
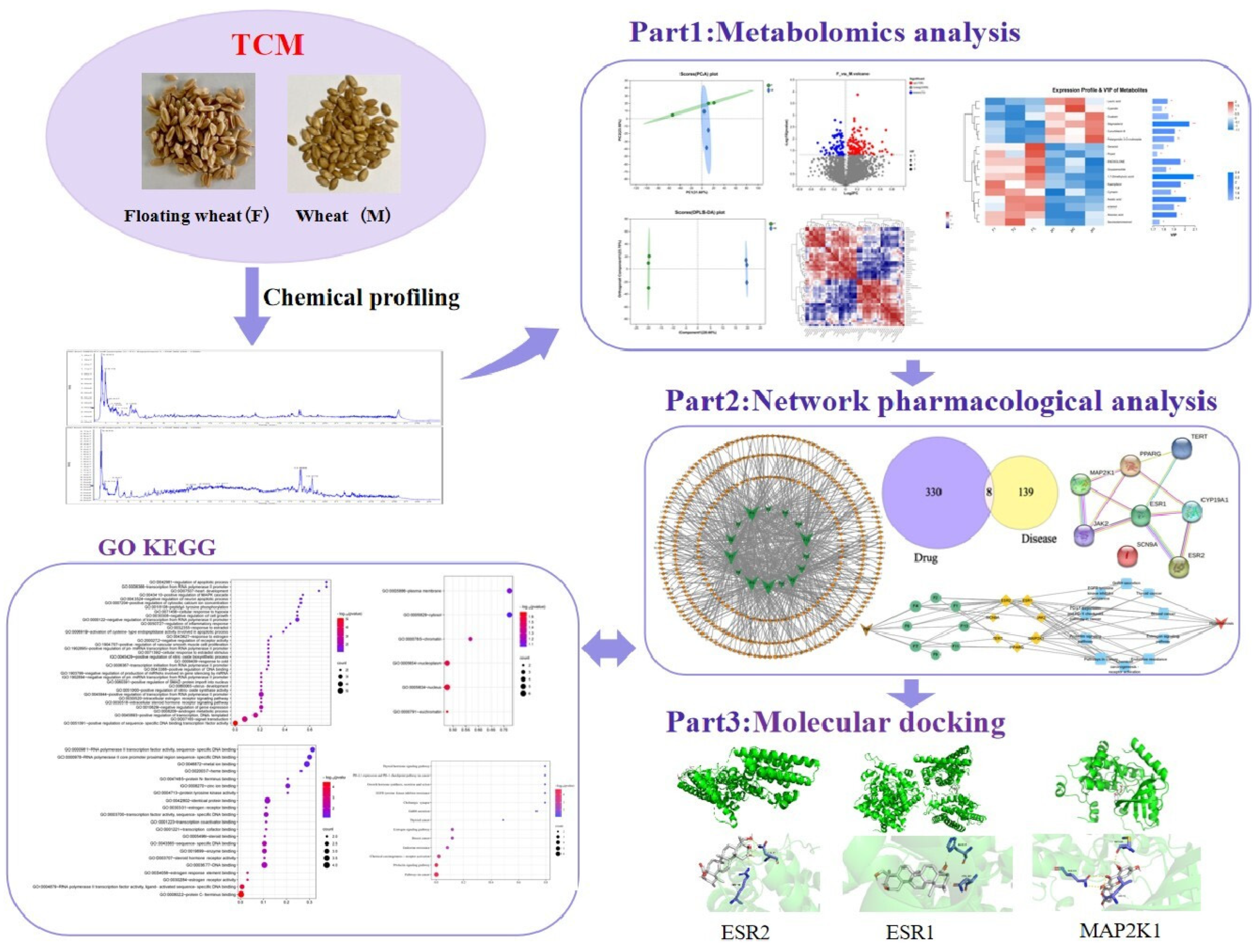
Revealing the Antiperspirant Components of Floating Wheat and Their Mechanisms of Action through Metabolomics and Network Pharmacology

Abstract
1. Introduction
2. Results and Discussion
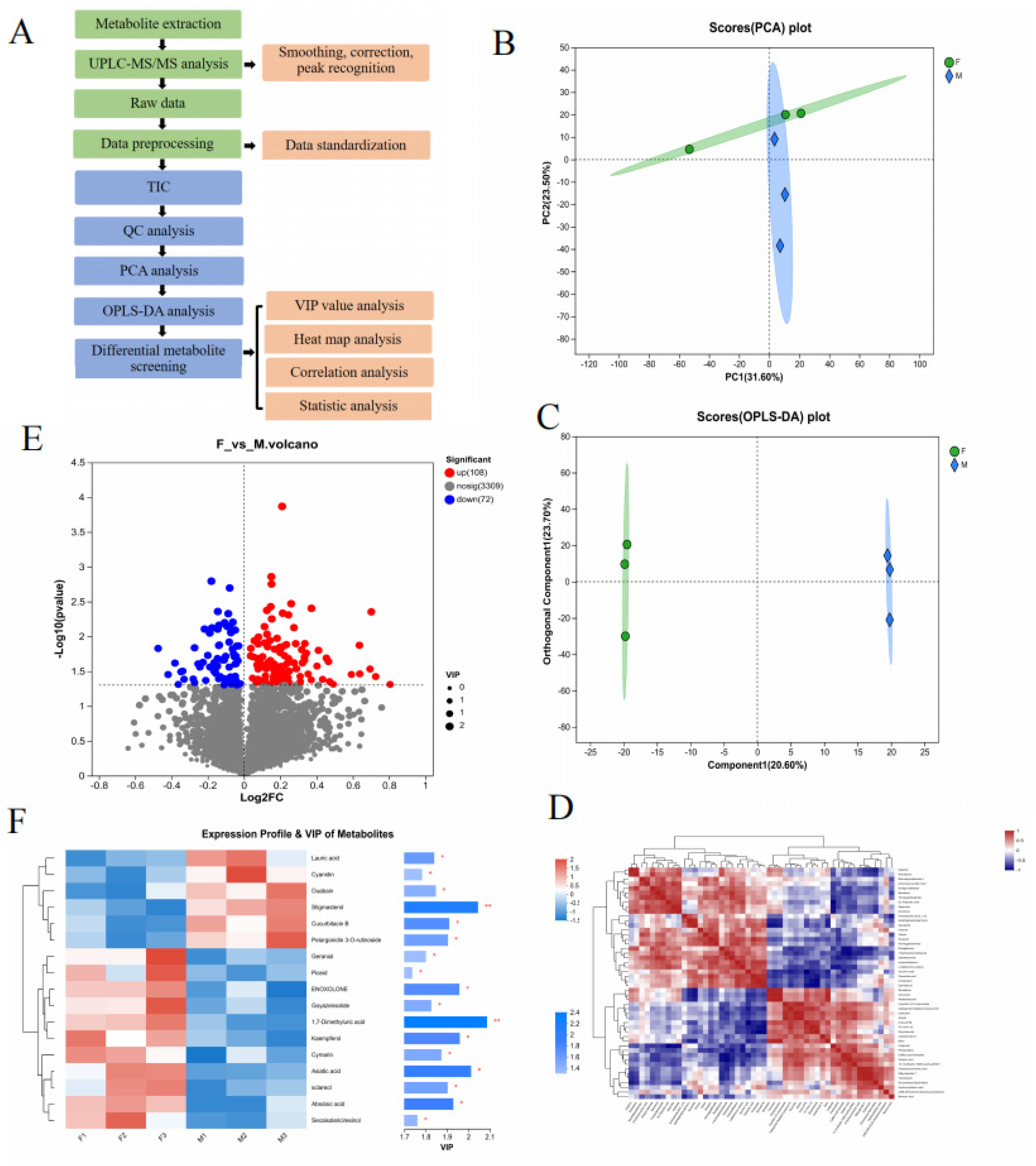
2.1. Metabolomic Analysis
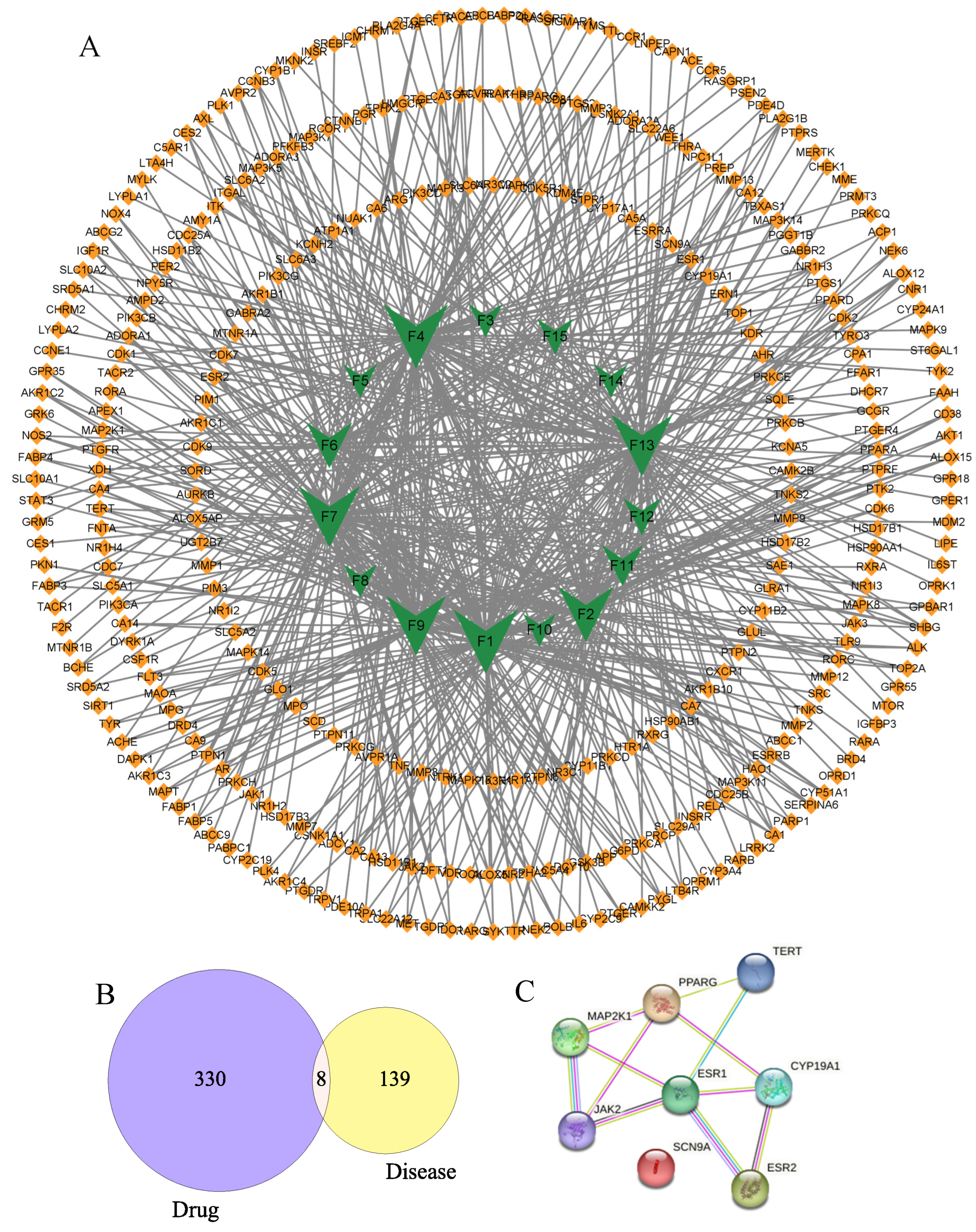
2.2. Component–Target Network Analysis
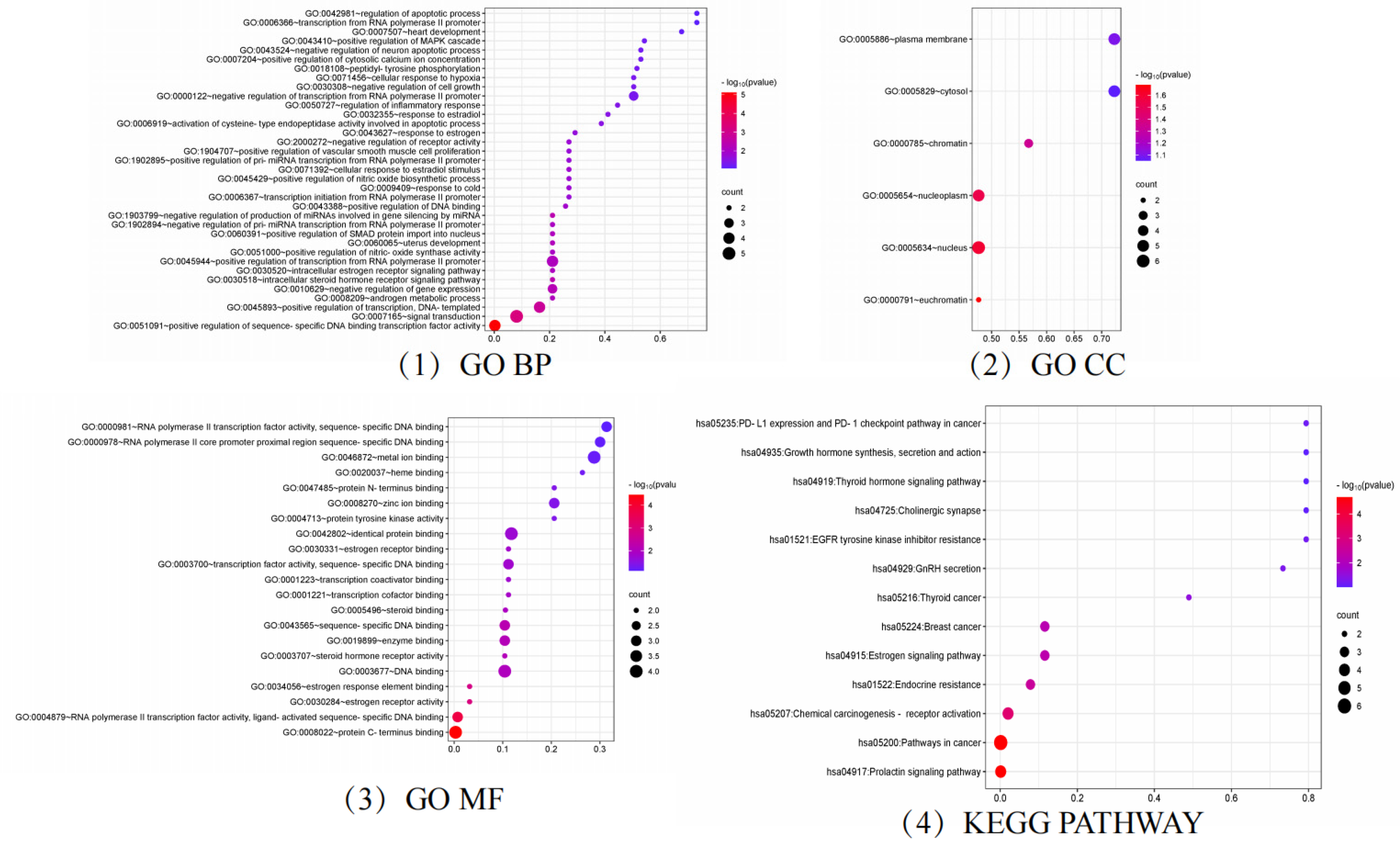
2.3. Analysis of GO and KEGG Results
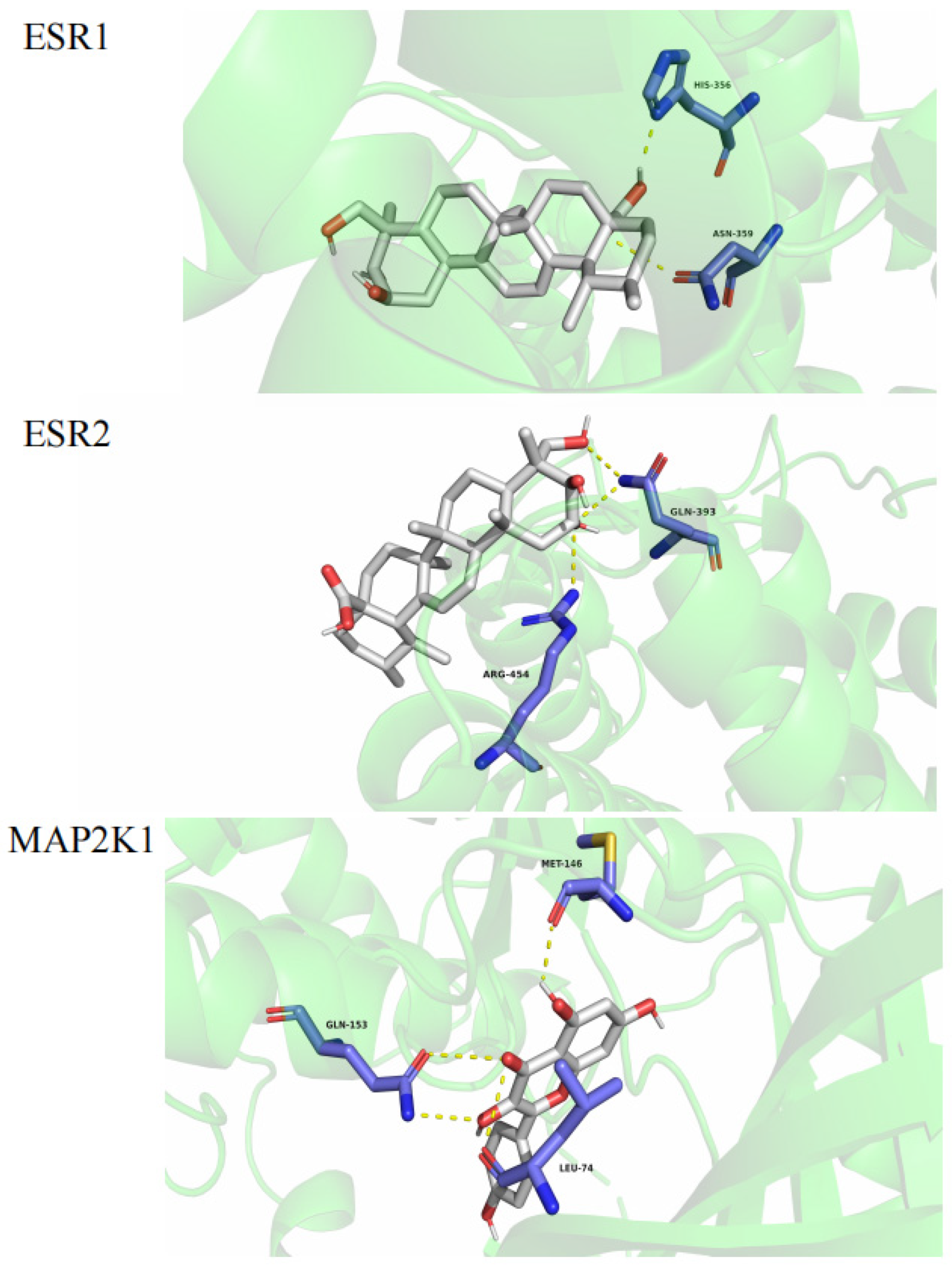
2.4. Molecular Docking Study
3. Materials and Methods
3.1. Materials and Laboratory Equipment
3.2. Sample Pretreatment
3.3. UPLC-MS/MS
3.4. Component–Target Network Construction
3.5. GO and KEGG Pathway Analyses
3.6. Molecular Docking
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cox, S.; Nasseri, R.; Rubin, R.S.; Santiago-Lastra, Y. Genitourinary Syndrome of Menopause. Med. Clin. N. Am. 2023, 107, 357–369. [Google Scholar] [CrossRef]
- Salvatore, S.; Benini, V.; Ruffolo, A.F.; Degliuomini, R.S.; Redaelli, A.; Casiraghi, A.; Candiani, M. Current challenges in the pharmacological management of genitourinary syndrome of menopause. Expert Opin. Pharmacother. 2023, 24, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, T.; Fang, S. Calibration; Henan Science and Technology Press: Zhengzhou, China, 2015. [Google Scholar]
- Tai, H.K.; Ding, Z.Z.; Zhao, S.X.; Yan, Y.Q. Modern Compendium of Materia Medica; China Medical Science and Technology Press: Beijing, China, 2001. [Google Scholar]
- Luo, T. Hygiene Treasure; China Medical Science and Technology Press: Beijing, China, 2011. [Google Scholar]
- Yuan, W. Quality Evaluation and Preliminary Analysis of Function of Floating Wheat from Different Origins; Henan University of Technology: Zhengzhou, China, 2021. [Google Scholar]
- Meng, S.; Li, H.; Yan, Y.; Zhang, L.; Cui, J.; Pei, M. Research on quality control of floating wheat herbs. Chin. J. Exp. Formul. 2012, 18, 124–127. [Google Scholar] [CrossRef]
- Chen, X. Discussion on microscopic identification of floating wheat. Med. Theory Pract. 2014, 27, 1527–1528. [Google Scholar] [CrossRef]
- Yuan, W.; Hui, M.; Tian, Q.; Huang, J. HPLC fingerprinting of floating wheat from different origins. J. Henan Univ. Technol. (Nat. Sci. Ed.) 2020, 41, 59–64. [Google Scholar] [CrossRef]
- Wu, H.; Li, X.; Yan, X.; An, L.; Luo, K.; Shao, M.; Jiang, Y.; Xie, R.; Feng, F. An untargeted metabolomics-driven approach based on LC-TOF/MS and LC-MS/MS for the screening of xenobiotics and metabolites of Zhi-Zi-Da-Huang decoction in rat plasma. J. Pharm. Biomed. Anal. 2015, 115, 315–322. [Google Scholar] [CrossRef]
- Hong, J.; Yang, L.; Zhang, D.; Shi, J. Plant Metabolomics: An Indispensable System Biology Tool for Plant Science. Int. J. Mol. Sci. 2016, 17, 767. [Google Scholar] [CrossRef]
- Amiour, N.; Imbaud, S.; Clément, G.; Agier, N.; Zivy, M.; Valot, B.; Balliau, T.; Armen, P.; Quilleré, I.; Cañas, R.; et al. The use of metabolomics integrated with transcriptomic and proteomic studies for identifying key steps involved in the control of nitrogen metabolism in crops such as maize. J. Exp. Bot. 2012, 63, 5017–5033. [Google Scholar] [CrossRef]
- Sung, J.; Lee, S.; Lee, Y.; Ha, S.; Song, B.; Kim, T.; Waters, B.M.; Krishnan, H.B. Metabolomic profiling from leaves and roots of tomato (Solanumlycopersicum L.) plants grown under nitrogen, phosphorus or potassium-deficient condition. Plant Sci. 2015, 241, 55–64. [Google Scholar] [CrossRef]
- Wang, H.; Huang, R.; Shi, S. Application of non-targeted metabolomics to analyze differential metabolites in willow root systems treated with ammonium nitrogen stress. J. Northeast For. Univ. 2023, 51, 48–52. [Google Scholar] [CrossRef]
- Li, Z.-T.; Zhang, F.-X.; Fan, C.-L.; Ye, M.-N.; Chen, W.-W.; Yao, Z.-H.; Yao, X.-S.; Dai, Y. Discovery of potential Q-marker of traditional Chinese medicine based on plant metabolomics and network pharmacology: Periplocae Cortex as an example. Phytomedicine 2021, 85, 153535. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Zhang, S.; Zhang, B.; Yang, K.; Li, X. Interpretation of “guidelines for webbased pharmacologic evaluation methods. Chin. Herb. Med. 2021, 52, 4119–4129. [Google Scholar]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network pharmacology databases for traditional Chinese medicine: Review and assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Fotis, C.; Antoranz, A.; Hatziavramidis, D.; Sakellaropoulos, T.; Alexopoulos, L.G. Network-based technologies for early drug discovery. Drug Discov. Today 2018, 23, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, S.; Niu, S.; Ma, X.; Li, H.; Jing, M.; Zhao, Y. Network pharmacology prediction and molecular docking-based strategy to explore the potential mechanism of Huanglian Jiedu Decoction against sepsis. Comput. Biol. Med. 2022, 144, 105389. [Google Scholar] [CrossRef]
- Manai-Djebali, H.; Taamalli, A.; Iswaldi, I.; Arráez-Román, D.; Segura-Carretero, A.; Marouani, A.; Mliki, A.; Martínez-Cañas, M.A.; Ghorbel, A. Chemometric analysis of Tunisian durum wheat metabolites using UPLC-ESI-QTOF-MS/MS. J. Food Sci. 2023, 88, 2439–2462. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, R.; Song, Y.; He, J.; Sun, J.; Bai, J.; An, Z.; Dong, L.; Zhan, Q.; Abliz, Z. RRLC-MS/MS -based metabonomics combined with in -depth analysis of metabolic correlation network: Finding potential biomarkers for breast cancer. Analyst 2009, 134, 2003–2011. [Google Scholar] [CrossRef]
- Lu, L.-N.; Liu, R.; Zhou, D.-W. Application of O-PLS in fundamental study of non-invasive measurement of human blood glucose concentration with near infrared spectroscopy. Guang Pu Xue Yu Guang Pu Fen Xi 2005, 25, 1950–1954. [Google Scholar]
- Liu, J.; Liu, J.; Tong, X.; Peng, W.; Wei, S.; Sun, T.; Wang, Y.; Zhang, B.; Li, W. Network Pharmacology Prediction and Molecular Docking-Based Strategy to Discover the Potential Pharmacological Mechanism of Huai Hua San Against Ulcerative Colitis. Drug Des. Dev. Ther. 2021, ume 15, 3255–3276. [Google Scholar] [CrossRef]
- Yang, M.; Li, Y.; Feng, C.; Zhang, X.; Liu, S.; Lin, J. Environmental behavior of steroid hormones and their influencing factors. J. Agric. Environ. Sci. 2012, 31, 849–856. [Google Scholar]
- Chang, X.; Wang, H.; Yang, Z.; Wang, Y.; Li, J.; Han, Z. ESR2 polymorphisms on prostate cancer risk: A systematic review and meta-analysis. Medicine 2023, 102, e33937. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Li, P.L.; Hua, Y.L.; Ji, P.; Yao, W.L.; Zhang, X.S.; Zhong, L.J.; Wei, Y.M. Effects of Tao Hong Si Wu Tang on Apoptosis and EMT through JAK2/STAT3 Signaling Pathway in Rats with Pulmonary Fibrosis Model. Chin. Bull. Pharmacol. 2023, 7, 1577–1583. Available online: http://kns.cnki.net/kcms/detail/34.1086.R.20230724.1344.050.html (accessed on 29 July 2023).
- Li, C.; Pan, J.; Xu, C.; Jin, Z.; Chen, X. A Preliminary Inquiry Into the Potential Mechanism of Huang-Lian-Jie-Du Decoction in Treating Rheumatoid Arthritis via Network Pharmacology and Molecular Docking. Front. Cell Dev. Biol. 2021, 9, 740266. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tang, H.; Tang, Q.; Chen, W. Decoding the mechanism of huanglian Jiedu decoction in treating pneumonia based on network pharmacology and molecular docking. Front. Cell Dev. Biol. 2021, 9, 638366. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Hu, J.; Jiang, J.; Xiao, G.; Yang, R.; Li, S.; Li, Y.; Huang, H.; Zhong, H.; Bi, X. Network Pharmacology and Molecular Docking-Based Prediction of the Mechanism of Qianghuo Shengshi Decoction against Rheumatoid Arthritis. BioMed. Res. Int. 2021, 2021, 6623912. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Piñero, J.; Saüch, J.; Sanz, F.; Furlong, L.I. The DisGeNET cytoscape app: Exploring and visualizing disease genomics data. Comput. Struct. Biotechnol. J. 2021, 19, 2960–2967. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; A Lempicki, R. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2008, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium. Expansion of the Gene Ontology knowledgebase and resources. Nucleic Acids Res. 2017, 45, D331–D338. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Seukep, A.J.; Guo, M. Recent Advances in Molecular Docking for the Research and Discovery of Potential Marine Drugs. Mar. Drugs 2020, 18, 545. [Google Scholar] [CrossRef] [PubMed]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Ye, M.; Luo, G.; Ye, D.; She, M.; Sun, N.; Lu, Y.-J.; Zheng, J. Network pharmacology, molecular docking integrated surface plasmon resonance technology reveals the mechanism of Toujie Quwen Granules against coronavirus disease 2019 pneumonia. Phytomedicine 2020, 85, 153401. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Identification | tR (min) | Ion Mode | Formula | Measured m/z | Theoretical m/z | MS Fragments | VIP Value | p Value | Structure Types | Source |
|---|---|---|---|---|---|---|---|---|---|---|---|
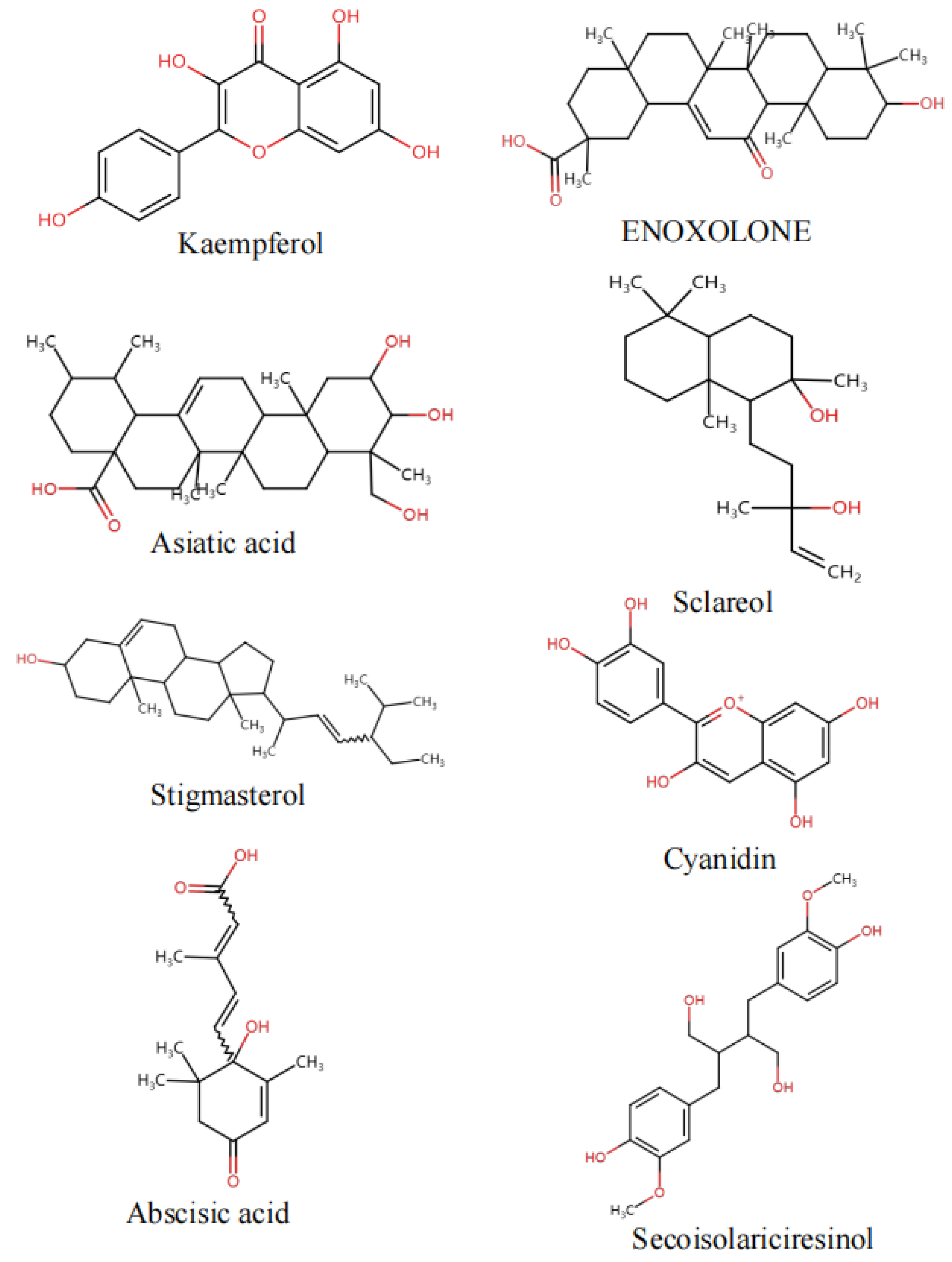
| F1 | Kaempferol | 8.887 | [M + H]+ | C15H10O6 | 287.05579 | 287.05502 | 61.0394, 69.0701, 111.1251, 269.2288, 287.0516, 287.0568 | 1.958 | 0.01738 | Flavonoids | F, M |
| F2 | Asiatic acid | 15.804 | [M + Na]+ | C30H48O5 | 511.32941 | 511.33939 | 300.1110, 349.2677, 449.3287, 511.2344, 511.2828, 511.3082, 511.3290, 511.3867 | 2.0109 | 0.01147 | Triterpenoids | F, M |
| F3 | Cymarin | 14.853 | [M + H]+ | C30H44O9 | 549.30524 | 549.30579 | 550.3086, 551.3120 | 1.8719 | 0.03239 | Others | F, M |
| F4 | Sclareol | 13.925 | [M + H − H2O]+ | C20H36O2 | 291.27084 | 291.2688 | 55.0529, 57.0687, 69.0690, 79.0565, 81.0701, 93.0666, 93.0725, 95.0840, 98.5628, 107.0848, 109.1009, 118.0788, 123.0839, 132.0581, 133.1004, 135.1157, 141.0690, 149.1010, 161.0966, 175.1505, 195.1757, 245.1874, 250.0731, 263.1998, 273.1864, 291.1551, 291.1969, 291.2718 | 1.9011 | 0.0278 | Diterpenoid | F, M |
| F5 | 1,7-Dimethyluric acid | 8.317 | [M + H]+ | C7H8N4O3 | 197.07199 | 197.06691 | 53.0353, 65.0410, 69.0698, 77.0393, 85.0622, 91.0511, 93.0694, 95.0471, 95.0819, 105.0685, 107.0836, 109.0987, 119.0835, 123.1155,128.0610, 131.0839, 133.1003, 135.1167, 137.0957, 140.0525, 151.1129, 155.1074, 161.0964, 171.0120, 179.1072, 197.0722, 197.1181 | 2.0852 | 0.004227 | Steroids | F, M |
| F6 | Stigmasterol | 21.832 | [M + H]+ | C29H48O | 395.36157 | 395.36722 | 396.3649, 397.3682 | 2.0434 | 0.007862 | Steroids | F, M |
| F7 | Enoxolone | 19.489 | [M + H]+ | C30H46O4 | 471.34607 | 471.34689 | 325.0317, 469.7909, 471.0971, 471.3142 | 1.9558 | 0.01902 | Triterpenoids | F, M |
| F8 | Ouabain | 8.012 | [M + H]+ | C29H60O20 | 585.29474 | 585.29102 | 585.2968 | 1.8457 | 0.03718 | Steroids | F, M |
| F9 | Cyanidin | 7.823 | [M]+ | C15H11O6 | 287.04913 | 287.05447 | 66.9806, 69.0682, 89.0590, 105.0666, 111.1155, 202.0814, 283.0601, 287.0614 | 1.7815 | 0.04971 | Others | F, M |
| F10 | Pelargonidin 3-O-rutinoside | 6.722 | [M]+ | [C27H31O14]+ | 579.17236 | 579.17078 | 549.1657, 561.1414, 579.1728 | 1.9023 | 0.02756 | Others | F, M |
| F11 | Abscisic acid | 9.078 | [M − H]− | C15H20O4 | 263.1319 | 263.1283 | 136.0574, 137.0663, 143.0734, 148.0572, 153.091, 161.1010, 163.0785, 189.1001, 201.1309, 204.1161 | 1.9278 | 0.01339 | Organic acid | F, M |
| F12 | Piceid | 3.831 | [M − H]− | C20H22O8 | 389.12888 | 389.12271 | 59.0125, 65.3812, 85.0703, 89.0241, 101.0230, 117.0439, 121.0454, 123.0454, 128.0361, 134.0366, 135.0439, 138.0333, 149.0596, 150.0318, 151.0397, 153.0543, 154.0292, 158.0407, 165.0587, 171.0766, 177.0163, 178.0279, 181.0443, 191.0726, 193.0511, 209.1208, 227.1399, 267.0635, 279.0677, 282.0882, 294.0857, 297.1192, 303.1496, 312.0914, 312.1022, 327.1270, 345.1371, 389.1862 | 1.7359 | 0.04879 | Polyphenols | F, M |
| F13 | Secoisolariciresinol | 8.654 | [M − H]− | C20H26O6 | 407.17181 | 407.17114 | 57.0342, 73.0289, 80.9415, 85.0344, 89.0221, 127.9367, 145.0341, 145.0685, 149.1069, 161.0283, 163.0796, 165.0953, 175.1069, 188.9399, 191.0908, 191.0922, 194.2182, 197.0443, 209.0041, 227.1967, 234.0806, 245.1406, 245.1470, 253.0777, 267.0733, 269.1503, 271.2068, 283.1877, 284.0556, 287.1893, 289.0115, 303.2969, 321.1612, 371.2426, 396.0325, 407.0913, 407.1902, 407.2478, 407.2561, 407.2623, 407.2643, 407.2705, 407.2767, 407.2993 | 1.7599 | 0.04208 | Terpenoids | F, M |
| F14 | Cucurbitacin B | 11.48 | [M − H]− | C32H46O8 | 557.30316 | 557.31201 | 558.3065, 559.3098 | 1.909 | 0.01928 | Terpenoids | F, M |
| F15 | Lauric acid | 20.208 | [M − H]− | C12H24O2 | 199.17165 | 199.17062 | 68.9935, 162.8397, 169.9996, 181.9982, 199.1714 | 1.8381 | 0.02488 | Others | F, M |
| F16 | Geraniol | 17.201 | [M + H − H2O]+ | C10H18O | 137.13045 | 137.133 | 55.0128, 55.0522, 57.0310, 67.0575, 69.0691, 79.0547, 81.0316, 81.0435, 81.0711, 91.0531, 93.0696, 93.0745, 94.06378, 95.0572, 95.0632, 95.0891, 107.0753, 108.5005, 137.0217, 137.0982, 137.1090 | 1.7995 | 0.04855 | Monoterpenes | F, M |
| F17 | Goyazensolide | 6.149 | [M − H]− | C19H20O7 | 359.1062 | 359.11362 | 360.1095, 361.1129 | 1.8243 | 0.03397 | Terpenoids | F, M |
| Target | Target (PDB ID) | Structure | Affinity (kcal/mol) |
|---|---|---|---|
| MAP2K1 | 5HZE |  | −13.2 |
| ESR1 | 1A52 |  | −8.5 |
| ESR2 | 1L2J |  | −7.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, S.; Tian, Q.; Hui, M.; Zhang, S. Revealing the Antiperspirant Components of Floating Wheat and Their Mechanisms of Action through Metabolomics and Network Pharmacology. Molecules 2024, 29, 553. https://doi.org/10.3390/molecules29030553
Dong S, Tian Q, Hui M, Zhang S. Revealing the Antiperspirant Components of Floating Wheat and Their Mechanisms of Action through Metabolomics and Network Pharmacology. Molecules. 2024; 29(3):553. https://doi.org/10.3390/molecules29030553
Chicago/Turabian StyleDong, Shengnan, Qing Tian, Ming Hui, and Shouyu Zhang. 2024. "Revealing the Antiperspirant Components of Floating Wheat and Their Mechanisms of Action through Metabolomics and Network Pharmacology" Molecules 29, no. 3: 553. https://doi.org/10.3390/molecules29030553
APA StyleDong, S., Tian, Q., Hui, M., & Zhang, S. (2024). Revealing the Antiperspirant Components of Floating Wheat and Their Mechanisms of Action through Metabolomics and Network Pharmacology. Molecules, 29(3), 553. https://doi.org/10.3390/molecules29030553