
The Effect of Chronic Treatment with the Inhibitor of Phosphodiesterase 5 (PDE5), Sildenafil, in Combination with L-DOPA on Asymmetric Behavior and Monoamine Catabolism in the Striatum and Substantia Nigra of Unilaterally 6-OHDA-Lesioned Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effect of Sildenafil and L-DOPA on Asymmetric Behavior of Unilaterally 6-OHDA-Lesioned Rats
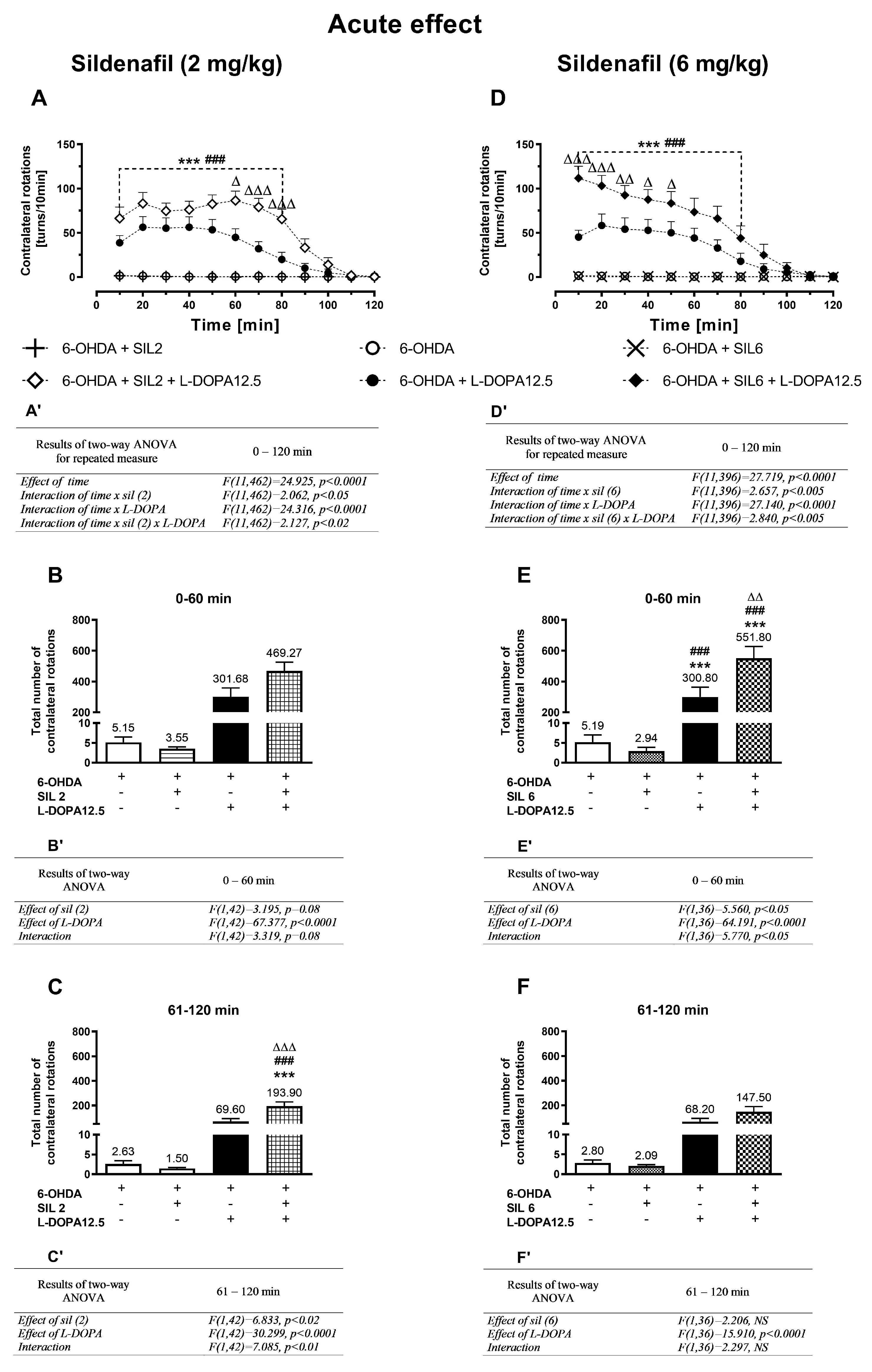
2.1.1. Acute Treatment with Sildenafil (2 mg/kg) and L-DOPA (12.5 mg/kg)
2.1.2. Acute Treatment with Sildenafil (6 mg/kg) and L-DOPA (12.5 mg/kg)
2.1.3. Chronic Treatment with Sildenafil (2 mg/kg) and L-DOPA (12.5 mg/kg)
2.1.4. Chronic Treatment with Sildenafil (6 mg/kg) and L-DOPA (12.5 mg/kg)
2.2. The Effect of Chronic Administration of Sildenafil and L-DOPA on DA Metabolism in the Striatum and Substantia Nigra of Unilaterally 6-OHDA-Lesioned Rats
2.2.1. Changes in Dopamine and Its Metabolite Concentrations in the Striatum
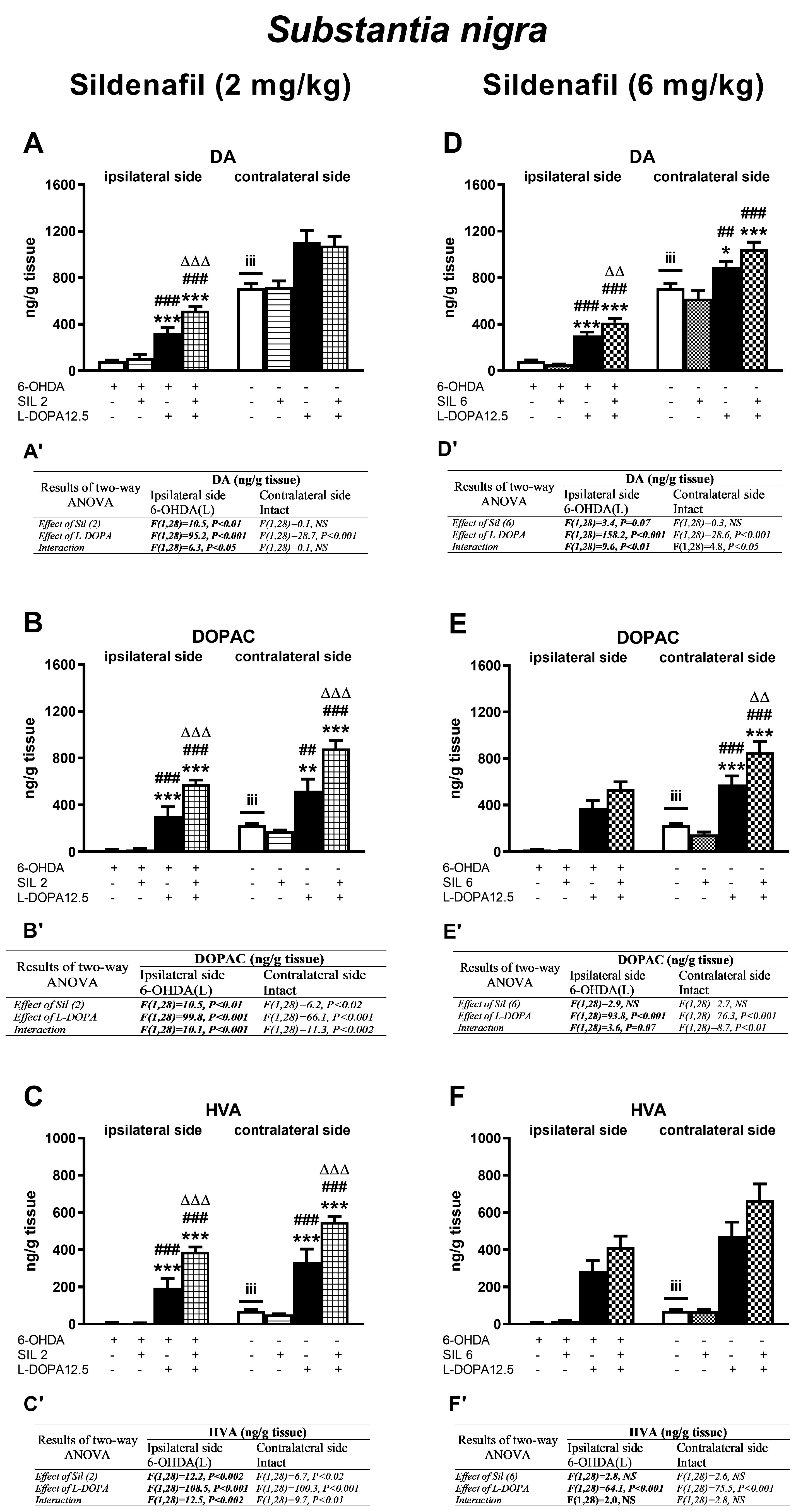
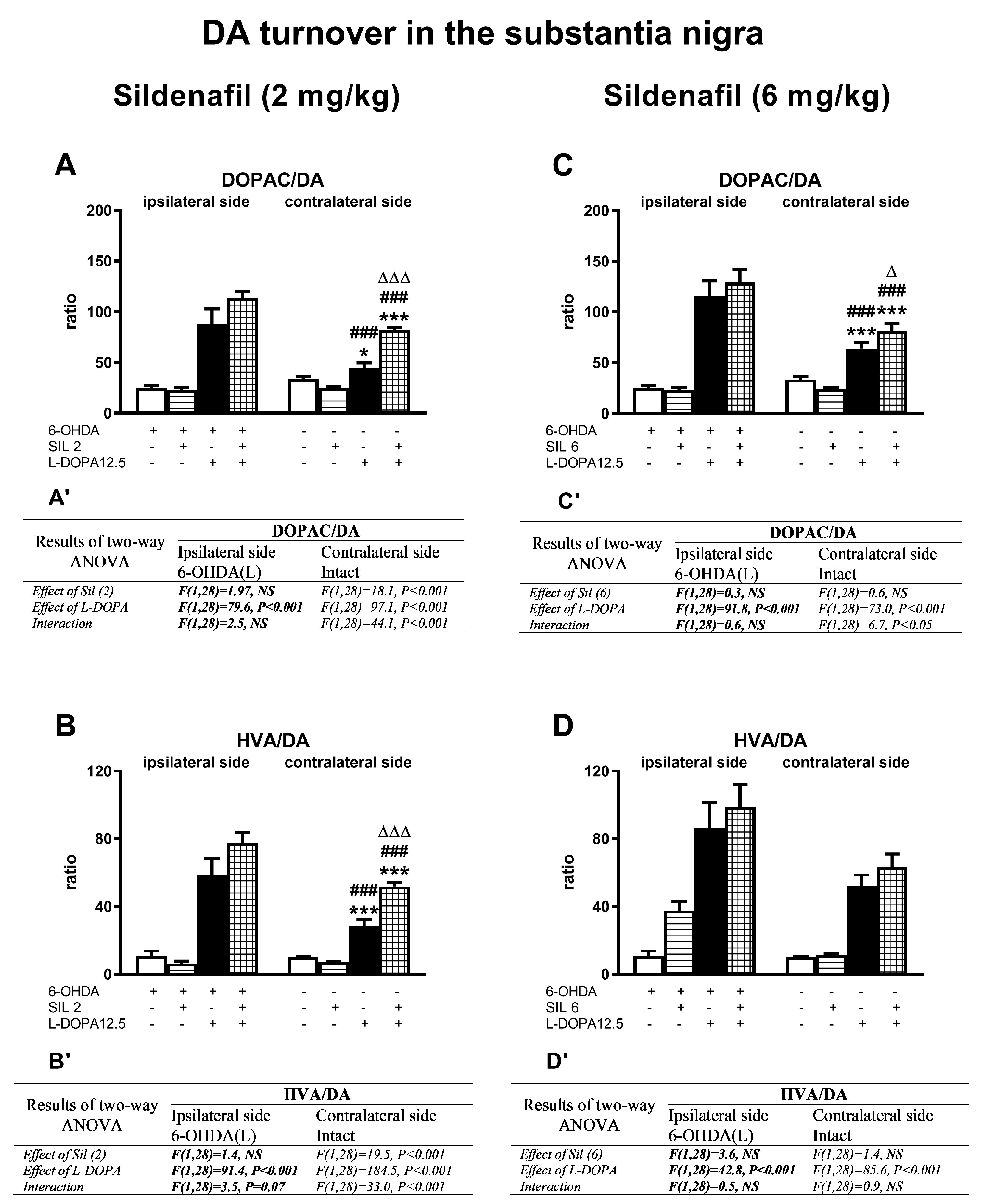
2.2.2. Changes in Dopamine and Its Metabolite Concentrations in the Substantia Nigra
2.2.3. Changes in Serotonin and Its Metabolite Concentrations in the Striatum
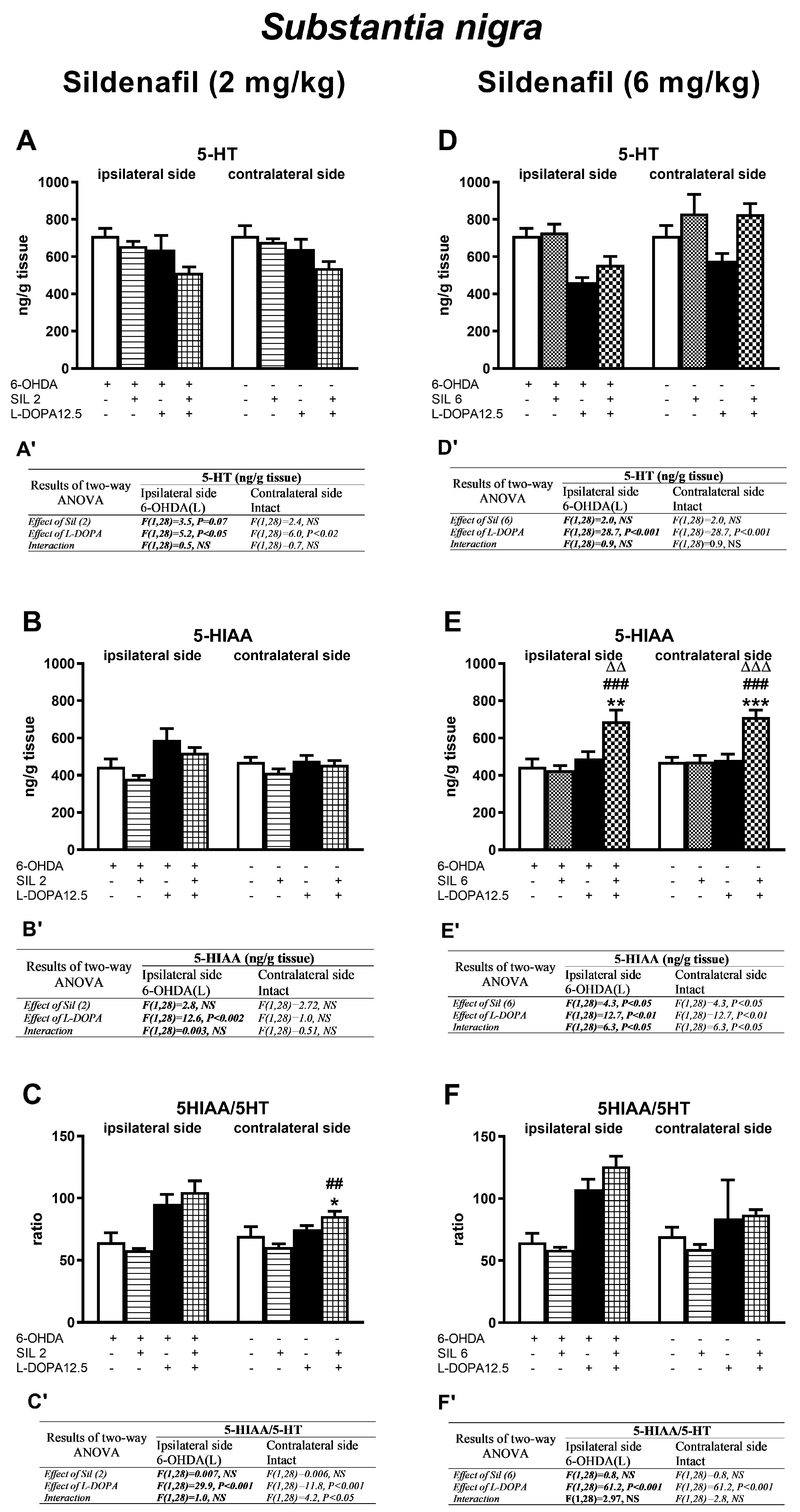
2.2.4. Changes in Serotonin and Its Metabolite Concentrations in the Substantia Nigra
3. Discussion
Limitation of the Study
4. Materials and Methods
4.1. Drugs
4.2. Animals, Surgery, and Drug Treatment
4.3. Asymmetric Behavior
4.4. The Tissue Sample Collection
4.5. Determination of the Concentrations of DA, 5-HT, and Their Metabolites in the Tissue Homogenates
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Bernheimer, H.; Birkmayer, W.; Hornykiewicz, O.; Jellinger, K.; Seitelberger, F. Brain dopamine and the syndromes of Parkinson and Huntington. Clinical, morphological and neurochemical correlations. J. Neurol. Sci. 1973, 20, 415–455. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Bisaglia, M. Oxidative stress and neuroinflammation in Parkinson’s disease: The role of dopamine oxidation products. Antioxidants 2023, 12, 955. [Google Scholar] [CrossRef]
- Ryan, B.J.; Hoek, S.; Fon, E.A.; Wade-Martins, R. Mitochondrial dysfunction and mitophagy in Parkinson’s: From familial to sporadic disease. Trends Biochem. Sci. 2015, 40, 200–210. [Google Scholar] [CrossRef]
- Castelli, V.; Benedetti, E.; Antonosante, A.; Catanesi, M.; Pitari, G.; Ippoliti, R.; Cimini, A.; D’angelo, M. Neuronal cells rearrangement during aging and neurodegenerative disease: Metabolism, oxidative stress and organelles dynamic. Front. Mol. Neurosci. 2019, 12, 132. [Google Scholar] [CrossRef]
- Li, H.; Yang, P.; Knight, W.; Guo, Y.; Perlmutter, J.S.; Benzinger, T.L.S.; Morris, J.C.; Xu, J. The interactions of dopamine and oxidative damage in the striatum of patients with neurodegenerative diseases. J. Neurochem. 2020, 152, 235–251. [Google Scholar] [CrossRef]
- Bej, E.; Cesare, P.; Volpe, A.R.; d’Angelo, M.; Castelli, V. Oxidative stress and neurodegeneration: Insights and therapeutic strategies for Parkinson’s Disease. Neurol. Int. 2024, 16, 502–517. [Google Scholar] [CrossRef]
- Rezak, M. Current pharmacotherapeutic treatment options in Parkinson’s disease. Dis. Mon. 2007, 53, 214–222. [Google Scholar] [CrossRef]
- Lindgren, H.S.; Rylander, D.; Ohlin, K.E.; Lundblad, M.; Cenci, M.A. The “motor complication syndrome” in rats with 6-OHDA lesions treated chronically with L-DOPA: Relation to dose and route of administration. Behav. Brain Res. 2007, 177, 150–159. [Google Scholar] [CrossRef]
- Armstrong, M.J.; Okun, M.S. Diagnosis and treatment of Parkinson disease: A review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef]
- García, A.M.; Redondo, M.; Martinez, A.; Gil, C. Phosphodiesterase 10 inhibitors: New disease modifying drugs for Parkinson’s disease? Curr. Med. Chem. 2014, 21, 1171–1187. [Google Scholar] [CrossRef] [PubMed]
- Beck, G.; Maehara, S.; Chang, P.L.; Papa, S.M. A selective phosphodiesterase 10A inhibitor reduces L-Dopa-induced dyskinesias in parkinsonian monkeys. Mov. Disord. 2018, 33, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Erro, R.; Mencacci, N.E.; Bhatia, K.P. The emerging role of phosphodiesterases in movement disorders. Mov Disord. 2021, 36, 2225–2243. [Google Scholar] [CrossRef]
- Kochoian, H.A.; Bure, C.; Papa, S.M. Targeting striatal glutamate and phosphodiesterases to control L-DOPA-induced dyskinesia. Cells 2023, 12, 2754. [Google Scholar] [CrossRef]
- Nongthombam, P.D.; Haobam, R. Targeting phosphodiesterase 4 as a potential therapy for Parkinson’s disease: A review. Mol. Biol. Rep. 2024, 51, 510. [Google Scholar] [CrossRef] [PubMed]
- Gerfen, C.R.; Surmeier, D.J. Modulation of striatal projection systems by dopamine. Annu. Rev. Neurosci. 2011, 33, 441–466. [Google Scholar] [CrossRef]
- Klaus, A.; Alves da Silva, J.; Costa, R.M. What, if, and when to move: Basal ganglia circuits and self-paced action initiation. Annu. Rev. Neurosci. 2019, 42, 459–483. [Google Scholar] [CrossRef]
- Villalba, R.M.; Smith, Y. Loss and remodeling of striatal dendritic spines in Parkinson’s disease: From homeostasis to maladaptive plasticity? J. Neural. Transm. (Vienna) 2018, 125, 431–447. [Google Scholar] [CrossRef]
- Zhuang, X.; Belluscio, L.; Hen, R. G(olf)alpha mediates dopamine D1 receptor signaling. J. Neurosci. 2000, 20, RC91. [Google Scholar] [CrossRef]
- Undieh, A.S. Pharmacology of signaling induced by dopamine D(1)-like receptor activation. Pharmacol. Ther. 2010, 128, 37–60. [Google Scholar] [CrossRef]
- Sammut, S.; Dec, A.; Mitchell, D.; Linardakis, J.; Ortiguela, M.; West, A.R. Phasic dopaminergic transmission increases NO efflux in the rat dorsal striatum via a neuronal NOS and a dopamine D(1/5) receptor-dependent mechanism. Neuropsychopharmacology 2006, 31, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Sammut, S.; Bray, K.E.; West, A.R. Dopamine D2 receptor-dependent modulation of striatal NO synthase activity. Psychopharmacology 2007, 191, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Park, D.J.; West, A.R. Regulation of striatal nitric oxide synthesis by local dopamine and glutamate interactions. J. Neurochem. 2009, 111, 1457–1465. [Google Scholar] [CrossRef]
- Hoque, K.E.; Indorkar, R.P.; Sammut, S.; West, A.R. Impact of dopamine glutamate interactions on striatal neuronal nitric oxide synthase activity. Psychopharmacology 2010, 207, 571–581. [Google Scholar] [CrossRef]
- Ariano, M.A.; Lewicki, J.A.; Brandwein, H.J.; Murad, F. Immunohistochemical localization of guanylate cyclase within neurons of rat brain. Proc. Natl. Acad. Sci. USA 1982, 79, 1316–1320. [Google Scholar] [CrossRef]
- Ding, J.D.; Burette, A.; Nedvetsky, P.I.; Schmidt, H.H.; Weinberg, R.J. Distribution of soluble guanylyl cyclase in the rat brain. J. Comp. Neurol. 2004, 472, 437–448. [Google Scholar] [CrossRef]
- Sancesario, G.; Morello, M.; Reiner, A.; Giacomini, P.; Massa, R.; Schoen, S.; Bernardi, G. Nitrergic neurons make synapses on dual-input dendritic spines of neurons in the cerebral cortex and the striatum of the rat: Implication for a postsynaptic action of nitric oxide. Neuroscience 2000, 99, 627–642. [Google Scholar] [CrossRef]
- Kelly, M.P. Cyclic nucleotide signaling changes associated with normal aging and age-related diseases of the brain. Cell Signal. 2018, 42, 281–291. [Google Scholar] [CrossRef]
- Neves-Zaph, S.R. Phosphodiesterase diversity and signal processing within cAMP signaling networks. Adv. Neurobiol. 2017, 17, 3–14. [Google Scholar]
- Sancesario, G.; Giorgi, M.; D’Angelo, V.; Medica, A.; Martorana, A.; Morello, M.; Bengtson, C.P.; Bernardi, G. Down-regulation of nitrergic transmission in the rat striatum after nigrostriatal deafferentiation. Eur. J. Neurosci. 2004, 20, 989–1000. [Google Scholar] [CrossRef]
- Sancesario, G.; Morrone, L.A.; D’Angelo, V.; Castelli, V.; Ferrazzoli, D.; Sica, F.; Martorana, A.; Sorge, R.; Cavaliere, F.; Bernardi, G.; et al. Levodopa-induced dyskinesias are associated with transient down-regulation of cAMP and cGMP in the caudate-putamen of hemiparkinsonian rats: Reduced synthesis or increased catabolism? Neurochem. Int. 2014, 79, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Weiner, N. Dopaminergic functional supersensitivity: Effects of chronic L-dopa and carbidopa treatment in an animal model of Parkinson’s disease. J. Pharmacol. Exp. Ther. 1993, 267, 1105–1111. [Google Scholar] [PubMed]
- Giorgi, M.; D’Angelo, V.; Esposito, Z.; Nuccetelli, V.; Sorge, R.; Martorana, A.; Stefani, A.; Bernardi, G.; Sancesario, G. Lowered cAMP and cGMP signalling in the brain during levodopa-induced dyskinesias in hemiparkinsonian rats: New aspects in the pathogenetic mechanisms. Eur. J. Neurosci. 2008, 28, 941–950. [Google Scholar] [CrossRef]
- Giorgi, M.; Melchiorri, G.; Nuccetelli, V.; D’Angelo, V.; Martorana, A.; Sorge, R.; Castelli, V.; Bernardi, G.; Sancesario, G. PDE10A and PDE10A-dependent cAMP catabolism are dysregulated oppositely in striatum and nucleus accumbens after lesion of midbrain dopamine neurons in rat: A key step in parkinsonism physiopathology. Neurobiol. Dis. 2011, 43, 293–303. [Google Scholar] [CrossRef]
- Nishi, A.; Snyder, G.L. Advanced research on dopamine signaling to develop drugs for the treatment of mental disorders: Biochemical and behavioral profiles of phosphodiesterase inhibition in dopaminergic neurotransmission. J. Pharmacol. Sci. 2010, 114, 6–16. [Google Scholar] [CrossRef]
- Ramirez, A.D.; Smith, S.M. Regulation of dopamine signaling in the striatum by phosphodiesterase inhibitors: Novel therapeutics to treat neurological and psychiatric disorders. Cent. Nerv. Syst. Agents Med. Chem. 2014, 14, 72–82. [Google Scholar] [CrossRef]
- Huang, J.X.; Zhu, B.L.; Xu, J.P.; Zhou, Z.Z. Advances in the development of phosphodiesterase 7 inhibitors. Eur. J. Med. Chem. 2023, 250, 115194. [Google Scholar] [CrossRef]
- Mostafa, T. Oral phosphodiesterase type 5 inhibitors: Nonerectogenic beneficial uses. J. Sex Med. 2008, 5, 2502–2518. [Google Scholar] [CrossRef]
- Baillie, G.D.; Tejeda, G.S.; Kelly, M.P. Therapeutic targeting of 3′,5′-cyclic nucleotide phosphodiesterases: Inhibition and beyond. Nat. Rev. Drug Discov. 2019, 18, 770–796. [Google Scholar] [CrossRef]
- Ala, M.; Mohammad Jafari, R.; Dehpour, A.R. Sildenafil beyond erectile dysfunction and pulmonary arterial hypertension: Thinking about new indications. Fundam. Clin. Pharmacol. 2021, 35, 235–259. [Google Scholar] [CrossRef]
- Pușcașu, C.; Zanfirescu, A.; Negreș, S.; Șeremet, O.C. Exploring the multifaceted potential of sildenafil in medicine. Medicina 2023, 59, 2190. [Google Scholar] [CrossRef] [PubMed]
- Kass, D.A.; Champion, H.C.; Beavo, J.A. Phosphodiesterase type 5: Expanding roles in cardiovascular regulation. Circ. Res. 2007, 101, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.S.; Geethakumari, A.M.; Biswas, K.H. Phosphodiesterase 5 (PDE5): Structure-function regulation and therapeutic applications of inhibitors. Biomed. Pharmacother. 2021, 134, 111128. [Google Scholar] [CrossRef] [PubMed]
- Menniti, F.S.; Ren, J.; Coskran, T.M.; Liu, J.; Morton, D.; Sietsma, D.K.; Som, A.; Stephenson, D.T.; Tate, B.A.; Finklestein, S.P. Phosphodiesterase 5A inhibitors improve functional recovery after stroke in rats: Optimized dosing regimen with implications for mechanism. J. Pharmacol. Exp. Ther. 2009, 331, 842–850. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Zhang, L.; Zhang, Z.; Tsang, W.; Lu, M.; Zhang, L.; Chopp, M. Sildenafil (Viagra) induces neurogenesis and promotes functional recovery after stroke in rats. Stroke 2002, 33, 2675–2680. [Google Scholar] [CrossRef]
- Marte, A.; Pepicelli, O.; Cavallero, A.; Raiteri, M.; Fedele, E. In vivo effects of phosphodiesterase inhibition on basal cyclic guanosine monophosphate levels in the prefrontal cortex, hippocampus and cerebellum of freely moving rats. J. Neurosci. Res. 2008, 86, 3338–3347. [Google Scholar] [CrossRef]
- Puerta, E.; Hervias, I.; Goñi-Allo, B.; Lasheras, B.; Jordan, J.; Aguirre, N. Phosphodiesterase 5 inhibitors prevent 3,4-methylenedioxymethamphetamine-induced 5-HT deficits in the rat. J. Neurochem. 2009, 108, 755–766. [Google Scholar] [CrossRef]
- Lorenc-Koci, E.; Czarnecka, A.; Lenda, T.; Kamińska, K.; Konieczny, J. Molsidomine, a nitric oxide donor, modulates rotational behavior and monoamine metabolism in 6-OHDA lesioned rats treated chronically with L-DOPA. Neurochem. Int. 2013, 63, 790–804. [Google Scholar] [CrossRef]
- Schwarting, R.K.W.; Huston, J.P. The unilateral 6-hydroxydopamine lesion model in behavioral brain research. Analysis of functional deficits, recovery and treatments. Prog. Neurobiol. 1996, 50, 275–331. [Google Scholar] [CrossRef]
- Duty, S.; Jenner, P. Animal models of Parkinson’s disease: A source of novel treatments and clues to the cause of the reduction of striatal dopamine levels: An in vivo microdialysis study in 6-hydroxydopamine-lesioned rats. J. Neurochem. 2011, 112, 444–452. [Google Scholar]
- Tronci, E.; Francardo, V. Animal models of l-DOPA-induced dyskinesia: The 6-OHDA-lesioned rat and mouse. J. Neural. Transm. 2018, 125, 1137–1144. [Google Scholar] [CrossRef]
- Arai, R.; Karasawa, N.; Geffard, M.; Nagatsu, T.; Nagatsu, I. Immunohistochemical evidence that central serotonin neurons produce dopamine from exogenous L-DOPA in the rat, with reference to the involvement of aromatic L-amino acid decarboxylase. Brain Res. 1994, 667, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Arai, R.; Karasawa, N.; Geffard, M.; Nagatsu, I. L-DOPA is converted to dopaminę in serotonergic fibers of the striatum of the rat: A double-labeling immunofluorescence study. Neurosci. Lett. 1995, 195, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Kannari, K.; Maeda, T.; Tomiyama, M.; Suda, T.; Matsunaga, M. Role of serotonergic neurons in L-DOPA-derived extracellular dopamine in the striatum of 6-OHDA-lesioned rats. NeuroReport 1999, 10, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Nagata, K.; Yoshida, Y.; Kannari, K. Serotonergic hyperinnervation into the dopaminergic denervated striatum compensates for dopamine conversion from exogenously administered L-DOPA. Brain Res. 2005, 1046, 230–233. [Google Scholar] [CrossRef]
- Kannari, K.; Shen, H.; Arai, A.; Tomiyama, M.; Baba, M. Reuptake of L-DOPA derived extracellular dopamine in the striatum with dopaminergic denervation via serotonin transporters. Neurosci. Lett. 2006, 402, 62–65. [Google Scholar] [CrossRef]
- Dray, A.; Gonye, T.J.; Oakley, N.R.; Tanner, T. Evidence for the existence of raphe projection to the substantia nigra. Brain Res. 1976, 113, 45–57. [Google Scholar] [CrossRef]
- van der Kooy, D.; Hattori, T. Dorsal raphe cells with collateral projections to the caudate-putamen and substantia nigra: A fluorescent retrograde double labeling study in the rat. Brain Res. 1980, 186, 1–7. [Google Scholar] [CrossRef]
- Steinbusch, H.W.; van der Kooy, D.; Verhofstad, A.A.; Pellegrino, A. Serotonergic and non-serotonergic projections from the nucleus raphe dorsalis to the caudate putamen complex in the rat, studied by a combined immunofluorescence and fluorescent retrograde axonal labeling technique. Neurosci. Lett. 1980, 19, 137–142. [Google Scholar] [CrossRef]
- Imai, H.; Steindler, D.A.; Kitai, S.T. The organization of divergent axonal projections from the midbrain raphe nuclei in the rat. J. Comp. Neurol. 1986, 243, 363–380. [Google Scholar] [CrossRef]
- Robertson, G.S.; Robertson, H.A. Evidence that L-dopa-induced rotational behavior is dependent on both striatal and nigral mechanisms. J. Neurosci. 1989, 9, 3326–3331. [Google Scholar] [CrossRef] [PubMed]
- Chastan, N.; Westby, G.W.; Yelnik, J.; Bardinet, E.; Do, M.C.; Agid, Y.; Welter, M.L. Effects of nigral stimulation on locomotion and postural stability in patients with Parkinson’s disease. Brain 2009, 132 Pt 1, 172–184. [Google Scholar] [CrossRef]
- Navailles, S.; Lagiére, M.; Contini, A.; De Deurwaerdére, P. Multisite intracerebral microdialysis to study the mechanism of L-DOPA induced dopamine and serotonin release in the parkinsonian brain. ACS Chem. Neurosci. 2013, 4, 680–692. [Google Scholar] [CrossRef]
- De Deurwaerdére, P.; Di Giovanni, G.; Millan, M.J. Expanding the repertoire of LDOPA’s actions: A comprehensive review of its functional neurochemistry. Prog. Neurobiol. 2017, 151, 57–100. [Google Scholar] [CrossRef]
- Jin, F.; Gong, Q.H.; Xu, Y.S.; Wang, L.N.; Jin, H.; Li, F.; Li, L.S.; Ma, Y.W.; Shi, J.S. Icariin, a phoshphodiesterase-5 inhibitor, improves learning and memory in APP/PS1 transgenic mice by stimulation of NO/cGMP signaling. Int. J. Neuropsychopharmacol. 2014, 17, 871–881. [Google Scholar] [CrossRef]
- Puzzo, D.; Loreto, C.; Giunta, S.; Musumeci, G.; Frasca, G.; Podda, M.V.; Palmeri, A. Effect of phosphodiesterase-5 inhibition on apoptosis and beta amyloid load in aged mice. Neurobiol. Aging 2014, 35, 520–531. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, J.Y.; Xue, X.; Dong, Y.X.; Miao, F.R.; Wang, Y.F.; Wu, C.F. A novel phosphodiesterase-5 inhibitor: Yonkenafil modulates neurogenesis, gliosis to improve cognitive function and ameliorates amyloid burden in an APP/PS1 transgenic mice model. Mech. Ageing Dev. 2015, 150, 34–45. [Google Scholar] [CrossRef]
- Kelly, P.H.; Iversen, S.D. Selective 6-OHDA-induced destruction of mesolimbic dopamine neurons: Abolition of psychostimulant-induced locomotor activity in rats. Eur. J. Pharmacol. 1976, 40, 45–56. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1986. [Google Scholar]
- Fornari, R.V.; Wichmann, R.; Atsak, P.; Atucha, E.; Barsegyan, A.; Beldjoud, H.; Messanvi, F.; Thuring, C.M.; Roozendaal, B. Rodent stereotaxic surgery and animal welfare outcome improvements for behavioral neuroscience. J. Vis. Exp. 2012, 59, 3528. [Google Scholar]
- Czarnecka, A.; Lenda, T.; Domin, H.; Konieczny, J.; Śmiałowska, M.; Lorenc-Koci, E. Alterations in the expression of nNOS in the substantia nigra and subthalamic nucleus of 6-OHDA-lesioned rats: The effects of chronic treatment with L-DOPA and the nitric oxide donor, molsidomine. Brain Res. 2013, 1541, 92–105. [Google Scholar] [CrossRef]
- Ungerstedt, U. Postsynaptic supersensitivity after 6-hydroxy-dopamine induced degeneration of the nigro-striatal dopamine system. Acta Physiol. Scand. 1971, 367, 69–93. [Google Scholar] [CrossRef]
- Frau, L.; Morelli, M.; Simola, N. Performance of movement in hemiparkinsonian rats influences the modification induced by dopamine agonists in striatal efferent dynorphinergic neurons. Exp. Neurol. 2013, 247, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Janis, K.L.; Brennan, R.T.; Drolet, R.E.; Behrouz, B.; Kaufman, S.K.; Lookingland, K.J.; Goudreau, J.L. Effects of sildenafil on nigrostriatal dopamine neurons in a murine model of Parkinson’s disease. J. Alzheimers Dis. 2008, 15, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Kamińska, K.; Lenda, T.; Konieczny, J.; Czarnecka, A.; Lorenc-Koci, E. Depressive like neurochemical and behavioral markerrs of Parkinson’s disease after 6-OHDA administration unilaterally to the rat medial forebrain bundle. Pharmacol. Rep. 2017, 69, 985–994. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenc-Koci, E.; Kamińska, K.; Lenda, T.; Konieczny, J. The Effect of Chronic Treatment with the Inhibitor of Phosphodiesterase 5 (PDE5), Sildenafil, in Combination with L-DOPA on Asymmetric Behavior and Monoamine Catabolism in the Striatum and Substantia Nigra of Unilaterally 6-OHDA-Lesioned Rats. Molecules 2024, 29, 4318. https://doi.org/10.3390/molecules29184318
Lorenc-Koci E, Kamińska K, Lenda T, Konieczny J. The Effect of Chronic Treatment with the Inhibitor of Phosphodiesterase 5 (PDE5), Sildenafil, in Combination with L-DOPA on Asymmetric Behavior and Monoamine Catabolism in the Striatum and Substantia Nigra of Unilaterally 6-OHDA-Lesioned Rats. Molecules. 2024; 29(18):4318. https://doi.org/10.3390/molecules29184318
Chicago/Turabian StyleLorenc-Koci, Elżbieta, Kinga Kamińska, Tomasz Lenda, and Jolanta Konieczny. 2024. "The Effect of Chronic Treatment with the Inhibitor of Phosphodiesterase 5 (PDE5), Sildenafil, in Combination with L-DOPA on Asymmetric Behavior and Monoamine Catabolism in the Striatum and Substantia Nigra of Unilaterally 6-OHDA-Lesioned Rats" Molecules 29, no. 18: 4318. https://doi.org/10.3390/molecules29184318