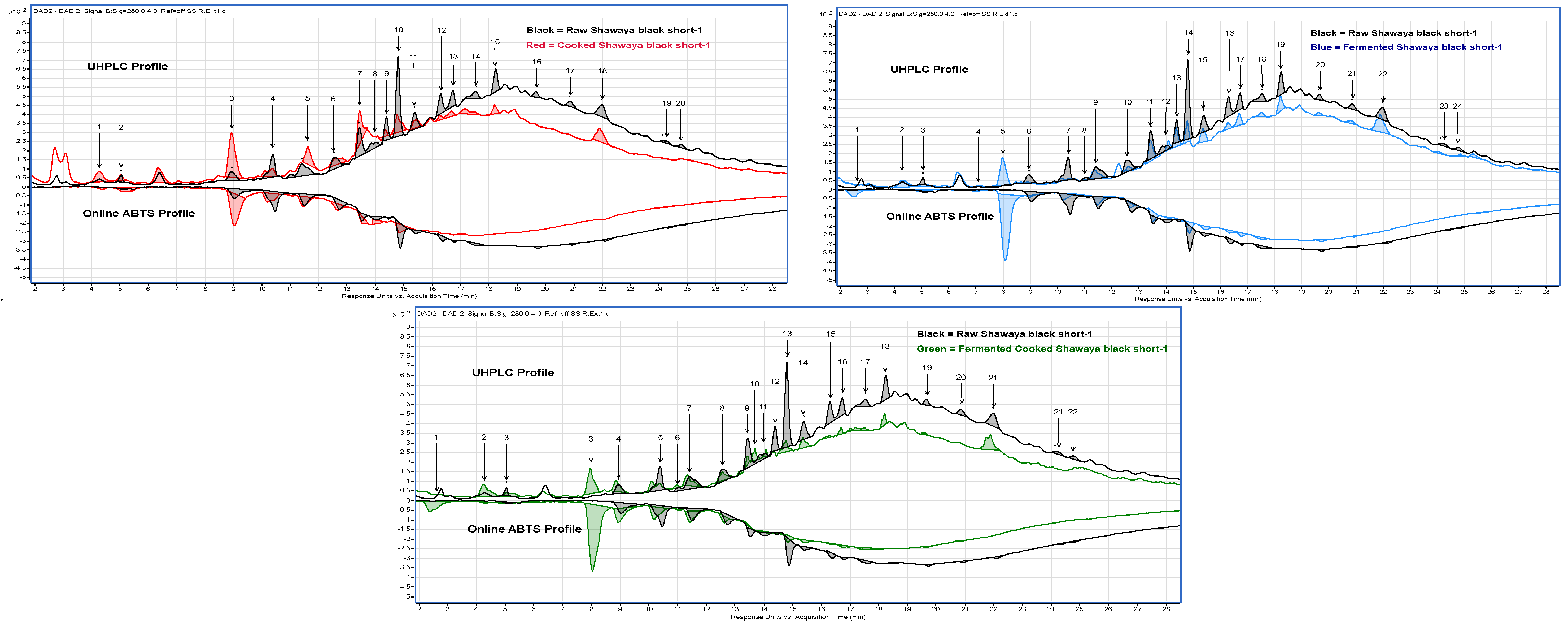
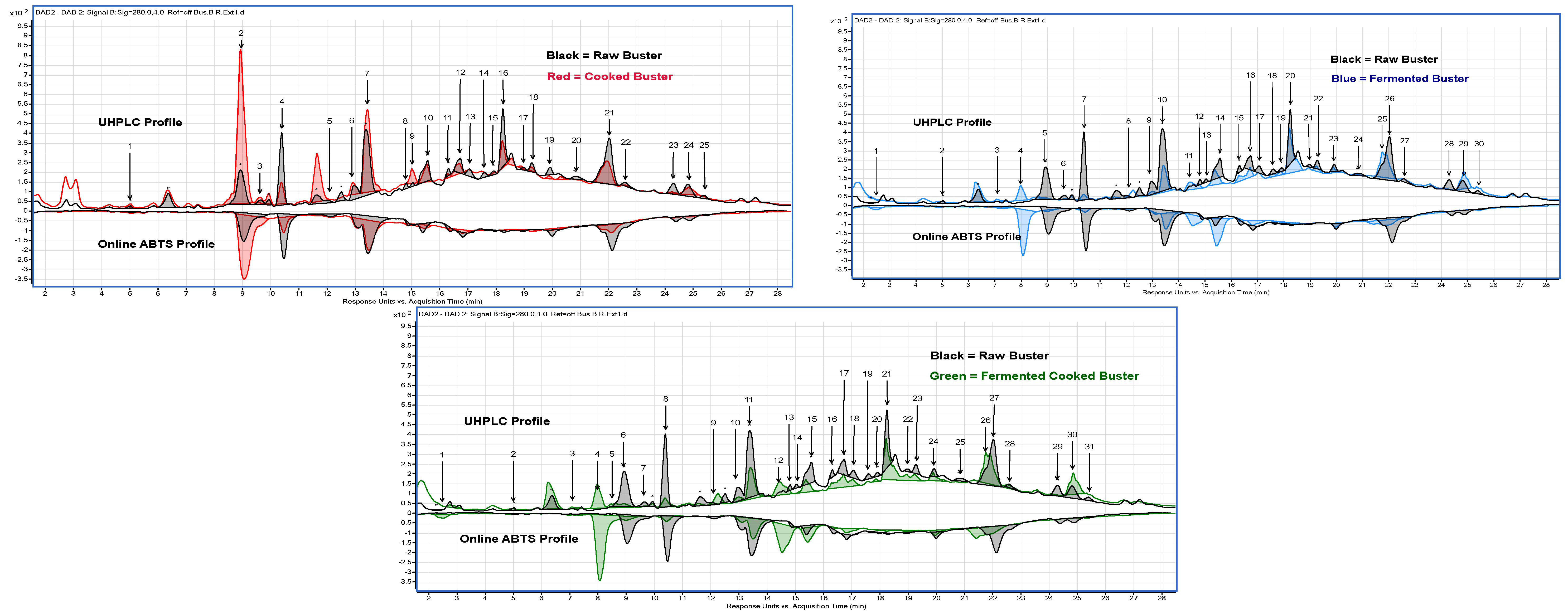
2.4. Quantification and Identification Using UHPLC with Online ABTS and QTOF LC-MS
3. Materials and Methods
3.1. Materials
3.1.1. Samples
3.1.2. Standards and Reagents
3.2. Methods
3.2.1. Flour Preparation
3.2.2. Cooking of Sorghum Flour
3.2.3. Fermentation of Sorghum Flour
3.2.4. Cooking of Fermented Sorghum Flour
3.2.5. Polyphenol Extraction
3.2.6. Total Phenolic Content
3.2.7. Total Proanthocyanidin Content (TPAC)
3.2.8. DPPH Radical Scavenging Activity
3.2.9. Ferric Reducing Ability of Plasma (FRAP) Assay
3.2.10. ABTS Radical Scavenging Activity
3.2.11. UHPLC with Online ABTS
3.2.12. Compound Identification Using QTOF LC-MS
3.2.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Li, Z.; Zhao, X.; Zhang, X.; Liu, H. Bioactive compounds and biological activities of sorghum grains. Foods 2021, 10, 2868. [Google Scholar] [CrossRef] [PubMed]
- Salih, S.A.; Ahmed, K.E.; Ezzdeen, L.T.; Hamza, A.A. Effect of fermentation and processing of Sorghum bicolor grains to produce traditional Sudanese hulu-muron phytochemicals and their biological activities. GSC Biol. Pharm. Sci. 2020, 10, 76–84. [Google Scholar] [CrossRef]
- Brantsen, J.F.; Herrman, D.A.; Ravisankar, S.; Awika, J.M. Effect of tannins on microwave-assisted extractability and color properties of sorghum 3-deoxyanthocyanins. Food Res. Int. 2021, 148, 963–9969. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Chinkwo, K.; Santhakumar, A.; Johnson, S.; Blanchard, C. Apoptosis induction pathway in human colorectal cancer cell line SW480 exposed to cereal phenolic extracts. Molecules 2019, 24, 2465. [Google Scholar] [CrossRef] [PubMed]
- Francis, N.; Rao, S.; Blanchard, C.; Santhakumar, A. Black sorghum phenolic extract regulates expression of genes associated with oxidative stress and inflammation in human endothelial cells. Molecules 2019, 24, 3321. [Google Scholar] [CrossRef] [PubMed]
- Callcott, E.T.; Blanchard, C.L.; Snell, P.; Santhakumar, A.B. The anti-inflammatory and antioxidant effects of pigmented rice consumption in an obese cohort. Food Funct. 2019, 10, 8016–8025. [Google Scholar] [CrossRef]
- Collins, A.; Santhakumar, A.B.; Francis, N.; Blanchard, C.; Chinkwo, K. Impact of sorghum (Sorghum bicolor L. Moench) phenolic compounds on cancer development pathways. Food Biosci. 2024, 59, 104177. [Google Scholar] [CrossRef]
- Misra, D.; Dutta, W.; Jha, G.; Ray, P. Interactions and regulatory functions of phenolics in soil-plant-climate nexus. Agronomy 2023, 13, 280. [Google Scholar] [CrossRef]
- Hithamani, G.; Srinivasan, K. Bioaccessibility of polyphenols from wheat (Triticum aestivum), sorghum (Sorghum bicolor), green gram (Vigna radiata), and chickpea (Cicer arietinum) as influenced by domestic food processing. J. Agric. Food Chem. 2014, 62, 11170–11179. [Google Scholar] [CrossRef]
- Khoddami, A.; Mohammadrezaei, M.; Roberts, T.H. Effects of sorghum malting on colour, major classes of phenolics and individual anthocyanins. Molecules 2017, 22, 1713. [Google Scholar] [CrossRef]
- Dlamini, N.R.; Taylor, J.R.N.; Rooney, L.W. The effect of sorghum type and processing on the antioxidant properties of African sorghum-based foods. Food Chem. 2007, 105, 1412–1419. [Google Scholar] [CrossRef]
- Adebo, O.A. African sorghum-based fermented foods: Past, current and future prospects. Nutrients 2020, 12, 1111. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.; Patel, A.S.; Kar, A.; Deshpande, S.S.; Tripathi, M.K. Effect of different processing conditions on proximate composition, anti-oxidants, anti-nutrients and amino acid profile of grain sorghum. Food Chem. 2019, 271, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, L.; Yang, T.; Ma, Y.; McClements, D.J.; Ren, F.; Tian, Y.; Jin, Z. A review of structural transformations and properties changes in starch during thermal processing of foods. Food Hydrocoll. 2021, 113, 106543. [Google Scholar] [CrossRef]
- Martínez, M.M.; Gómez, M. Rheological and microstructural evolution of the most common gluten-free flours and starches during bread fermentation and baking. J. Food Eng. 2017, 197, 78–86. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Zhang, X.; Liu, H. The effects of processing on bioactive compounds and biological activities of sorghum grains. Molecules 2022, 27, 3246. [Google Scholar] [CrossRef] [PubMed]
- Susanti, D.Y.; Sediawan, W.B.; Fahrurrozi, M.; Hidayat, M.; Putri, A.Y. Encapsulation of red sorghum extract rich in proanthocyanidins: Process formulation and mechanistic model of foam-mat drying at various temperature. Chem. Eng. Process.-Process Intensif. 2021, 164, 108375. [Google Scholar] [CrossRef]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed]
- Duodu, K.; Taylor, J.; Belton, P.; Hamaker, B. Factors affecting sorghum protein digestibility. J. Cereal Sci. 2003, 38, 117–131. [Google Scholar] [CrossRef]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic compositions and antioxidant activities differ significantly among sorghum grains with different applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Blanchard, C.L. Investigation of phenolic compounds with antioxidant activity in barley and oats affected by variation in growing location. Cereal Chem. 2020, 97, 772–782. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Metwally, A.S.; Nassan, M.A.; Abdallah, K.; Eldemery, F.; Tufarelli, V.; Laudadio, V.; Kishawy, A.T.Y. Potential application of cornelian cherry extract on broiler chickens: Growth, expression of antioxidant biomarker and glucose transport genes, and oxidative stability of frozen meat. Animals 2021, 11, 1038. [Google Scholar] [CrossRef] [PubMed]
- Dia, V.P.; Pangloli, P.; Jones, L.; McClure, A.; Patel, A. Phytochemical concentrations and biological activities of Sorghum bicolor alcoholic extracts. Food Funct. 2016, 7, 3410–3420. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.C.; Johnson, S.K.; Bornman, J.F.; Bennett, S.J.; Fang, Z.X. Changes in whole grain polyphenols and antioxidant activity of six sorghum genotypes under different irrigation treatments. Food Chem. 2017, 214, 199–207. [Google Scholar] [CrossRef]
- Awika, J.; Taylor, J.R.N. Gluten-Free Ancient Grains; Woodhead Publishing: Sawston, UK, 2017; pp. 21–54. [Google Scholar]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum grain: From genotype, nutrition, and phenolic profile to its health benefits and food applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [PubMed]
- Punia, H.; Tokas, J.; Malik, A.; Sangwan, S.; Sangwan, S. Characterization of phenolic compounds and antioxidant activity in sorghum [Sorghum bicolor (L.) Moench] grains. Cereal Res. Commun. 2021, 49, 343–353. [Google Scholar] [CrossRef]
- D‘Almeida, C.; Mameri, H.; Menezes, N.; dos Santos Menezes, N.; de Carvalho, C.W.; Queiroz, C.; Morel, M.; Takeiti, C.; Ferreira, M. Effect of extrusion and turmeric addition on phenolic compounds and kafirin properties in tannin and tannin-free sorghum. Food Res. Int. 2021, 149, 110663. [Google Scholar] [CrossRef] [PubMed]
- Udachan, I.S.; Sahu, A.K.; Hend, F.M. Extraction and characterization of sorghum (Sorghum Bicolor L. Moench) starch. Int. Food Res. J. 2012, 19, 315–319. [Google Scholar]
- Syeunda, C.; Awika, J.M. Mechanisms of flavonoid inhibition of Maillard reaction product formation in relation to whole grains processing. Food Chem. 2024, 449, 139237. [Google Scholar] [CrossRef]
- Bianco-Gomes, A.C.; Nogueira, L.D.S.; Bono-Lopes, N.V.M.; Gouvêa-Souza, C.P.; Boldrini-França, J.; Gomes, V.M.; Cherene, M.B.; Alves, N.E.G.; Vasconcelos, C.M. Dry heat and pressure favor bioactive compounds preservation and peptides formation in sorghum [Sorghum bicolor (L.) Moench]. Curr. Res. Food Sci. 2022, 5, 117–124. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Adebiyi, J.A.; Kayitesi, E. Co-influence of fermentation time and temperature on physicochemical properties, bioactive components and microstructure of ting (a Southern African food) from whole grain sorghum. Food Biosci. 2018, 25, 118–127. [Google Scholar] [CrossRef]
- Zaroug, M.; Orhan, I.E.; Senol, F.S.; Yagi, S. Comparative antioxidant activity appraisal of traditional Sudanese kisra prepared from two sorghum cultivars. Food Chem. 2014, 156, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Moreno, C.R.; Fernández, P.C.R.; Rodríguez, E.O.C.; Carrillo, J.M.; Rochín, S.M. Extrusion of Metals, Polymers and Food Products; IntechOpen: London, UK, 2018; pp. 104–124. [Google Scholar]
- Alazzouni, A.; Dkhil, M.; Gadelmawla, M.; Gabri, M.; Farag, A.; Hassan, B. Ferulic acid as anticarcinogenic agent against 1,2-dimethylhydrazine induced colon cancer in rats. J. King Saud Univ. Sci. 2021, 33, 101354. [Google Scholar] [CrossRef]
- Khan, A.; Khan, R.; Qamar, W.; Lateef, M.; Ali, F.; Tahir, M.; Muneeb, U.; Sultana, S. Caffeic acid attenuates 12-O-tetradecanoyl-phorbol-13-acetate (TPA)-induced NF-kB and COX-2 expression in mouse skin: Abrogation of oxidative stress, inflammatory responses and proinflammatory cytokine production. Food Chem. Toxicol. 2012, 50, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Proietti, I.; Mantovani, A.; Mouquet-Rivier, C.; Guyot, J.P. Modulation of chelating factors, trace minerals and their estimated bioavailability in Italian and African sorghum (Sorghum bicolor (L.) Moench) porridges. Int. J. Food Sci. Technol. 2013, 48, 1526–1532. [Google Scholar] [CrossRef]
- Correia, I.; Nunes, A.; Barros, A.S.; Delgadillo, I. Comparison of the effects induced by different processing methods on sorghum proteins. J. Cereal Sci. 2010, 51, 146–151. [Google Scholar] [CrossRef]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Wu, G.; Johnson, S.K.; Blanchard, C.L. Characterization of phenolic compounds and antioxidant activity in sorghum grains. J. Cereal Sci. 2018, 84, 103–111. [Google Scholar] [CrossRef]
- Noreen, H.; Semmar, N.; Farman, M.; McCullagh, J.S.O. Measurement of total phenolic content and antioxidant activity of aerial parts of medicinal plant Coronopus didymus. Asian Pac. J. Trop. Med. 2017, 10, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; McClung, A.M.; Chen, M.H. Phytochemicals and antioxidant capacities in rice brans of different color. J. Food Sci. 2011, 76, C117–C126. [Google Scholar] [CrossRef]
- Zhu, Y.; Shi, Z.; Yao, Y.; Hao, Y.; Ren, G. Antioxidant and anti-cancer activities of proanthocyanidins-rich extracts from three varieties of sorghum (Sorghum bicolor) bran. Food Agric. Immunol. 2017, 28, 1530–1543. [Google Scholar] [CrossRef]
- Sompong, R.; Siebenhandl-Ehn, S.; Linsberger-Martin, G.; Berghofer, E. Physicochemical and antioxidative properties of red and black rice varieties from Thailand, China and Sri Lanka. Food Chem. 2011, 124, 132–140. [Google Scholar] [CrossRef]
- Woo, K.S.; Ko, J.Y.; Jeong, H.S. Effect of milling time on antioxidant compounds and activities of methanol extracts of sorghum [Sorghum bicolor (L.) Moench]. Food Sci. Biotechnol. 2014, 23, 1741–1746. [Google Scholar] [CrossRef]
- García-Villalba, R.; Espín, J.C.; Tomás-Barberán, F.A. Chromatographic and spectroscopic characterization of urolithins for their determination in biological samples after the intake of foods containing ellagitannins and ellagic acid. J. Chromatogr. A 2016, 1428, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Ed Nignpense, B.; Chinkwo, K.A.; Blanchard, C.L.; Santhakumar, A.B. Black sorghum phenolic extract modulates platelet activation and platelet microparticle release. Nutrients 2020, 12, 1760. [Google Scholar] [CrossRef]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Blanchard, C.L. Q-TOF LC/MS identification and UHPLC-Online ABTS antioxidant activity guided mapping of barley polyphenols. Food Chem. 2018, 266, 323–328. [Google Scholar] [CrossRef]
- ChemSpider. In Royal Society of Chemistry. 2024. Available online: https://www.chemspider.com/Search.aspx (accessed on 3 February 2023).
- PubChem. In National Center for Biotechnology Information. 2024. Available online: https://pubchem.ncbi.nlm.nih.gov/search/#collection=compounds (accessed on 3 February 2023).
- MassBank. MassBank Project. 2006. Available online: https://massbank.eu/MassBank/Search (accessed on 3 February 2023).
- Metlin. 10550 North Torrey Pines Road BCC-007, La Jolla, CA 92037 USA, 2006. Available online: https://metlin.scripps.edu (accessed on 3 February 2023).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Processing Technique | TPC mg/g GAE | TPAC mg/g CE | ABTS mg 100 g−1 TE | DPPH mg/g TE | FRAP mg/g TE |
|---|---|---|---|---|---|---|
| BlackSs (Shawaya short black-1) | Raw | 9.66 ± 1.66 a | 8.38 ± 0.44 a | 2.07 ± 0.22 a | 11.10 ± 0.06 a | 23.27 ± 3.50 a |
| Cooked | 7.38 ± 0.53 b | 4.88 ± 0.67 b | 1.47 ± 0.25 b | 10.8 ± 0.09 b | 28.57 ± 2.22 b | |
| Fermented | 7.93 ± 1.67 c | 6.44 ± 0.30 c | 1.52 ± 0.08 bc | 10.70 ± 0.25 bc | 15.80 ± 1.24 c | |
| Fermented Cooked | 10.93 ± 0.85 d | 7.05 ± 0.58 c | 2.25 ± 0.09 a | 11.30 ± 0.06 a | 26.81 ± 3.63 ab | |
| BlackSb (Shawaya black) | Raw | 6.67 ± 0.64 a | 2.01 ± 0.08 a | 0.92 ± 0.04 a | 2.56 ± 0.14 ab | 7.29 ± 1.26 a |
| Cooked | 5.97 ± 0.61 ab | 2.46 ± 0.21 a | 1.04 ± 0.12 b | 2.76 ± 0.04 c | 9.13 ± 1.55 a | |
| Fermented | 6.73 ± 0.77 a | 2.45 ± 0.14 a | 1.14 ± 0.07 bc | 2.61 ± 0.07 ad | 11.98 ± 2.00 b | |
| Fermented Cooked | 6.86 ± 0.73 ac | 2.65 ± 0.19 b | 1.36 ± 0.07 d | 2.75 ± 0.08 ce | 12.37 ± 1.98 bc | |
| RedBu1 (Buster.B) | Raw | 0.67 ± 0.08 a | 1.31 ± 0.25 a | 0.87 ± 0.24 a | 2.40 ± 0.19 a | 5.09 ± 0.62 a |
| Cooked | 0.57 ± 0.06 b | 1.18 ± 0.18 a | 0.91 ± 0.17 a | 2.40 ± 0.18 a | 5.81 ± 1.18 a | |
| Fermented | 0.52 ± 0.06 bc | 1.40 ± 0.33 a | 0.86 ± 0.04 a | 1.99 ± 0.17 b | 9.88 ± 1.08 b | |
| Fermented Cooked | 0.58 ± 0.06 bd | 1.44 ± 0.25 a | 1.00 ± 0.09 a | 2.33 ± 0.12 ac | 10.49 ± 0.94 bc | |
| RedBa1 (Bazley.B) | Raw | 0.50 ± 0.13 a | 1.27 ± 0.07 a | 0.89 ± 0.06 a | 2.29 ± 0.18 a | 6.69 ± 0.66 a |
| Cooked | 0.67 ± 0.25 ac | 1.31 ± 0.30 a | 1.07 ± 0.20 a | 2.61 ± 0.13 b | 8.24 ± 1.46 b | |
| Fermented | 0.59 ± 0.05 a | 1.24 ± 0.23 a | 0.89 ± 0.22 a | 1.80 ± 0.17 c | 9.72 ± 0.64 c | |
| Fermented Cooked | 0.47 ± 0.10 ab | 1.43 ± 0.26 a | 0.94 ± 0.11 a | 2.15 ± 0.15 a | 7.71 ± 1.49 ab | |
| WhiteLi1 (Liberty.B) | Raw | 0.33 ± 0.01 a | 0.05 ± 0.01 a | 0.25 ± 0.05 a | 0.83 ± 0.19 a | 1.55 ± 0.22 a |
| Cooked | 0.30 ± 0.03 ab | 0.12 ± 0.05 b | 0.42 ± 0.08 b | 1.32 ± 0.21 b | 2.40 ± 0.23 bc | |
| Fermented | 0.40 ± 0.04 ac | 0.14 ± 0.03 bc | 0.46 ± 0.19 bc | 0.88 ± 0.21 a | 2.79 ± 0.55 bc | |
| Fermented Cooked | 0.34 ± 0.11 a | 0.13 ± 0.03 bd | 0.54 ± 0.07 b | 1.08 ± 0.20 a | 2.70 ± 0.50 c | |
| RedBu2 (Buster.CC) | Raw | 0.58 ± 0.09 ab | 0.78 ± 0.05 ab | 0.69 ± 0.15 a | 1.58 ± 0.18 a | 3.74 ± 0.42 a |
| Cooked | 0.49 ± 0.04 a | 0.94 ± 0.10 a | 0.84 ± 0.11 a | 2.39 ± 0.16 b | 6.82 ± 0.57 b | |
| Fermented | 0.42 ± 0.05 ac | 1.02 ± 0.16 a | 0.73 ± 0.05 a | 1.90 ± 0.19 c | 5.11 ± 0.91 c | |
| Fermented Cooked | 0.55 ± 0.18 a | 1.21 ± 0.46 ac | 0.92 ± 0.14 b | 1.85 ± 0.19 cd | 4.58 ± 0.63 acd | |
| RedBa2 (Bazley.CC) | Raw | 0.56 ± 0.04 a | 1.08 ± 0.28 a | 0.84 ± 0.16 a | 1.98 ± 0.17 a | 5.56 ± 0.40 a |
| Cooked | 0.45 ± 0.03 b | 1.15 ± 0.23 a | 0.87 ± 0.10 ab | 2.47 ± 0.13 b | 7.68 ± 1.22 b | |
| Fermented | 0.43 ± 0.06 a | 1.03 ± 0.23 a | 0.68 ± 0.08 ac | 1.70 ± 0.19 c | 4.55 ± 0.72 a | |
| Fermented Cooked | 0.55 ± 0.12 bc | 1.52 ± 0.37 b | 0.95 ± 0.18 abd | 2.08 ± 0.16 a | 5.53 ± 0.65 a | |
| WhiteLi2 (Liberty.CC) | Raw | 0.43 ± 0.02 a | 0.04 ± 0.01 a | 0.41 ± 0.19 a | 0.78 ± 0.22 a | 1.07 ± 0.35 a |
| Cooked | 0.30 ± 0.02 b | 0.08 ± 0.02 b | 0.47 ± 0.16 a | 0.99 ± 0.21 a | 1.69 ± 0.32 a | |
| Fermented | 0.29 ± 0.06 bc | 0.11 ± 0.02 c | 0.46 ± 0.17 a | 0.78 ± 0.21 a | 2.45 ± 0.77 b | |
| Fermented Cooked | 0.38 ± 0.03 d | 0.21 ± 0.04 d | 0.46 ± 0.21 a | 0.97 ± 0.23 a | 2.89 ± 0.65 b |
| Peak | Tentative ID | Level of ID | m/z | Retention Time | Sorghum Variety | Sorghum Processing Technique | |||
|---|---|---|---|---|---|---|---|---|---|
| Raw | Cooked | Fermented | Fermented Cooked | ||||||
| 1 | trans-Piceid | 2 | 251.0797 | 1.06 | WhiteLi1, RedBa1, RedBu1, RedBu2 | √ | √ | ||
| 2 | trans cinnamic acid | 2 | 147.0310 | 1.07 | BlackSs | √ | |||
| 3 | 6-methoxy-7-hydroxycoumarin | 3 | 193.0360 | 1.09 | RedBu1, BlackSb | √ | √ | ||
| 4 | Methyl trans-cinnamic acid | 4 | 161.0479 | 1.10 | WhiteLi1, RedBa2 | √ | |||
| 5 | Malic acid | 2 | 133.0491 | 1.11 | RedBa2, RedBu2 | √ | √ | ||
| 6 | 6,8-dimethyl-4-hydroxycoumarin | 3 | 191.0900 | 1.32 | RedBu1, RedBu2, BlackSb | √ | √ | ||
| 7 | 6-prenylnaringenin | 4 | 341.1112 | 1.70 | RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | ||
| 8 | Apigenin-7-O-glucoside | 3 | 413.1686 | 2.16 | WhiteLi2 | √ | √ | ||
| 9 | Coumarin | 1 | 145.0510 | 2.47 | RedBa1, RedBa2, RedBu2, BlackSs | √ | √ | √ | |
| 10 | Protocatechuic acid | 1 | 153.0198 | 3.00 | BlackSs | √ | √ | ||
| 11 | Gallic acid | 1 | 170.1200 | 3.14 | WhiteLi1 | √ | |||
| 12 | Vanillin | 1 | 153.0188 | 4.03 | BlackSs | √ | |||
| 13 | Apigenin-7-O-glucoside | 3 | 413.1680 | 4.39 | RedBa1 | √ | √ | ||
| 14 | Quercetin-3,4’-O-di-beta-glucopyranoside | 4 | 625.1985 | 4.50 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 15 | Linoleic acid | 3 | 279.1110 | 4.91 | RedBa1 | √ | |||
| 16 | Catechin | 1 | 289.0726 | 5.14 | BlackSs | √ | |||
| 17 | Malvin | 5 | 655.2112 | 5.47 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, BlackSs | √ | √ | ||
| 18 | 4-methyl-7-aminocoumarin | 3 | 175.0630 | 6.35 | WhiteLi1, WhiteLi2 | √ | √ | ||
| 19 | Hippuric acid | 1 | 179.0388 | 7.30 | WhiteLi1, WhiteLi2, RedBa1, RedBa2 | √ | √ | √ | |
| 20 | 4-Acetylbutyric acid | 2 | 131.0721 | 7.55 | WhiteLi1, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | ||
| 21 | Caffeic acid | 1 | 181.0512 | 7.75 | WhiteLi1, WhiteLi2, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | ||
| 22 | 3-aminobenzoic acid | 2 | 137.0622 | 8.55 | RedBa2, BlackSb | √ | |||
| 23 | Procyanidin b1 isomer | 4 | 577.1386 | 8.97 | BlackSs | √ | √ | ||
| 24 | Malic acid | 2 | 135.0455 | 9.03 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | ||
| 25 | 2-Isopropylmalic acid | 3 | 177.0201 | 9.20 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs | √ | √ | √ | √ |
| 26 | Hippuric acid | 1 | 179.0358 | 9.05 | WhiteLi2, RedBu1, RedBu2, BlackSb | √ | √ | ||
| 27 | Catechin | 1 | 289.0733 | 9.08 | BlackSs | √ | |||
| 28 | Caffeic acid | 1 | 181.0516 | 9.51 | WhiteLi2, BlackSs, BlackSb | √ | √ | ||
| 29 | Kaempferol-3-O-xyloside | 3 | 417.1057 | 9.66 | WhiteLi2, RedBa1, RedBa2, BlackSb | √ | √ | ||
| 30 | Luteolin derivative | 3 | 415.1262 | 10.05 | WhiteLi1, WhiteLi2, RedBa1, RedBu2 | √ | |||
| 31 | Apigeninidin | 2 | 253.0737 | 10.43 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 32 | 2-Isopropylmalic acid | 2 | 177.0192 | 10.91 | WhiteLi2 | √ | |||
| 33 | Procyanidin C1 | 4 | 865.1978 | 10.45 | BlackSs | √ | √ | √ | √ |
| 34 | Epigallocatechin | 2 | 307.1409 | 11.42 | BlackSs | √ | |||
| 35 | Catechin | 1 | 289.0739 | 11.46 | BlackSs | √ | √ | √ | |
| 36 | trans cinnamic acid | 2 | 147.0453 | 11.79 | RedBa1, RedBu2, BlackSs | √ | √ | ||
| 37 | trans-Resveratroloside | 3 | 371.1000 | 12.08 | WhiteLi1, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | ||
| 38 | Isoferulic acid | 2 | 195.0668 | 12.09 | BlackSb | √ | √ | ||
| 39 | 2-Hydroxyhippuric acid | 2 | 253.0729 | 12.11 | WhiteLi2 | √ | |||
| 40 | trans-Resveratroloside | 3 | 371.1001 | 12.16 | WhiteLi2, BlackSs, BlackSb | √ | √ | √ | |
| 41 | Catechin derivative | 3 | 720.1572 | 12.73 | BlackSs | √ | √ | √ | √ |
| 42 | N’.n’-dicafferoylspermidine | 2 | 468.2130 | 13.21 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 43 | Ferulic acid | 1 | 468.2156 | 13.56 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 44 | Luteolin | 1 | 285.0736 | 13.93 | BlackSs | √ | |||
| 45 | Catechin derivative | 3 | 289.0758 | 14.21 | BlackSs, BlackSb | √ | √ | ||
| 46 | Malic acid | 2 | 135.0458 | 14.53 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSb | √ | √ | ||
| 47 | Catechin | 1 | 289.0738 | 14.60 | BlackSs | √ | √ | ||
| 48 | 2-Isopropylmalic acid | 2 | 177.0192 | 14.63 | RedBa2 | √ | |||
| 49 | Apigenin | 1 | 269.0475 | 14.85 | BlackSs, BlackSb | √ | √ | √ | |
| 50 | Taxifolin-O-pentoside | 3 | 399.1681 | 14.89 | RedBa1 | √ | |||
| 51 | Pentahydroxyflavanone-(3 → 4)-catechin-7-O-glucoside | 4 | 883.2293 | 15.00 | RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 52 | Eriodictyol deoxyhexoside | 3 | 433.1181 | 15.12 | RedBa1, RedBa2, RedBu1, RedBu2 | √ | √ | ||
| 53 | Eriodictyol deoxyhexoside | 3 | 433.1169 | 15.45 | RedBu1, RedBu2, BlackSs | √ | √ | √ | |
| 54 | Glucomalcomiin | 2 | 482.2334 | 15.39 | RedBa1, RedBu1, BlackSs, BlackSb | √ | √ | √ | |
| 55 | Procyanidin C1 | 4 | 865.2171 | 15.43 | RedBa2, RedBu2, BlackSs | √ | √ | ||
| 56 | 3-aminobenzoic acid | 2 | 137.0613 | 15.45 | WhiteLi1, RedBu1 | √ | √ | ||
| 57 | Kaempferol | 2 | 187.0980 | 15.67 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 58 | Epicat-(4beta → 6)-epicatechin-(2beta- > 7,4beta → 8)-epicatechin | 2 | 867.2377 | 16.36 | RedBu1 | √ | √ | ||
| 59 | Procyanidin C1 | 4 | 867.2382 | 16.37 | RedBa1, RedBa2, RedBu1, RedBu2, BlackSs | √ | √ | √ | √ |
| 60 | Procyanidin b1 isomer | 4 | 577.1590 | 16.79 | WhiteLi1, WhiteLi2 | √ | |||
| 61 | Luteolin-7-O-glucoside | 2 | 447.0954 | 16.81 | RedBa1, RedBa2, RedBu1, RedBu2 | √ | √ | √ | |
| 62 | Pyrano-eriodictyol-(3 → 4)-catechin-7-O-glucoside | 4 | 867.2362 | 16.99 | RedBa1, RedBa2, RedBu1, BlackSs | √ | √ | √ | √ |
| 63 | Apigenin | 1 | 271.0632 | 17.05 | RedBa1, BlackSs, BlackSb | √ | √ | ||
| 64 | Taxifolin | 2 | 303.0527 | 17.69 | RedBa1, RedBu1, BlackSs, BlackSb | √ | √ | √ | |
| 65 | Malvidin-3-O-glucoside | 2 | 329.2133 | 17.91 | RedBa1 | √ | |||
| 66 | Procyanidin | 2 | 429.2133 | 17.98 | RedBa1, RedBa2, RedBu1, RedBu2, BlackSb | √ | √ | √ | √ |
| 67 | Pyrano-naringenin-(3 → 4)-catechin-7-O-glucoside isomer | 3 | 851.2415 | 18.55 | RedBa1, RedBa2, RedBu1, RedBu2, BlackSs | √ | √ | √ | √ |
| 68 | Taxifolin | 2 | 303.0879 | 19.23 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 69 | Kaempferol | 2 | 187.0984 | 19.32 | WhiteLi2 | √ | |||
| 70 | Luteolin derivative | 3 | 415.1061 | 19.35 | BlackSb | √ | |||
| 71 | Taxifolin-O-pentoside | 2 | 399.1102 | 19.75 | BlackSb | √ | |||
| 72 | Quercetin-3-O-glucuronide | 2 | 478.9223 | 19.84 | WhiteLi1, WhiteLi2 | √ | |||
| 73 | Eriodictyol deoxyhexoside | 3 | 721.1826 | 19.98 | RedBa1 | √ | |||
| 74 | Pentahydroxyflavanone-(3 → 4)-catechin-7-O-glucoside isomer | 4 | 721.1780 | 19.99 | RedBa1, RedBa2, RedBu1, RedBu2 | √ | √ | √ | √ |
| 75 | Eriodictyol deoxyhexoside | 3 | 433.1121 | 20.61 | RedBa2 | √ | |||
| 76 | Luteolin | 1 | 285.0766 | 21.45 | RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 77 | 7-O-methyl-luteolinidin | 5 | 395.2135 | 21.49 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs | √ | √ | √ | √ |
| 78 | Voacamine | 2 | 705.1871 | 21.83 | RedBa1, RedBa2, RedBu2 | √ | √ | √ | |
| 79 | Procyanidin | 2 | 429.1216 | 22.12 | BlackSb | √ | |||
| 80 | Luteolin derivative | 3 | 415.1070 | 22.41 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | √ |
| 81 | Taxifolin-O-pentoside | 3 | 399.1104 | 24.36 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | √ | |
| 82 | Apigenin | 1 | 271.0627 | 24.43 | RedBu1 | √ | |||
| 83 | Procyanidin | 2 | 429.1212 | 24.91 | WhiteLi1, WhiteLi2, RedBa1, RedBa2, RedBu1, RedBu2, BlackSs, BlackSb | √ | √ | ||
| 84 | Apigenin | 1 | 269.0471 | 25.17 | RedBa1 | √ | |||
| 85 | Gallic acid hexoside | 3 | 331.2498 | 25.92 | RedBa2, RedBu1, RedBu2, BlackSs | √ | √ | ||
| 86 | Malvidin-3-O-glucoside | 5 | 329.2539 | 25.93 | RedBa1, RedBu1 | √ | √ | ||
| 87 | Apigenin-7-O-glucoside | 2 | 413.1276 | 27.05 | BlackSb | √ | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collins, A.; Santhakumar, A.; Latif, S.; Chinkwo, K.; Francis, N.; Blanchard, C. Impact of Processing on the Phenolic Content and Antioxidant Activity of Sorghum bicolor L. Moench. Molecules 2024, 29, 3626. https://doi.org/10.3390/molecules29153626
Collins A, Santhakumar A, Latif S, Chinkwo K, Francis N, Blanchard C. Impact of Processing on the Phenolic Content and Antioxidant Activity of Sorghum bicolor L. Moench. Molecules. 2024; 29(15):3626. https://doi.org/10.3390/molecules29153626
Chicago/Turabian StyleCollins, Aduba, Abishek Santhakumar, Sajid Latif, Kenneth Chinkwo, Nidhish Francis, and Christopher Blanchard. 2024. "Impact of Processing on the Phenolic Content and Antioxidant Activity of Sorghum bicolor L. Moench" Molecules 29, no. 15: 3626. https://doi.org/10.3390/molecules29153626
APA StyleCollins, A., Santhakumar, A., Latif, S., Chinkwo, K., Francis, N., & Blanchard, C. (2024). Impact of Processing on the Phenolic Content and Antioxidant Activity of Sorghum bicolor L. Moench. Molecules, 29(15), 3626. https://doi.org/10.3390/molecules29153626