

The Mechanism of Aniline Blue Degradation by Short-Chain Dehydrogenase (SDRz) in Comamonas testosteroni

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Biodegradation Ability of Different Strains to Aniline Blue
2.2. Optimization of Conditions for Aniline Blue Biodegradation by Strain CT1
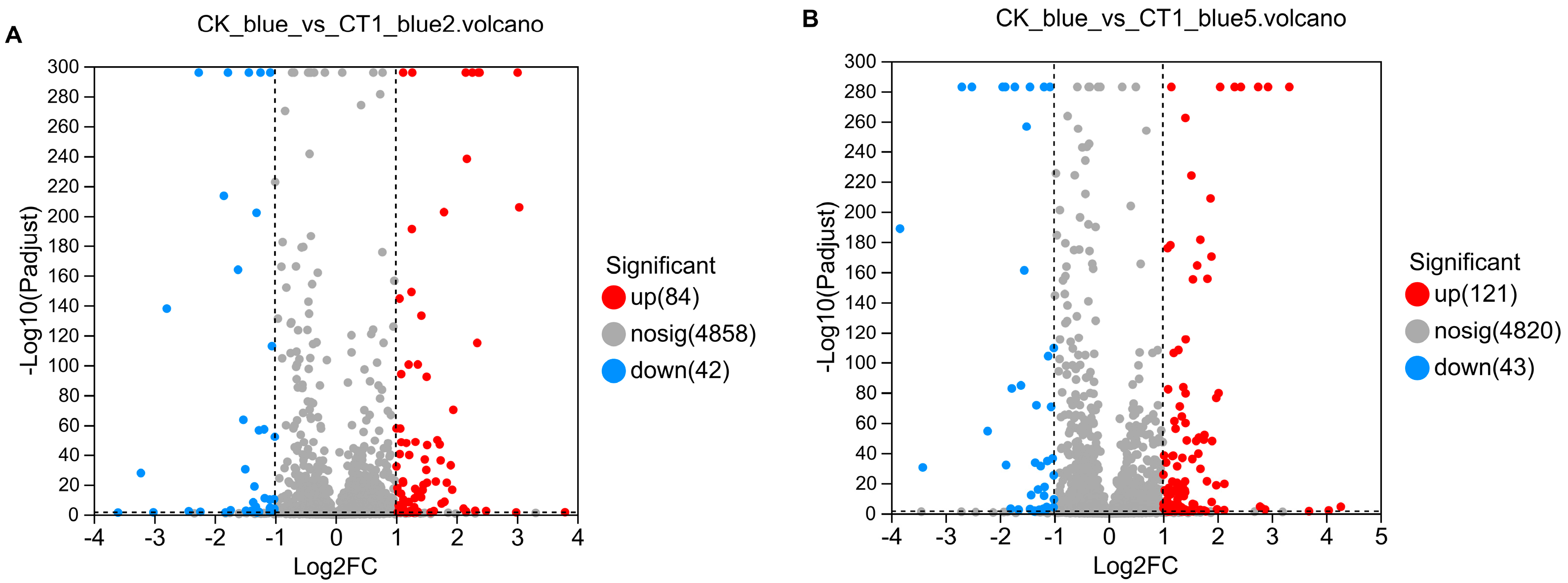
2.3. Screening of Key Genes Involved in Aniline Blue Degradation
2.4. Homology Analysis of SDRz
2.5. Construction of SDRz-Expressing Strain
2.6. Construction of SDRz Knockout Mutant and Gene Functional Verification
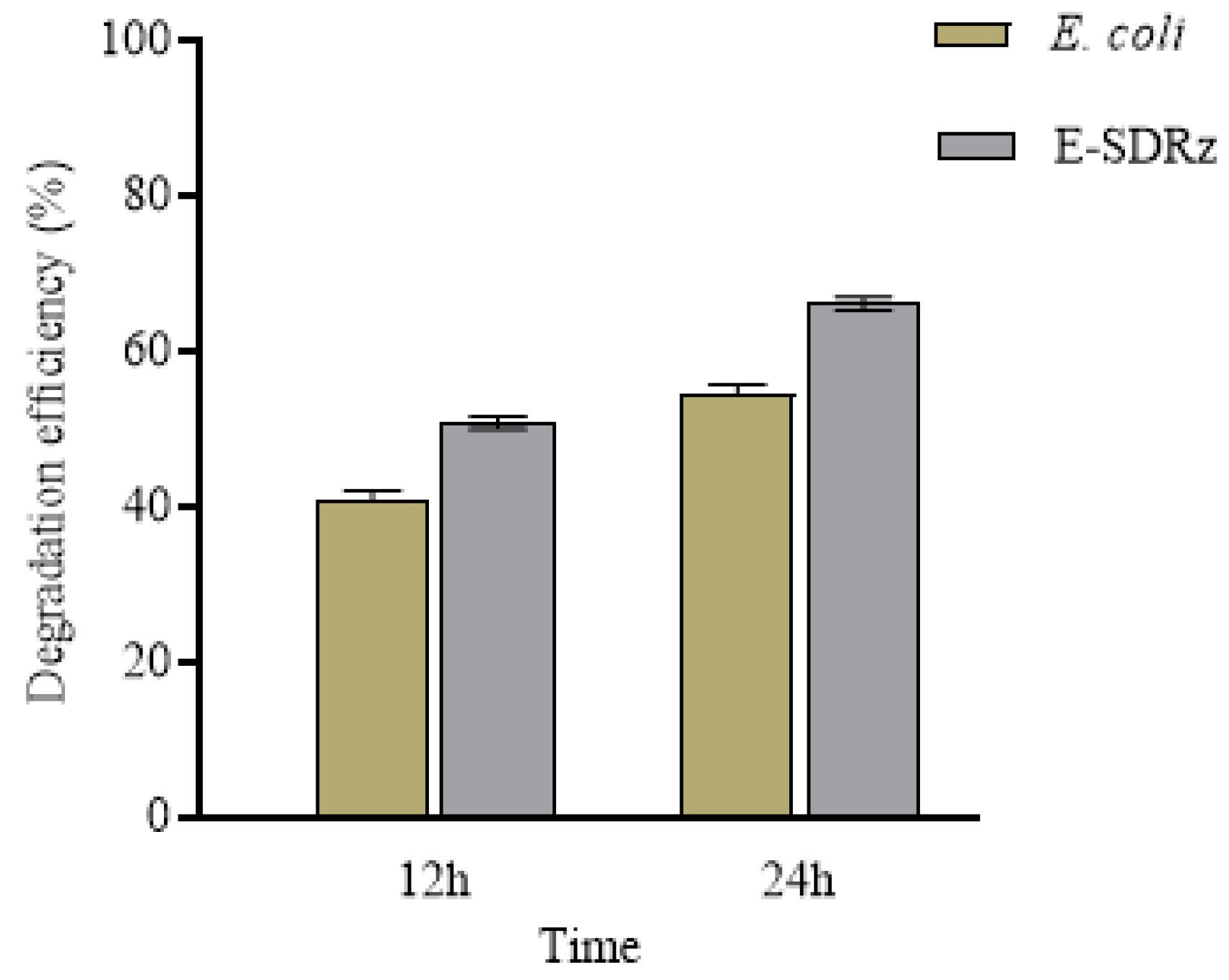
2.7. Expression and Purification of rSDRz
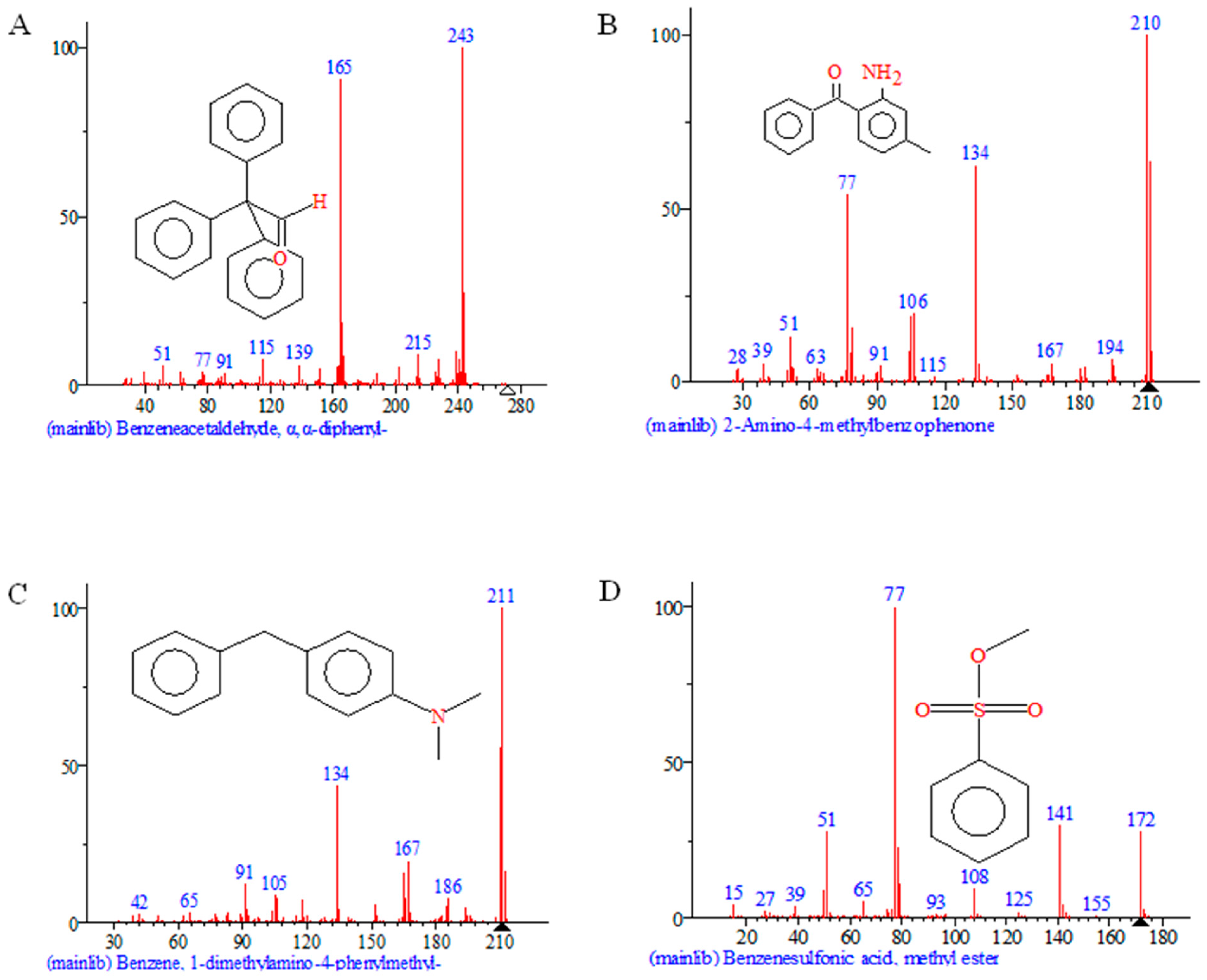
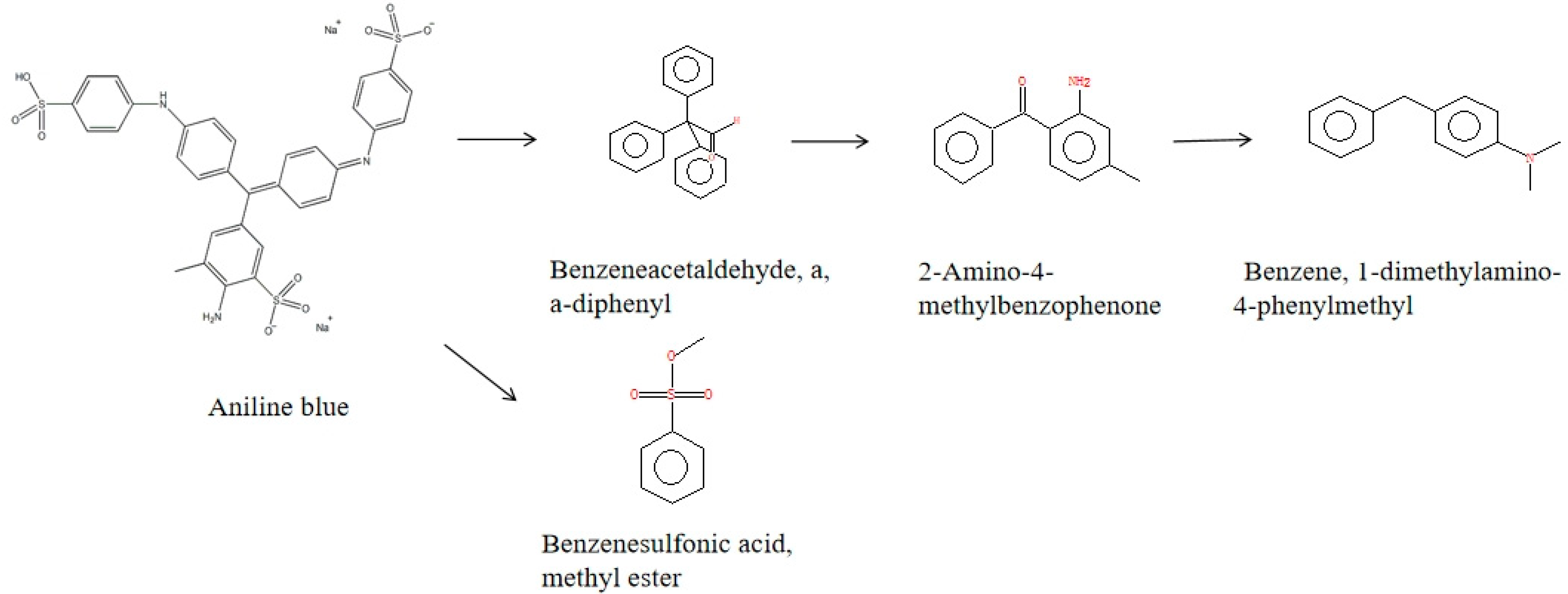
2.8. Analysis of Aniline Blue Degradation Products by rSDRz
2.9. Environmental Effects and Prospects of Aniline Blue Biodegradation
3. Materials and Methods
3.1. Experimental Materials
3.2. Experimental Methods
3.2.1. Biological Degradation of Aniline Blue by Different Strains
3.2.2. Optimization of Aniline Blue Biodegradation Conditions
3.2.3. Prokaryotic Transcriptomic Analysis of Aniline Blue Degradation by CT1
3.2.4. Construction of SDRz-Expressing Strains
3.2.5. Construction of SDRz Knockout Strains
3.2.6. Construction of the SDRz Protein Expression Vector
3.2.7. Induction and Purification of rSDRz
3.2.8. Functional Verification of SDRz
3.2.9. Enzymatic Properties of rSDRz
3.2.10. Detection of Aniline Blue Degradation Products by GC-MS
3.2.11. Toxicity Analysis of Aniline Blue Degradation Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aragaw, T.A. A review of dye biodegradation in textile wastewater, challenges due to wastewater characteristics, and the potential of alkaliphiles. J. Hazard. Mater. Adv. 2024, 16, 100493. [Google Scholar] [CrossRef]
- You, X.; Li, E.; Liu, J.; Li, S. Using Natural Biomacromolecules for Adsorptive and Enzymatic Removal of Aniline Blue from Water. Molecules 2018, 23, 1606. [Google Scholar] [CrossRef] [PubMed]
- Ogugbue, C.J.; Sawidis, T. Bioremediation and Detoxification of Synthetic Wastewater Containing Triarylmethane Dyes by Aeromonas hydrophila Isolated from Industrial Effluent. Biotechnol. Res. Int. 2011, 2011, 967925. [Google Scholar] [CrossRef] [PubMed]
- Husain, Q. Potential applications of the oxidoreductive enzymes in the decolorization and detoxification of textile and other synthetic dyes from polluted water: A review. Crit. Rev. Biotechnol. 2006, 26, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Azzam, A.B.; El-Sheikh, S.M.; Geioushy, R.A.; Salah, B.A.; El-Dars, F.M.; Helal, A.S. Facile fabrication of a novel BiPO4 phase junction with enhanced photocatalytic performance towards aniline blue degradation. RSC Adv. 2019, 9, 17246–17253. [Google Scholar] [CrossRef]
- Usman, M.A.; Aftab, R.A.; Zaidi, S.; Adnan, S.M.; Rao, R.A.K. Adsorption of aniline blue dye on activated pomegranate peel: Equilibrium, kinetics, thermodynamics and support vector regression modelling. Int. J. Environ. Sci. Technol. 2022, 19, 8351–8368. [Google Scholar] [CrossRef]
- Guo, J.; Zhou, J.; Wang, D.; Tian, C.; Wang, P.; Uddin, M.S. A novel moderately halophilic bacterium for decolorizing azo dye under high salt condition. Biodegradation 2008, 19, 15–19. [Google Scholar] [CrossRef]
- Lü, C.; Luo, X.F.; Dong, X.J.; Peng, J.Q.; Cao, F. New cost-effective mediator enhanced enzymatic degradation of aniline blue. J. Environ. Biol. 2021, 42, 99–105. [Google Scholar] [CrossRef]
- Cheng, C.; Luo, Z.; Chen, B.; Xie, Y.; Liu, X.; Zhang, D.; Pan, C. Biodegradation of aniline blue dye by salt-tolerant Bacillus thuringiensis DHC4 isolated from soil-feeding termite guts. J. Clean. Prod. 2024, 457, 142489. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Liu, J.; Li, R.; Shen, B. Isolation of a thermophilic bacterium, Geobacillus sp. SH-1, capable of degrading aliphatic hydrocarbons and naphthalene simultaneously, and identification of its naphthalene degrading pathway. Bioresour. Technol. 2012, 124, 83–89. [Google Scholar] [CrossRef]
- Zeinali, M.; Vossoughi, M.; Ardestani, S.K. Naphthalene metabolism in Nocardia otitidiscaviarum strain TSH1, a moderately thermophilic microorganism. Chemosphere 2008, 72, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Zeinali, M.; Vossoughi, M.; Ardestani, S.K. Degradation of phenanthrene and anthracene by Nocardia otitidiscaviarum strain TSH1, a moderately thermophilic bacterium. J. Appl. Microbiol. 2008, 105, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Menn, F.M.; Applegate, B.M.; Sayler, G.S. NAH plasmid-mediated catabolism of anthracene and phenanthrene to naphthoic acids. Applenvironmicrobiol 1993, 59, 1938. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Li, C.; Xu, Y.X.; Wang, Q.; Ma, X.; Chen, S.W.; Yu, J.; Yang, Y. Isolation of a Microorganic Strain for the High Volume Degradation of Aniline Blue and Its Application in Natural Sewage Treatment. Comput. Mol. Biosci. 2018, 8, 149–165. [Google Scholar] [CrossRef]
- Orsini, J.; Tam, E.; Hauser, N.; Rajayer, S. Polymicrobial Bacteremia Involving Comamonas testosteroni. Case Rep. Med. 2014, 2014, 578127. [Google Scholar] [CrossRef]
- Wang, Y.H.; Huang, Z.; Liu, S.J. Chemotaxis Towards Aromatic Compounds: Insights from Comamonas testosteroni. Int. J. Mol. Sci. 2019, 20, 2701. [Google Scholar] [CrossRef]
- Sammoni, A.; Abdalah, A.; Al-Aissami, M. Comamonas testosteroni bacteremia: A rare unusual pathogen detected in a burned patient: Case report and literature review. Ann. Med. Surg. (Lond). 2022, 75, 103371. [Google Scholar] [CrossRef]
- Lu, Q.; Sun, X.; Jiang, Z.; Cui, Y.; Li, X.; Cui, J. Effects of Comamonas testosteroni on dissipation of polycyclic aromatic hydrocarbons and the response of endogenous bacteria for soil bioremediation. Environ. Sci. Pollut. Res. Int. 2022, 29, 82351–82364. [Google Scholar] [CrossRef]
- Vural, C.; Diallo, M.M.; Ozdemir, G. Assessment of Comamonas testosteroni strain PT9 as a rapid phthalic acid degrader for industrial wastewaters. J. Basic Microbiol. 2022, 62, 508–517. [Google Scholar] [CrossRef]
- Zhu, G.; Zhang, Y.; Chen, S.; Wang, L.; Zhang, Z.; Rittmann, B.E. How bioaugmentation with Comamonas testosteroni accelerates pyridine mono-oxygenation and mineralization. Environ. Res. 2021, 193, 110553. [Google Scholar] [CrossRef]
- Xu, M.; Yang, X.; Zhang, J.; Liu, D.; Zhang, C.; Wu, M.; Musazade, E.; Maser, E.; Xiong, G.; Guo, L. The mechanism of anthracene degradation by tryptophan -2,3-dioxygenase (T23D) in Comamonas testosteroni. Chem.-Biol. Interact. 2024, 393, 110950. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Liu, Y.; Li, H.; Yang, X.; Yue, W.; Zhang, Y.; Liu, D.; Wu, M.; Wang, D.; Xiong, G.; et al. Anthracene degradation involved by antibiotic biosynthesis monooxygenase (ABM) in Comamonas testosteroni. Int. Biodeterior. Biodegrad. 2024, 190, 105790. [Google Scholar] [CrossRef]
- Xu, M.; Wu, M.; Zhang, Y.; Zhang, H.; Liu, W.; Chen, G.; Xiong, G.; Guo, L. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by bacterial mixture. Int. J. Environ. Sci. Technol. 2022, 19, 3833–3844. [Google Scholar] [CrossRef]
- Wang, P.; Zheng, D.; Peng, W.; Wang, Y.; Wang, X.; Xiong, W.; Liang, R. Characterization of 17β-hydroxysteroid dehydrogenase and regulators involved in estrogen degradation in Pseudomonas putida SJTE-1. Appl. Microbiol. Biotechnol. 2019, 103, 2413–2425. [Google Scholar] [CrossRef]
- Stambergova, H.; Skarydova, L.; Dunford, J.E.; Wsol, V. Biochemical properties of human dehydrogenase/reductase (SDR family) member 7. Chem.-Biol. Interact. 2014, 207, 52–57. [Google Scholar] [CrossRef]
- Kavanagh, K.L.; Jörnvall, H.; Persson, B.; Oppermann, U. Medium- and short-chain dehydrogenase/reductase gene and protein families: The SDR superfamily: Functional and structural diversity within a family of metabolic and regulatory enzymes. Cell. Mol. Life Sci. CMLS 2008, 65, 3895–3906. [Google Scholar] [CrossRef]
- Ebert, B.; Kisiela, M.; Maser, E. Transcriptional regulation of human and murine short-chain dehydrogenase/reductases (SDRs) —An in silico approach. Drug Metab. Rev. 2016, 48, 183–217. [Google Scholar] [CrossRef]
- Gong, W.; Kisiela, M.; Schilhabel, M.B.; Xiong, G.; Maser, E. Genome sequence of Comamonas testosteroni ATCC 11996, a representative strain involved in steroid degradation. J. Bacteriol. 2012, 194, 1633–1634. [Google Scholar] [CrossRef]
- Bhatia, C.; Oerum, S.; Bray, J.; Kavanagh, K.L.; Shafqat, N.; Yue, W.; Oppermann, U. Towards a systematic analysis of human short-chain dehydrogenases/reductases (SDR): Ligand identification and structure–activity relationships. Chem.-Biol. Interact. 2015, 234, 114–125. [Google Scholar] [CrossRef]
- Ji, W.; Chen, Y.; Zhang, H.; Zhang, X.; Li, Z.; Yu, Y. Cloning, expression and characterization of a putative 7alpha-hydroxysteroid dehydrogenase in Comamonas testosteroni. Microbiol. Res. 2014, 169, 148–154. [Google Scholar] [CrossRef]
- Bhatia, C. Investigation into Structural and Functional Relationships of Short-Chain Dehydrogenases and Reductases (SDRs) Using a Compound Library. Ph.D. Thesis, University of Oxford, Oxford, UK, 2018. [Google Scholar]
- Schleheck, D.; Knepper, T.P.; Fischer, K.; Cook, A.M. Mineralization of individual congeners of linear alkylbenzenesulfonate by defined pairs of heterotrophic bacteria. Appl. Environ. Microbiol. 2004, 70, 4053–4063. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Denger, K.; Huhn, T.; Schleheck, D. Two enzymes of a complete degradation pathway for linear alkylbenzenesulfonate (LAS) surfactants: 4-sulfoacetophenone Baeyer-Villiger monooxygenase and 4-sulfophenylacetate esterase in Comamonas testosteroni KF-1. Appl. Environ. Microbiol. 2012, 78, 8254–8263. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Kesberg, A.I.; Labutti, K.M.; Pitluck, S.; Bruce, D.; Hauser, L.; Copeland, A.; Woyke, T.; Lowry, S.; Lucas, S.; et al. Permanent draft genome sequence of Comamonas testosteroni KF-1. Stand. Genom. Sci. 2013, 8, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xiao, X.; Xu, C.; Cao, D.; Du, D. Decolorization and detoxification of a sulfonated triphenylmethane dye aniline blue by Shewanella oneidensis MR-1 under anaerobic conditions. Appl. Microbiol. Biotechnol. 2013, 97, 7439–7446. [Google Scholar] [CrossRef]
- Yonghui, L.; Liang, C.; Xingbing, H.E.; Yaoqing, H.E.; Xia, Z. Biodegradation of Aniline Blue Dye by a Fungus Mucoromycotina sp. HS-3Mucoromycotina sp. HS-3. Microbiol. China 2010, 37, 1727–1733. [Google Scholar]
- Emtiazi, G.; Satarii, M.; Mazaherion, F. The utilization of aniline, chlorinated aniline, and aniline blue as the only source of nitrogen by fungi in water. Water Res. 2001, 35, 1219–1224. [Google Scholar] [CrossRef]
- Ma, Y.; Li, L.; Lu, M.; He, J. Isolation of Aniline Blue-discoloring Actinomyces and its decolorization characteristics. Acta Sci. Circumstantiae 2016, 36, 4361–4366. [Google Scholar]
- Filling, C.; Berndt, K.D.; Benach, J.; Knapp, S.; Prozorovski, T.; Nordling, E.; Ladenstein, R.; Jörnvall, H.; Oppermann, U. Critical residues for structure and catalysis in short-chain dehydrogenases/reductases. J. Biol. Chem. 2002, 277, 25677–25684. [Google Scholar] [CrossRef]
- Hildebrandt, P.; Musidlowska, A.; Bornscheuer, U.T.; Altenbuchner, J. Cloning, functional expression and biochemical characterization of a stereoselective alcohol dehydrogenase from Pseudomonas fluorescens DSM50106. Appl. Microbiol. Biotechnol. 2002, 59, 483–487. [Google Scholar]
- Kleiger, G.; Eisenberg, D. GXXXG and GXXXA motifs stabilize FAD and NAD(P)-binding Rossmann folds through C(alpha)-H... O hydrogen bonds and van der waals interactions. J. Mol. Biol. 2002, 323, 69–76. [Google Scholar] [CrossRef]
- Murugesan, K.; Yang, I.H.; Kim, Y.M.; Jeon, J.R.; Chang, Y.S. Enhanced transformation of malachite green by laccase of Ganoderma lucidum in the presence of natural phenolic compounds. Appl. Microbiol. Biotechnol. 2009, 82, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, Y.; Park, H.J.; Lee, J.S.; Kwak, S.N.; Jung, W.H.; Lee, S.G.; Kim, D.; Lee, Y.C.; Oh, T.K. Structural insight into bioremediation of triphenylmethane dyes by Citrobacter sp. triphenylmethane reductase. J. Biol. Chem. 2008, 283, 31981–31990. [Google Scholar] [CrossRef] [PubMed]
- Navada, K.K.; Kulal, A. Enhanced production of laccase from gamma irradiated endophytic fungus: A study on biotransformation kinetics of aniline blue and textile effluent decolourisation. J. Environ. Chem. Eng. 2020, 8, 103550. [Google Scholar] [CrossRef]
- Shedbalkar, U.; Dhanve, R.; Jadhav, J. Biodegradation of triphenylmethane dye cotton blue by Penicillium ochrochloron MTCC 517. J. Hazard. Mater. 2008, 157, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Qanungo, K. Removal of aniline blue from aqueous solution using adsorption: A mini review. Mater. Today Proc. 2021, 37, 2290–2293. [Google Scholar] [CrossRef]
- Mishra, S.; Maiti, A. The efficacy of bacterial species to decolourise reactive azo, anthroquinone and triphenylmethane dyes from wastewater: A review. Environ. Sci. Pollut. Res. 2018, 25, 8286–8314. [Google Scholar] [CrossRef]
- Aruna, B.; Lodi, S.; Shiva Kumar, E.; Rani, P.; Prasad, D.V.; VijayaLakshmi, D. Decolorization of Acid Blue 25 dye by individual and mixed bacterial consortium isolated from textile effluents. Int. J. Curr. Microbiol. App. Sci. 2015, 4, 1015–1024. [Google Scholar]
- Gong, W.; Xiong, G.; Maser, E. Oligomerization and negative autoregulation of the LysR-type transcriptional regulator HsdR from Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2012, 132, 203–211. [Google Scholar] [CrossRef]
- Matsunaga, T.; Shintani, S.; Hara, A. Multiplicity of mammalian reductases for xenobiotic carbonyl compounds. Drug Metab. Pharmacokinet. 2006, 21, 1–18. [Google Scholar] [CrossRef]
- Menzel, R.; Yeo, H.L.; Rienau, S.; Li, S.; Steinberg, C.E.; Stürzenbaum, S.R. Cytochrome P450s and short-chain dehydrogenases mediate the toxicogenomic response of PCB52 in the nematode Caenorhabditis elegans. J. Mol. Biol. 2007, 370, 1–13. [Google Scholar] [CrossRef]
- Elgarahy, A.M.; Elwakeel, K.Z.; Mohammad, S.H.; Elshoubaky, G.A. A critical review of biosorption of dyes, heavy metals and metalloids from wastewater as an efficient and green process. Clean. Eng. Technol. 2021, 4, 100209. [Google Scholar] [CrossRef]
- Talalay, P.; Dobson, M.M.; Tapley, D.F. Oxidative degradation of testosterone by adaptive enzymes. Nature 1952, 170, 620–621. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Draus, E.; Luo, Y.; Maser, E. 3alpha-Hydroxysteroid dehydrogenase/carbonyl reductase as a tool for isolation and characterization of a new marine steroid degrading bacterial strain. Chem.-Biol. Interact. 2009, 178, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zheng, D.; Liang, R. Isolation and characterization of an estrogen-degrading Pseudomonas putida strain SJTE-1. 3 Biotech 2019, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xiong, G.; Maser, E. Characterization of the steroid degrading bacterium S19-1 from the Baltic Sea at Kiel, Germany. Chem.-Biol. Interact. 2011, 191, 83–88. [Google Scholar] [CrossRef]
- Xu, M.; He, L.; Sun, P.; Wu, M.; Cui, X.; Liu, D.; Adomako-Bonsu, A.G.; Geng, M.; Xiong, G.; Guo, L.; et al. Critical Role of Monooxygenase in Biodegradation of 2,4,6-Trinitrotoluene by Buttiauxella sp. S19-1. Molecules 2023, 28, 1969. [Google Scholar] [CrossRef]
- Li, M.; Zhao, X.; Zhang, X.; Wu, D.; Leng, S. Biodegradation of 17β-estradiol by Bacterial Co-culture Isolated from Manure. Sci. Rep. 2018, 8, 3787. [Google Scholar] [CrossRef]
- Liang, L.; Song, X.; Kong, J.; Shen, C.; Huang, T.; Hu, Z. Anaerobic biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by a facultative anaerobe Pseudomonas sp. JP1. Biodegradation 2014, 25, 825–833. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Huang, Y.; He, J.; He, L.; Zhang, J.; Yu, L.; Musazade, E.; Maser, E.; Xiong, G.; Xu, M.; et al. The Mechanism of Aniline Blue Degradation by Short-Chain Dehydrogenase (SDRz) in Comamonas testosteroni. Molecules 2024, 29, 5405. https://doi.org/10.3390/molecules29225405
Zhang C, Huang Y, He J, He L, Zhang J, Yu L, Musazade E, Maser E, Xiong G, Xu M, et al. The Mechanism of Aniline Blue Degradation by Short-Chain Dehydrogenase (SDRz) in Comamonas testosteroni. Molecules. 2024; 29(22):5405. https://doi.org/10.3390/molecules29225405
Chicago/Turabian StyleZhang, Chuanzhi, Yong Huang, Jiaxin He, Lei He, Jinyuan Zhang, Lijing Yu, Elshan Musazade, Edmund Maser, Guangming Xiong, Miao Xu, and et al. 2024. "The Mechanism of Aniline Blue Degradation by Short-Chain Dehydrogenase (SDRz) in Comamonas testosteroni" Molecules 29, no. 22: 5405. https://doi.org/10.3390/molecules29225405
APA StyleZhang, C., Huang, Y., He, J., He, L., Zhang, J., Yu, L., Musazade, E., Maser, E., Xiong, G., Xu, M., & Guo, L. (2024). The Mechanism of Aniline Blue Degradation by Short-Chain Dehydrogenase (SDRz) in Comamonas testosteroni. Molecules, 29(22), 5405. https://doi.org/10.3390/molecules29225405