
Ultrasound as Green Technology for the Valorization of Pumpkin Leaves: Intensification of Protein Recovery

 , , , , , , , and
, , , , , , , and 
Abstract
:1. Introduction
2. Results and Discussion
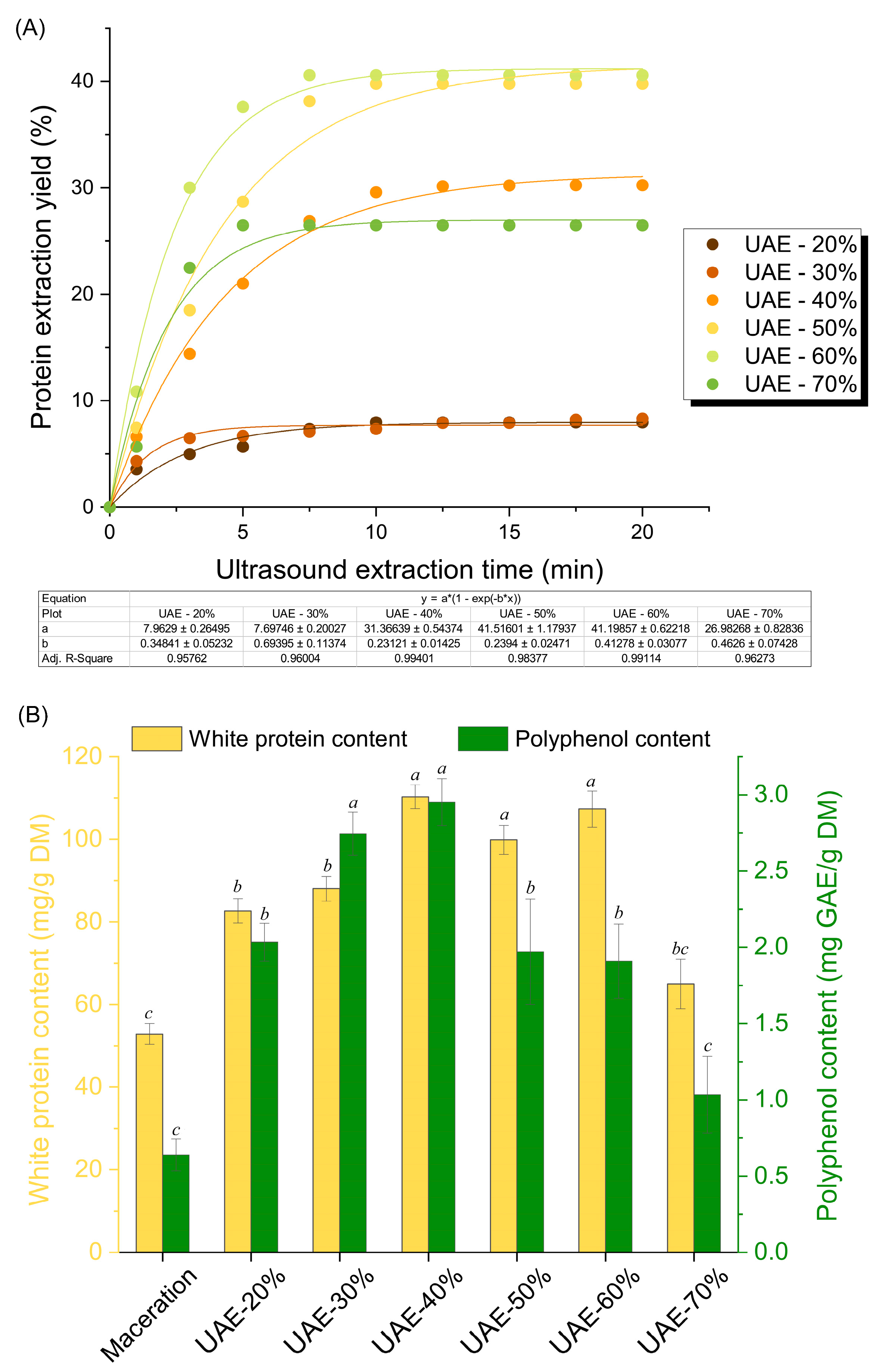
2.1. Impact of Ultrasound-Assisted Extraction on Protein Recovery and Bioactivity of RuBisCO-Rich Protein Fractions
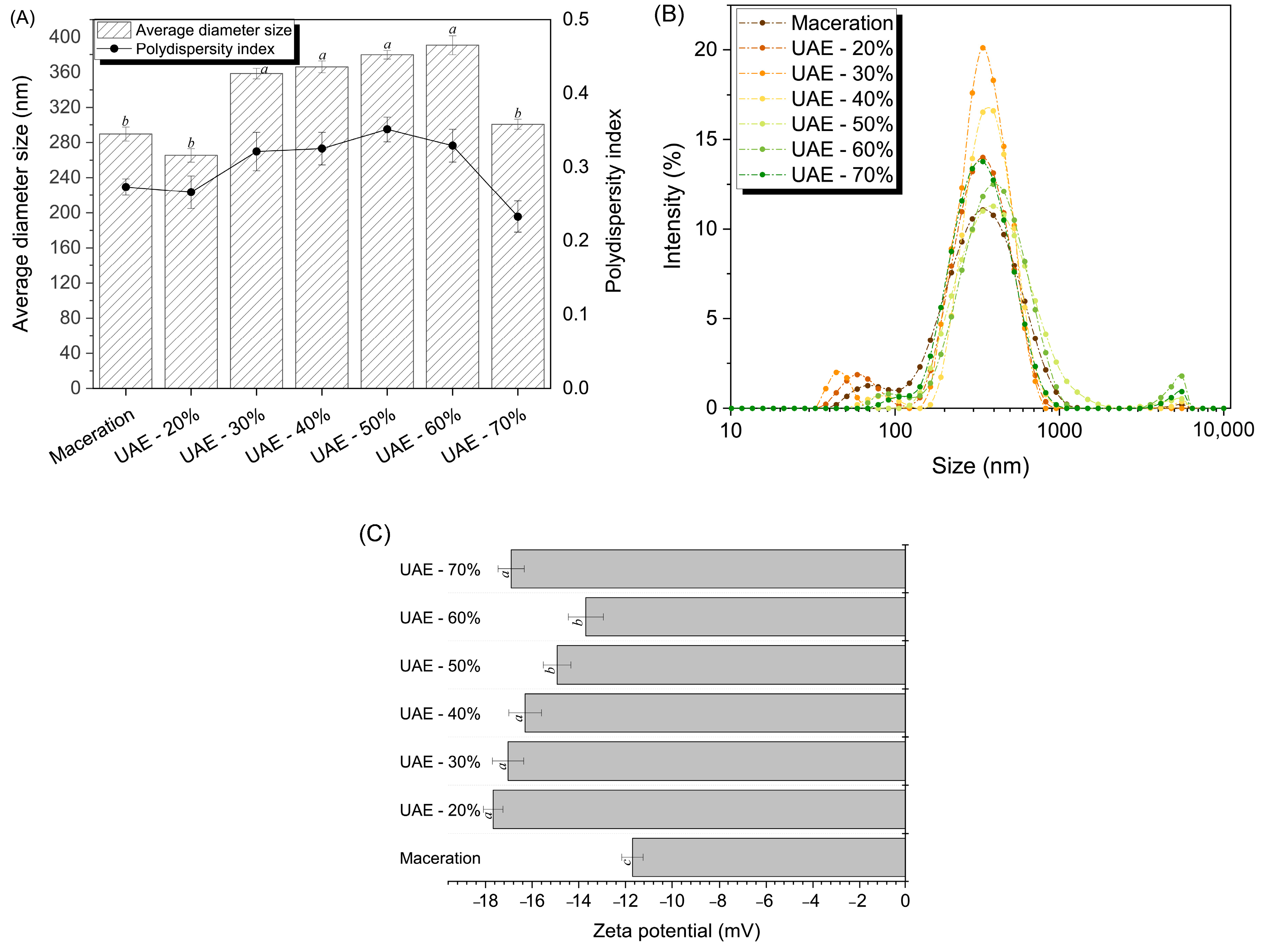
2.2. Structural and Morphological Characterization of RuBisCO-Rich Protein Fractions
2.2.1. Zeta Potential, Particle Size Analysis, and Molecular Weight Distribution
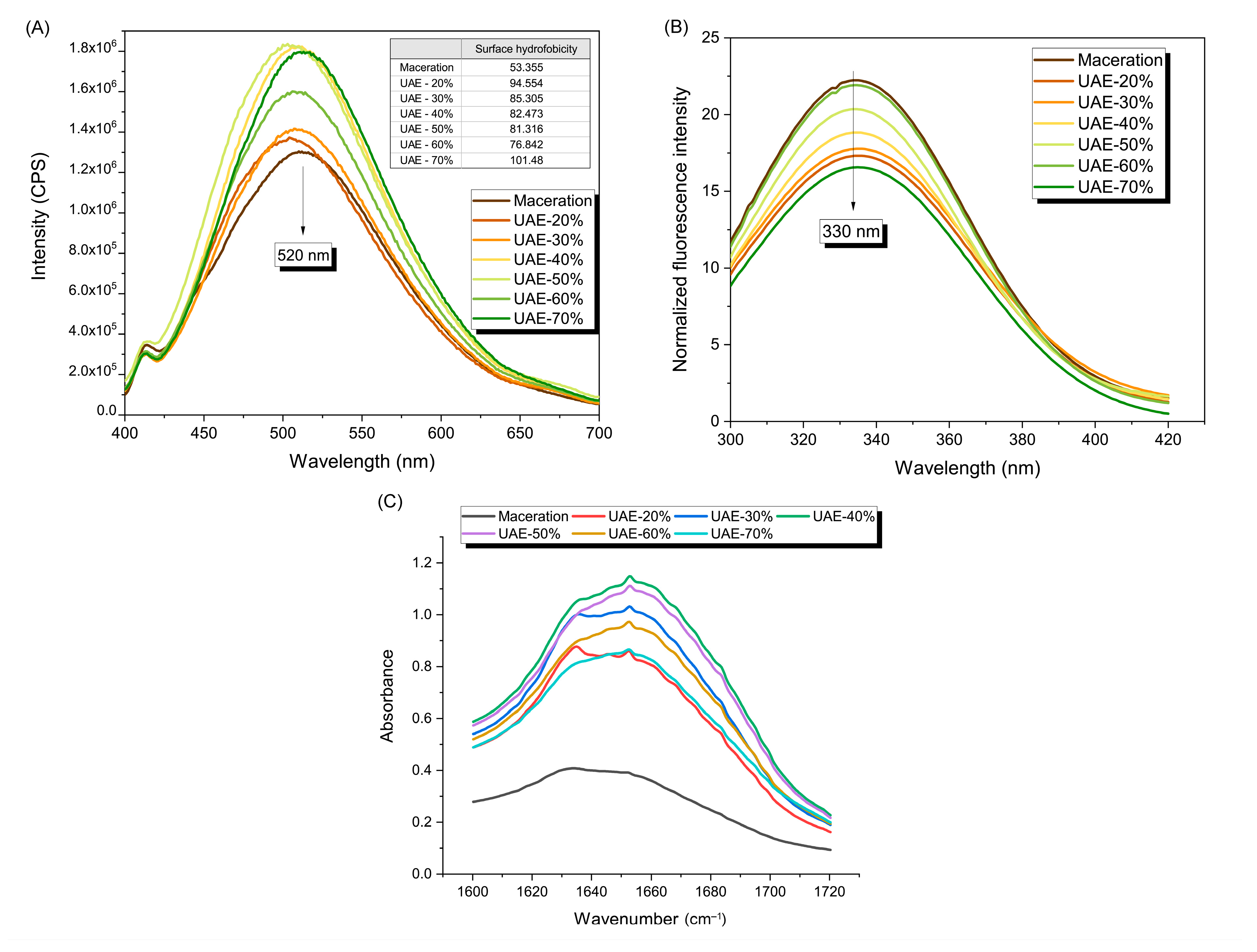
2.2.2. Surface Hydrophobicity, Intrinsic Fluorescence Spectra, and Secondary Structure of the Protein Side Chain
2.2.3. Scanning Electron Microscopy Analysis
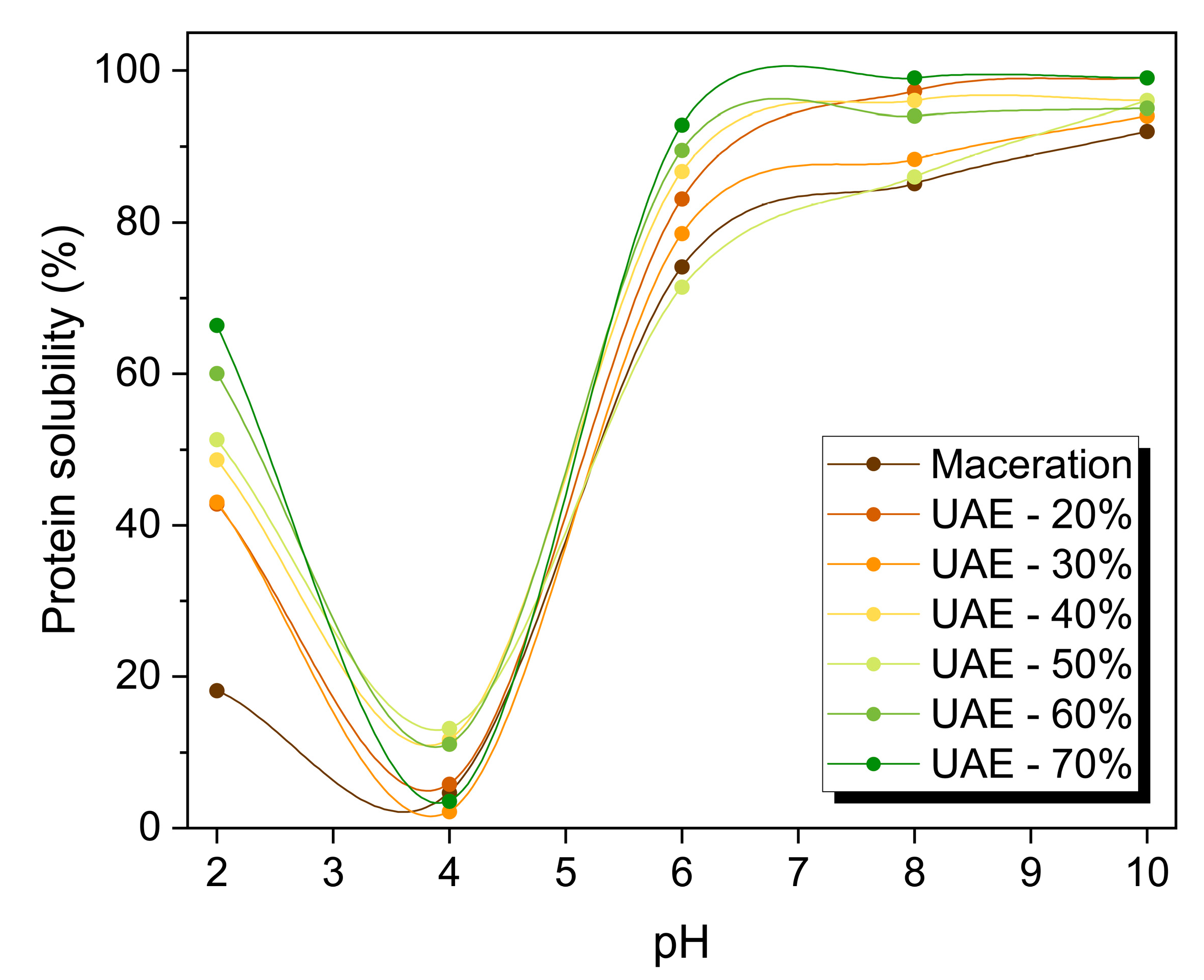
2.3. Functionality of RuBisCO-Rich Protein Fractions
3. Materials and Methods
3.1. Plant Material
3.2. Chemical Reagents
3.3. Ultrasound-Assisted Extraction of Proteins from Pumpkin Leaves
3.3.1. Evaluation of Protein Content and Protein Recovery
3.3.2. Evaluation of Total Polyphenol Content and Polyphenol Recovery
3.4. Molecular Structure and Morphology Characterization of RuBisCO-Rich Protein Fractions
3.4.1. Evaluation of the Presence of RuBisCO Protein Subunits
3.4.2. Evaluation of Intrinsic Fluorescence Spectra and Surface Hydrophobicity
3.4.3. Secondary Protein Structure Analysis by Used FTIR Spectrometry
3.4.4. Evaluation of Average Diameter Size and Surface Electrical Charge
3.4.5. Morphological Characterization by Used FESEM Analysis
3.5. In Vitro Bioactivities of RuBisCO-Rich Protein Fractions
3.5.1. Evaluation of ABTS•+ Radical Scavenging Activity
3.5.2. Evaluation of Ferrous Ions’ Chelating Activity
3.5.3. Evaluation of Hydroxyl Radical Scavenging Activity
3.6. Functional Properties of RuBisCO-Rich Protein Fractions
3.6.1. Evaluation of Protein Solubility
3.6.2. Evaluation of Water-Holding and Oil-Holding Capacities
3.6.3. Evaluation of Emulsification Activity and Stability Index
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heppner, S.; Livney, Y.D. Green leaves as a promising source for sustainable food protein: Seeking the productivity-functionality balance. Trends Food Sci. Technol. 2023, 142, 104207. [Google Scholar] [CrossRef]
- Munialo, C.D. A review of alternative plant protein sources, their extraction, functional characterisation, application, nutritional value, and pinch points to being the solution to sustainable food production. Int. J. Food Sci. Technol. 2024, 59, 462–472. [Google Scholar] [CrossRef]
- Aiking, H.; de Boer, J. The next protein transition. Trends Food Sci. Technol. 2020, 105, 515–522. [Google Scholar] [CrossRef]
- Hadidi, M.; Palacios, J.C.O.; McClements, D.J.; Mahfouzi, M.; Moreno, A. Alfalfa as a sustainable source of plant-based food proteins. Trends Food Sci. Technol. 2023, 135, 202–214. [Google Scholar] [CrossRef]
- Santamaría-Fernández, M.; Lübeck, M. Production of leaf protein concentrates in green biorefineries as alternative feed for monogastric animals. Anim. Feed Sci. Technol. 2020, 268, 114605. [Google Scholar] [CrossRef]
- Tenorio, A.T.; Gieteling, J.; De Jong, G.A.; Boom, R.M.; Van Der Goot, A.J. Recovery of protein from green leaves: Overview of crucial steps for utilisation. Food Chem. 2016, 203, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tenorio, A.T.; Kyriakopoulou, K.E.; Suarez-Garcia, E.; van den Berg, C.; van der Goot, A.J. Understanding differences in protein fractionation from conventional crops, and herbaceous and aquatic biomass-consequences for industrial use. Trends Food Sci. Technol. 2018, 71, 235–245. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Kasote, D.M.; Ullah, N.; Usman, K.; Alsafran, M. RuBisCO: A sustainable protein ingredient for plant-based foods. Front. Sustain. Food Syst. 2024, 8, 1389309. [Google Scholar] [CrossRef]
- Grácio, M.; Oliveira, S.; Lima, A.; Ferreira, R.B. RuBisCO as a protein source for potential food applications: A review. Food Chem. 2023, 419, 135993. [Google Scholar] [CrossRef]
- Dukić, J.; Košpić, K.; Kelava, V.; Mavrić, R.; Nutrizio, M.; Balen, B.; Butorac, A.; Öztop Halil, M.; Jambrak, A.R. Alternative methods for RuBisCO extraction from sugar beet waste: A comparative approach of ultrasound and high voltage electrical discharge. Ultrason. Sonochem. 2023, 99, 106535. [Google Scholar] [CrossRef]
- Hu, Z.; Hu, C.; Li, Y.; Jiang, Q.; Li, Q.; Fang, C. Pumpkin seed oil: A comprehensive review of extraction methods, nutritional constituents, and health benefits. J. Sci. Food Agric. 2024, 104, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, M.; Aghababaei, F.; McClements, D.J. Enhanced alkaline extraction techniques for isolating and modifying plant-based proteins. Food Hydrocoll. 2023, 145, 109132. [Google Scholar] [CrossRef]
- Kumar, M.; Tomar, M.; Punia, S.; Grasso, S.; Arrutia, F.; Choudhary, J.; Singh, S.; Verma, P.; Mahapatra, A.; Patil, S.; et al. Cottonseed: A sustainable contributor to global protein requirements. Trends Food Sci. Technol. 2021, 111, 100–113. [Google Scholar] [CrossRef]
- Rahman, M.M.; Lamsal, B.P. Ultrasound-assisted extraction and modification of plant-based proteins: Impact on physicochemical, functional, and nutritional properties. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1457–1480. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Yao, H.; Zhou, J.; Duan, Y.; Zhang, H.; Ma, H. Advances in renewable plant-derived protein source: The structure, physicochemical properties affected by ultrasonication. Ultrason. Sonochem. 2019, 53, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, G.; Yildiz, S.; Karaca, A.C.; Yemiş, O. Ultrasound and enzyme-pretreated extraction for the valorization of pea pod proteins. J. Food Process Eng. 2023, 46, e14452. [Google Scholar] [CrossRef]
- Ge, S.; Tong, X.; Gao, C.; Xu, H.; He, R.; Wu, Q.; Wang, J. Kinetics of silkworm pupae protein extraction at different ultrasonic frequency and temperature: Effects on physicochemical properties, functional properties and oxidation resistance. Process Biochem. 2022, 122, 36–52. [Google Scholar] [CrossRef]
- Flores-Jiménez, N.T.; Ulloa, J.A.; Urías-Silvas, J.E.; Hidalgo-Millán, A. Modification of rheological properties of animal and vegetable proteins treated with high-intensity ultrasound: A review. Food Front. 2023, 4, 700–720. [Google Scholar] [CrossRef]
- Das, R.S.; Tiwari, B.K.; Chemat, F.; Garcia-Vaquero, M. Impact of ultrasound processing on alternative protein systems: Protein extraction, nutritional effects and associated challenges. Ultrason. Sonochemistry 2022, 91, 106234. [Google Scholar] [CrossRef]
- Chemat, F.; Rombaut, N.; Sicaire, A.G.; Meullemiestre, A.; Fabiano-Tixier, A.S.; Abert-Vian, M. Ultrasound assisted extraction of food and natural products. Mechanisms, techniques, combinations, protocols and applications. A review. Ultrason. Sonochem. 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Milićević, N.; Kojić, P.; Sakač, M.; Mišan, A.; Kojić, J.; Perussello, C.; Banjac, V.; Pojić, M.; Tiwari, B. Kinetic modelling of ultrasound-assisted extraction of phenolics from cereal brans. Ultrason. Sonochem. 2021, 79, 105761. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.U.; Jawaid, P.; Uchiyama, H.; Kondo, T. Comparison of free radicals formation induced by cold atmospheric plasma, ultrasound, and ionizing radiation. Arch. Biochem. Biophys. 2016, 605, 19–25. [Google Scholar] [CrossRef]
- Hadidi, M.; Khaksar, F.B.; Pagan, J.; Ibarz, A. Application of Ultrasound-Ultrafiltration-Assisted alkaline isoelectric precipitation (UUAAIP) technique for producing alfalfa protein isolate for human consumption: Optimization, comparison, physicochemical, and functional properties. Food Res. Int. 2020, 130, 108907. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cheng, X.; Song, X.; Ouyang, D.; Wang, J.; Wu, Q.; Jia, J. Ultrasonic assisted extraction of mulberry leaf protein: Kinetic model, structural and functional properties, in vitro digestion. Process Biochem. 2023, 128, 12–21. [Google Scholar] [CrossRef]
- Deng, Y.; Yang, X.; Yan, T.; Xu, W.; Li, J.; Niu, R.; Zhao, R.; Wang, H.; Wang, H.; Chen, T.; et al. Ultrasound-induced cell disintegration and its ultrastructure characterization for the valorisation of Chlorella pyrenoidosa protein. Bioresour. Technol. 2023, 381, 129046. [Google Scholar] [CrossRef] [PubMed]
- Dabbour, M.; Jiang, H.; Mintah, B.K.; Wahia, H.; He, R. Ultrasonic-assisted protein extraction from sunflower meal: Kinetic modeling, functional, and structural traits. Innov. Food Sci. Emerg. Technol. 2021, 74, 102824. [Google Scholar] [CrossRef]
- Perović, M.N.; Jugović, Z.D.K.; Antov, M.G. Heat-induced nanoparticles from pumpkin leaf protein for potential application as β-carotene carriers. Future Foods 2024, 9, 100310. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, Y.; Xu, L.; Ma, H. An efficient ultrasound-assisted extraction method of pea protein and its effect on protein functional properties and biological activities. Lwt—Food Sci. Technol. 2020, 127, 109348. [Google Scholar] [CrossRef]
- Haniewicz, P.; De Sanctis, D.; Büchel, C.; Schröder, W.P.; Loi, M.C.; Kieselbach, T.; Bochtler, M.; Piano, D. Isolation of monomeric photosystem II that retains the subunit PsbS. Photosynth. Res. 2013, 118, 199–207. [Google Scholar] [CrossRef]
- Huang, Z.; Shen, L.; Wang, W.; Mao, Z.; Yi, X.; Kuang, T.; Shen, J.-R.; Zhang, X.; Han, G. Structure of photosystem I-LHCI-LHCII from the green alga Chlamydomonas reinhardtii in State 2. Nat Commun. 2021, 12, 1100. [Google Scholar] [CrossRef]
- Pagliano, C.; Nield, J.; Marsano, F.; Pape, T.; Barera, S.; Saracco, G.; Barber, J. Proteomic characterization and three-dimensional electron microscopy study of PSII–LHCII supercomplexes from higher plants. Biochim. Biophys. Acta (BBA)-Bioenerg. 2014, 1837, 1454–1462. [Google Scholar] [CrossRef]
- Eftink, M.R. Fluorescence techniques for studying protein structure. Methods Biochem. Anal. 1991, 35, 127–205. [Google Scholar] [CrossRef]
- Yeasmen, N.; Orsat, V. Pulsed ultrasound assisted extraction of alternative plant protein from sugar maple leaves: Characterization of physical, structural, thermal, electrical, and techno-functional properties. Food Hydrocoll. 2024, 152, 109960. [Google Scholar] [CrossRef]
- Vera-Salgado, J.; Calderón-Chiu, C.; Calderón-Santoyo, M.; Barros-Castillo, J.C.; López-García, U.M.; Ragazzo-Sánchez, J.A. Ultrasound-assisted extraction of Artocarpus heterophyllus L. leaf protein concentrate: Solubility, foaming, emulsifying, and antioxidant properties of protein hydrolysates. Colloids Interfaces. 2022, 6, 50. [Google Scholar] [CrossRef]
- Sadat, A.; Joye, I.J. Peak fitting applied to Fourier transform infrared and Raman spectroscopic analysis of proteins. Appl. Sci. 2020, 10, 5918. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 20th ed.; Latimer, G.W., Jr., Ed.; AOAC International: Rockville, MD, USA, 2016. [Google Scholar]
- Hartree, E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Jakovetić Tanasković, S.; Šekuljica, N.; Jovanović, J.; Gazikalović, I.; Grbavčić, S.; Đorđević, N.; Vukašinović Sekulić, M.; Hao, J.; Luković, N.; Knežević-Jugović, Z. Upgrading of valuable food component contents and anti-nutritional factors depletion by solid-state fermentation: A way to valorize wheat bran for nutrition. J. Cereal Sci. 2021, 99, 103159. [Google Scholar] [CrossRef]
- Laemmli, U. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Kato, A.; Nakai, S. Hydrophobicity determined by a fluorescence probe method and its correlation with surface properties of proteins. Biochim. Biophys. Acta 1980, 624, 13–20. [Google Scholar] [CrossRef]
- Li, C.; Huang, X.; Peng, Q.; Shan, Y.; Xue, F. Physicochemical properties of peanut protein isolate-glucomannan conjugates prepared by ultrasonic treatment. Ultrason. Sonochem. 2014, 21, 1722–1727. [Google Scholar] [CrossRef]
- Knežević-Jugović, Z.; Culetu, A.; Mijalković, J.; Duta, D.; Stefanović, A.; Šekuljica, N.; Đorđević, V.; Antov, M. Impact of Different Enzymatic Processes on Antioxidant, Nutritional and Functional Properties of Soy Protein Hydrolysates Incorporated into Novel Cookies. Foods 2023, 12, 24. [Google Scholar] [CrossRef]
- Chung, S.-K.; Osawa, T.; Kawakishi, S. Hydroxyl Radical-scavenging Effects of Spices and Scavengers from Brown Mustard (Brassica nigra). Biosci. Biotechnol. Biochem. 1997, 61, 118–123. [Google Scholar] [CrossRef]
- Jovanović, J.; Stefanović, A.; Culetu, A.; Duta, D.; Luković, N.; Tanasković, S.J.; Šekuljica, N.; Knežević-Jugović, Z. Enzymatic treatment of soy protein concentrate: Influence on the potential techno-functional and antioxidant properties. J. Hyg. Eng. Des. (JHED) 2020, 30, 58–68. Available online: https://keypublishing.org/jhed/wp-content/uploads/2020/11/05.-Full-paper-Jelena-Jovanovic.pdf (accessed on 10 July 2024).
- Moure, A.; Domínguez, H.; Zúñiga, M.A.E.; Soto, C.; Chamy, R. Characterisation of protein concentrates from pressed cakes of Guevina avellana (Chilean hazelnut). Food Chem. 2002, 78, 179–186. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | ABTS Radical Scavenging Activity (mmol TE/g) | Metal-Ion Chelating Activity (mmol EE/g) | Hydroxyl Radical Scavenging Activity (mmol TE/g) | Protein Content (%) |
|---|---|---|---|---|
| Maceration | 0.871 ± 0.058 ab | 0.380 ± 0.022 c | 1.380 ± 0.023 ac | 52.07 ± 1.25 b |
| UAE-20% | 0.893 ± 0.058 a | 0.364 ± 0.026 c | 1.540 ± 0.069 bc | 54.52 ± 2.78 b |
| UAE-30% | 0.794 ± 0.032 abc | 0.545 ± 0.014 a | 1.716 ± 0.025 b | 62.47 ± 2.07 a |
| UAE-40% | 0.657 ± 0.035 c | 0.462 ± 0.014 b | 2.126 ± 0.016 a | 56.92 ± 0.34 b |
| UAE-50% | 0.521 ± 0.032 d | 0.248 ± 0.013 d | 2.072 ± 0.032 a | 63.95 ± 0.39 a |
| UAE-60% | 0.443 ±0.029 e | 0.229 ± 0.013 d | 1.371 ± 0.015 c | 63.06 ± 0.05 a |
| UAE-70% | 0.436 ±0.025 e | 0.225 ± 0.013 d | 1.334 ± 0.015 a | 62.51 ± 2.83 a |
| Secondary Structure | Band Assignment in the RuBisCO-Rich Protein Fraction (%) | ||||||
|---|---|---|---|---|---|---|---|
| Without Ultrasound * | UAE-20% | UAE-30% | UAE-40% | UAE-50% | UAE-60% | UAE-70% | |
| β-sheet (intermolecular) | 34.55 ± 1.09 a | 19.58 ± 1.12 b | 20.02 ± 1.00 b | 15.08 ± 2.04 c | 16.51 ± 1.55 c | 16.33 ± 1.25 c | 18.22 ± 1.30 b |
| β-sheet (extended) | 15.09 ± 1.06 c | 23.57 ± 1.15 a | 18.95 ± 0.95 b | 17.61 ± 1.24 b | 20.14 ± 1.80 ba | 22.19 ± 1.20 a | 23.28 ± 1.45 a |
| Random coil | n.d. | 13.60 ± 0.85 b | 10.48 ± 1.45 b | 20.39 ± 2.54 a | 11.16 ± 2.02 b | 5.21 ± 1.54 c | n.d. |
| α-helix | 36.71 ± 1.14 b | 12.29 ± 1.65 e | 18.59 ± 1.42 d | 20.44 ± 1.72 c | 25.32 ± 2.00 c | 37.67 ± 1.40 b | 57.24 ± 1.20 a |
| β-turn | 13.64 ± 1.24 d | 30.97 ± 1.42 a | 31.98 ± 2.01 a | 26.49 ± 2.00 b | 26.87 ± 1.72 b | 18.61 ± 2.64 c | 1.26 ± 0.25 e |
| Samples | WHC (g/gproteins) | OHC (g/gproteins) | EAI (m2/gproteins) | ESI (h) |
|---|---|---|---|---|
| Maceration | 4.05 ± 0.58 ab | 6.58 ± 0.51 a | 8.77 ± 0.25 a | 3.56 ± 0.058 ab |
| UAE-20% | 4.71 ± 0.78 a | 5.15 ± 0.42 a | 6.62 ± 0.44 b | 3.97 ± 0.065 a |
| UAE-30% | 3.72 ± 0.62 b | 3.94 ± 0.52 b | 5.54 ± 0.24 c | 3.34 ± 0.054 b |
| UAE-40% | 2.43 ± 0.45 c | 2.91 ± 1.07 b | 5.21 ± 0.45 c | 1.66 ± 0.073 c |
| UAE-50% | 2.98 ± 0.62 c | 3.15 ± 0.38 b | 5.51 ± 0.31 c | 1.11 ± 0.085 d |
| UAE-60% | 2.48 ± 0.39 c | 2.58 ± 0.34 c | 5.13 ± 0.22 c | 0.28 ± 0.076 e |
| UAE-70% | 2.20 ± 0.35 c | 2.18 ± 0.33 c | n.d. | n.d. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijalković, J.; Šekuljica, N.; Jakovetić Tanasković, S.; Petrović, P.; Balanč, B.; Korićanac, M.; Conić, A.; Bakrač, J.; Đorđević, V.; Bugarski, B.; et al. Ultrasound as Green Technology for the Valorization of Pumpkin Leaves: Intensification of Protein Recovery. Molecules 2024, 29, 4027. https://doi.org/10.3390/molecules29174027
Mijalković J, Šekuljica N, Jakovetić Tanasković S, Petrović P, Balanč B, Korićanac M, Conić A, Bakrač J, Đorđević V, Bugarski B, et al. Ultrasound as Green Technology for the Valorization of Pumpkin Leaves: Intensification of Protein Recovery. Molecules. 2024; 29(17):4027. https://doi.org/10.3390/molecules29174027
Chicago/Turabian StyleMijalković, Jelena, Nataša Šekuljica, Sonja Jakovetić Tanasković, Predrag Petrović, Bojana Balanč, Marija Korićanac, Ana Conić, Jelena Bakrač, Verica Đorđević, Branko Bugarski, and et al. 2024. "Ultrasound as Green Technology for the Valorization of Pumpkin Leaves: Intensification of Protein Recovery" Molecules 29, no. 17: 4027. https://doi.org/10.3390/molecules29174027