
Integrating Transcriptome and Chemical Analyses to Provide Insights into Biosynthesis of Terpenoids and Flavonoids in the Medicinal Industrial Crop Andrographis paniculate and Its Antiviral Medicinal Parts
 , , , and
, , , and
Abstract
:
1. Introduction
2. Results
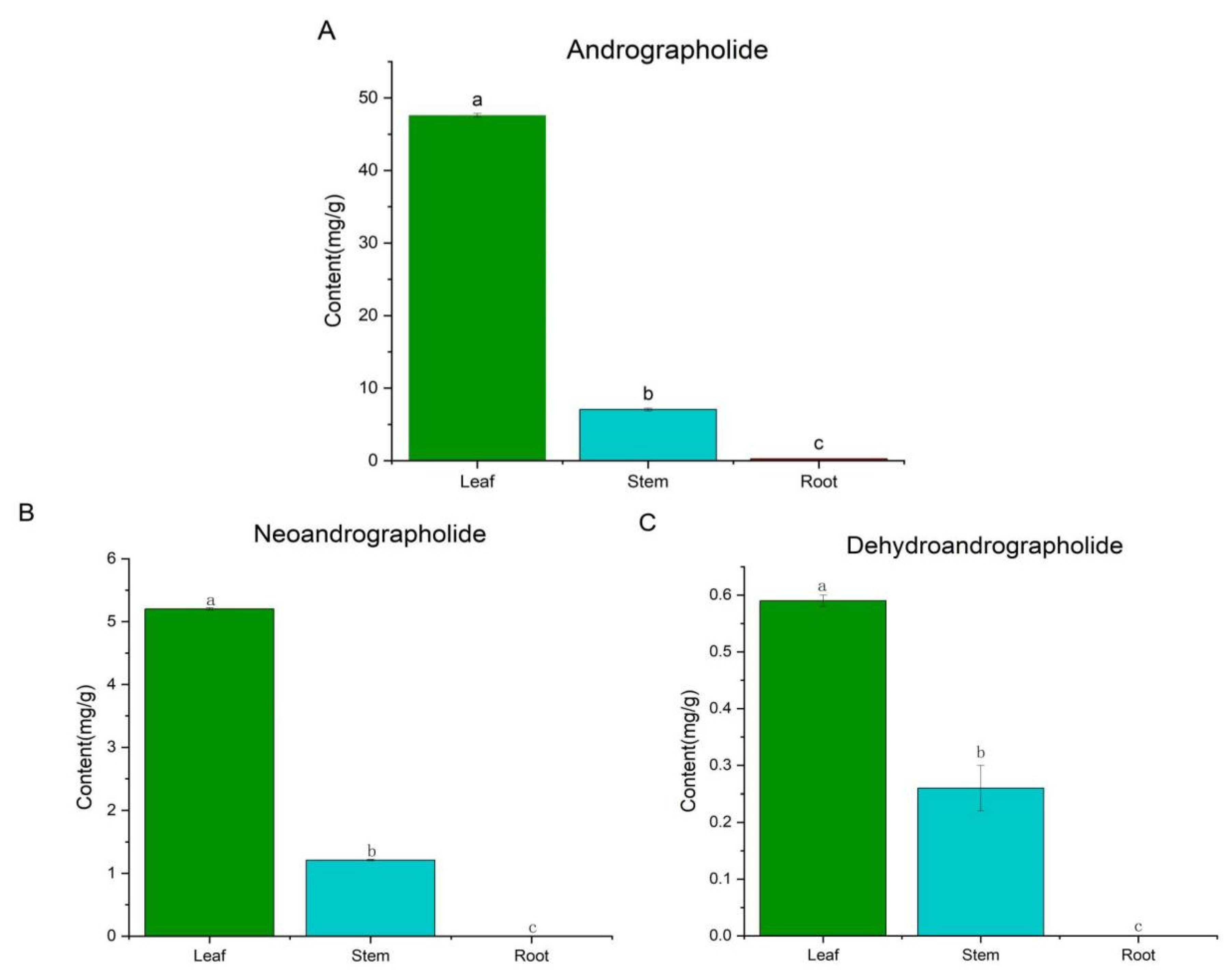
2.1. UPLC-MS
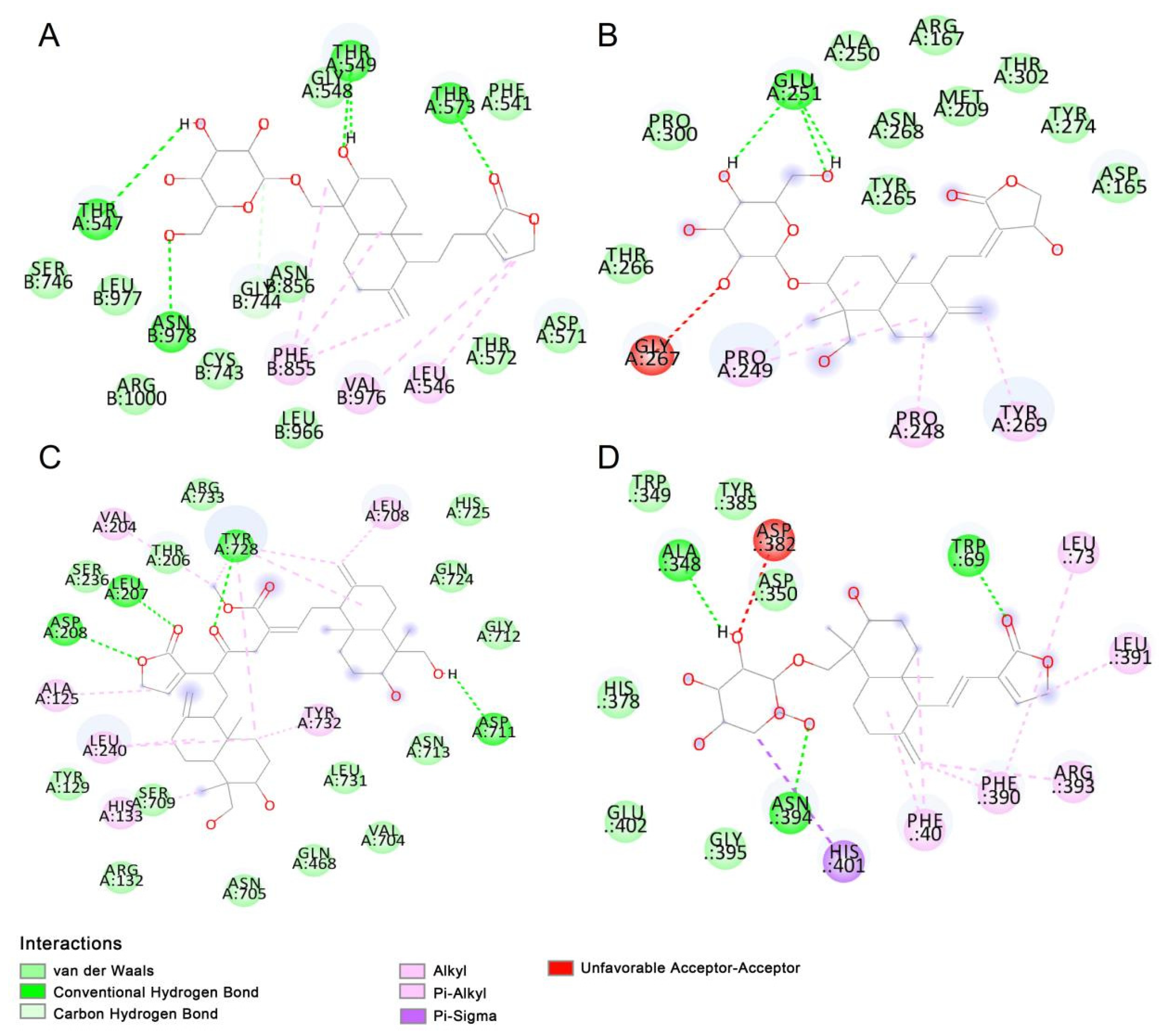
2.2. Molecular Docking
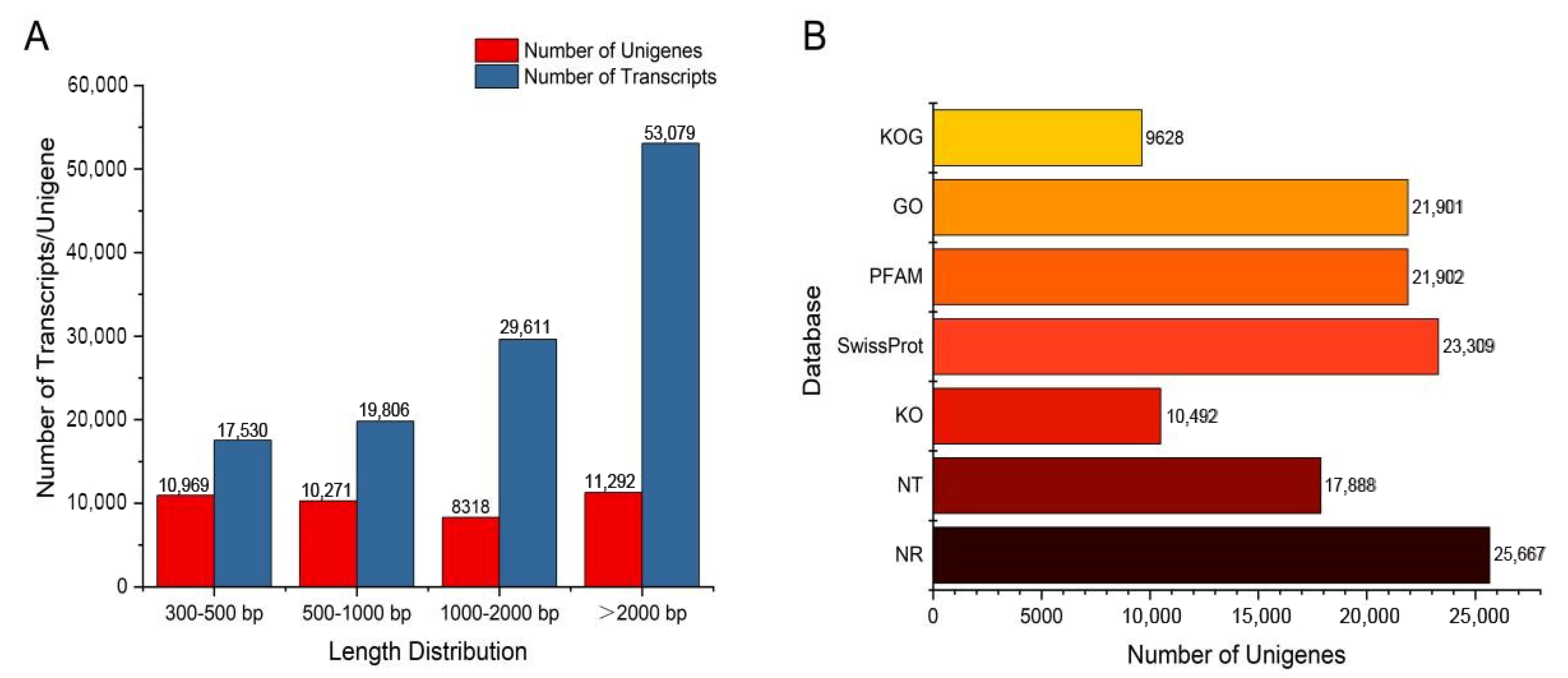
2.3. Transcriptome Sequencing and De Novo Assembly of A. paniculata
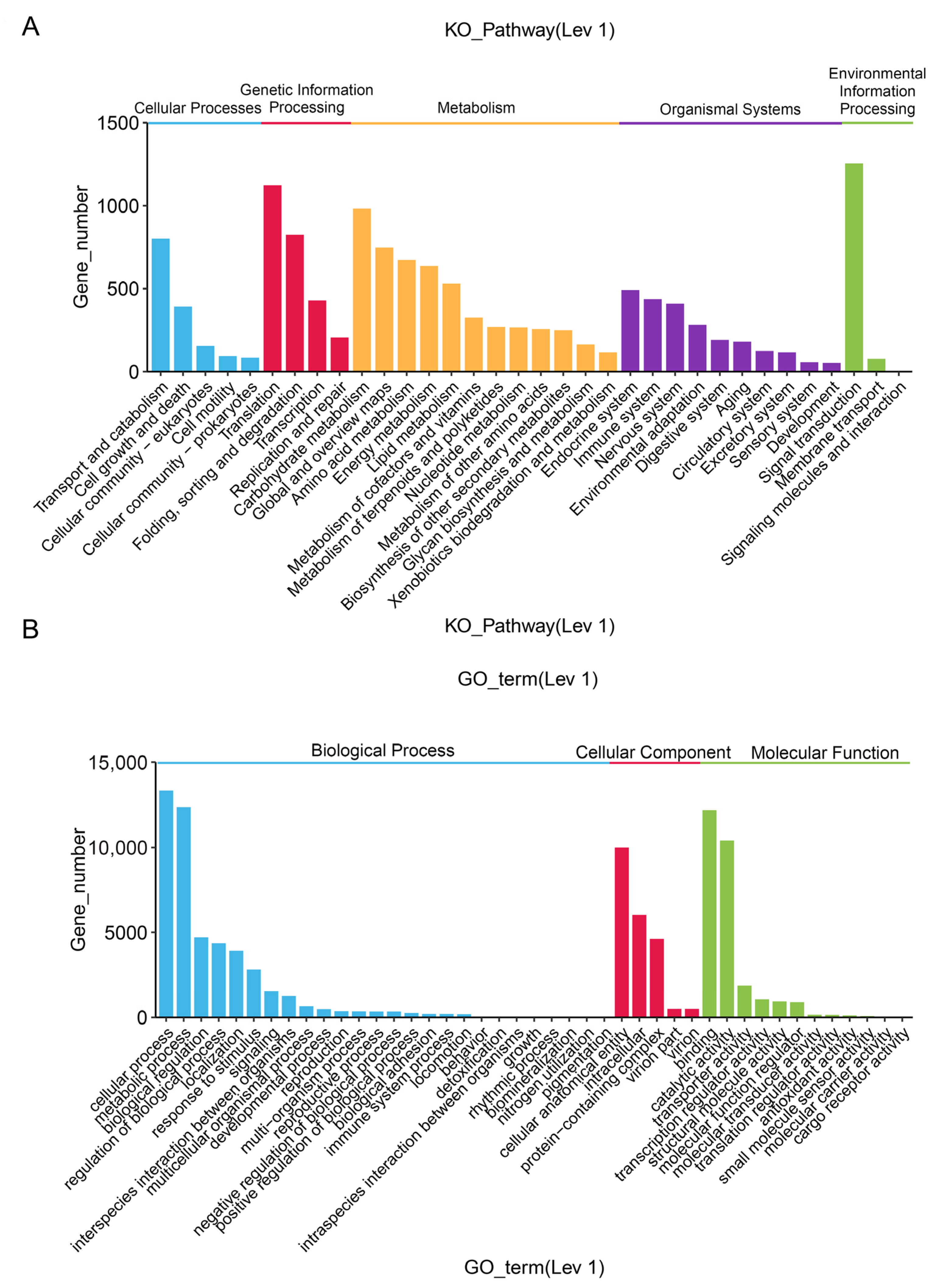
2.4. Functional Annotation of the Unigenes
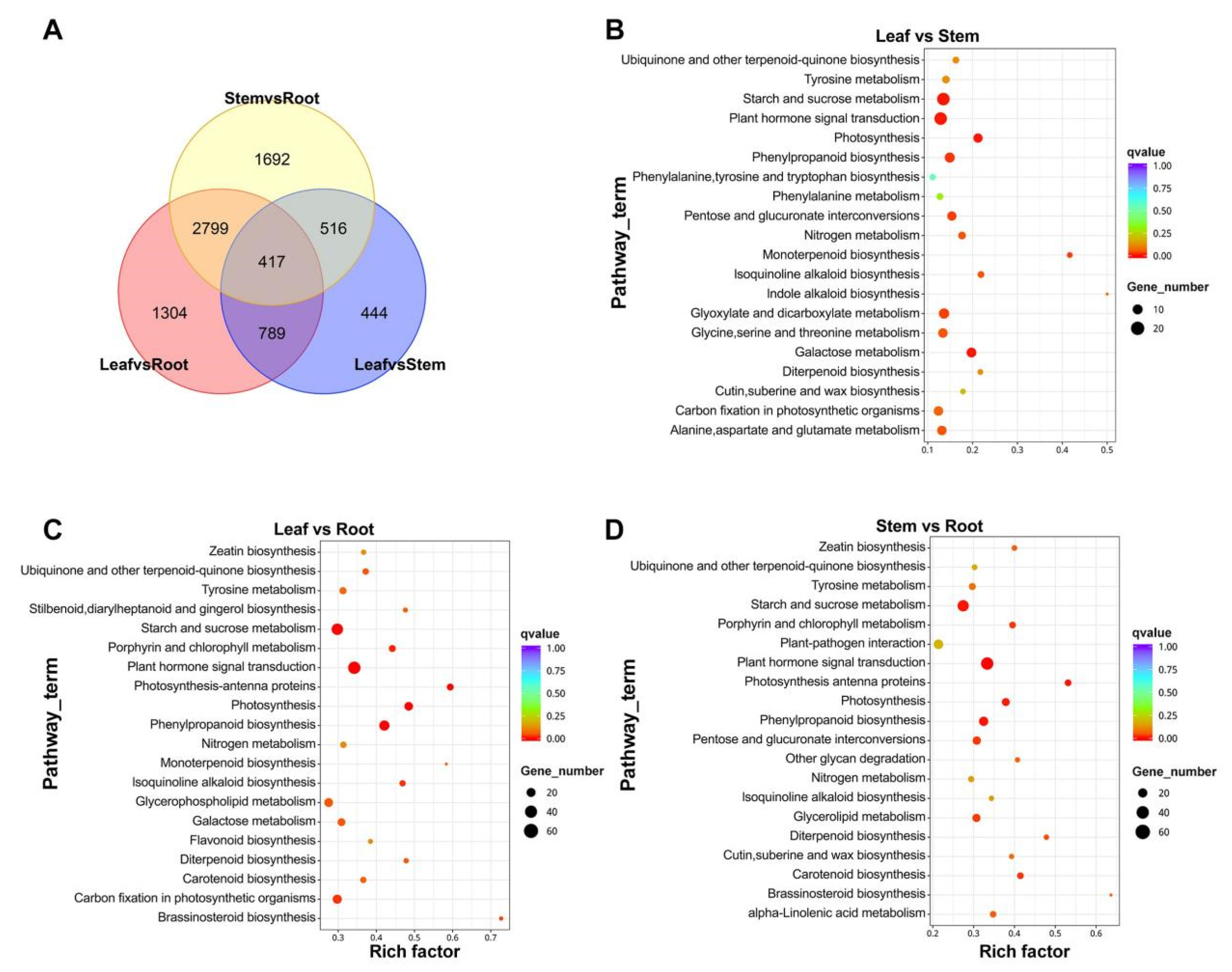
2.5. Differential Expression Analysis of Unigenes
2.6. Transcriptome Analysis of Active Ingredients
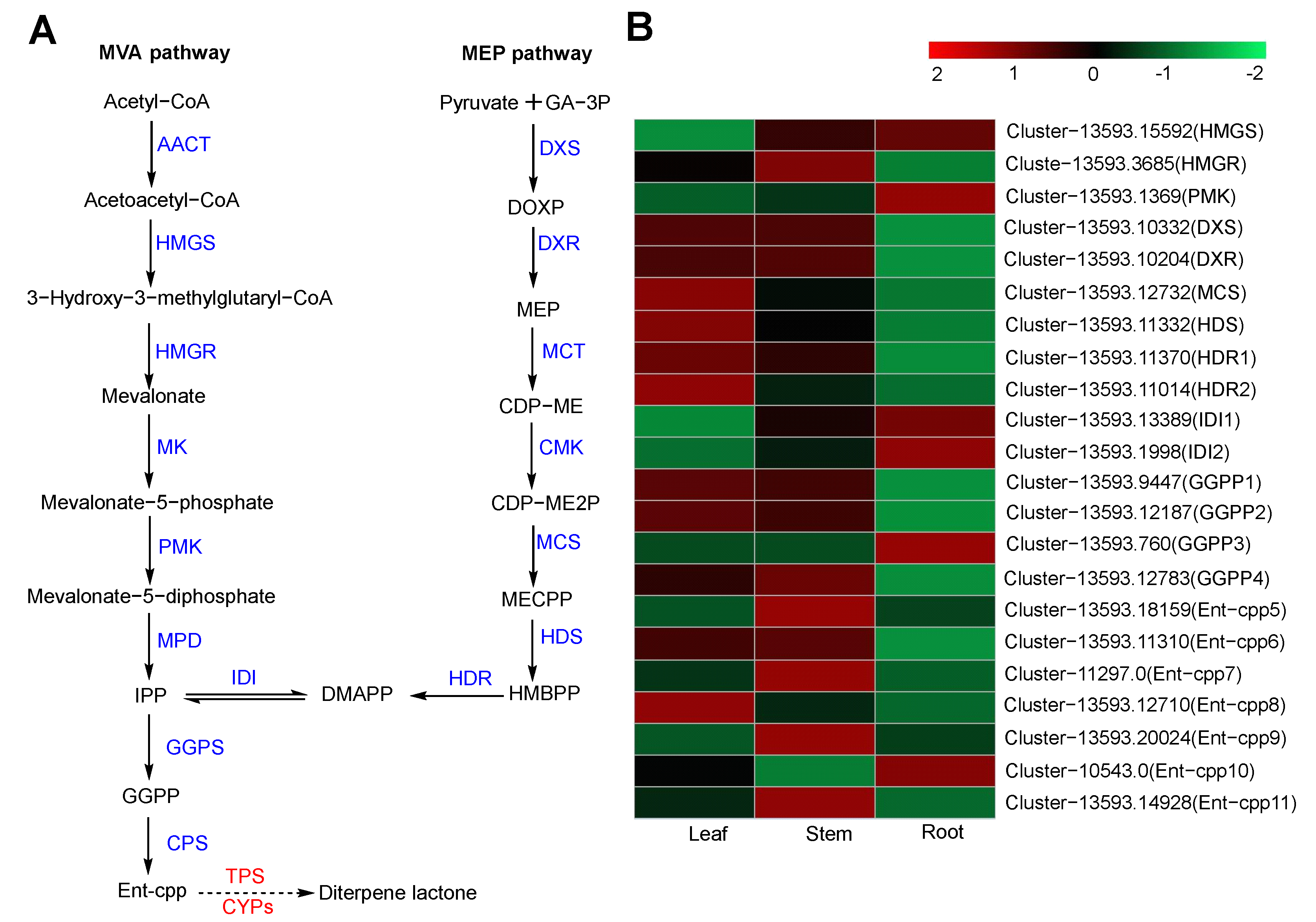
2.6.1. Diterpene Biosynthesis
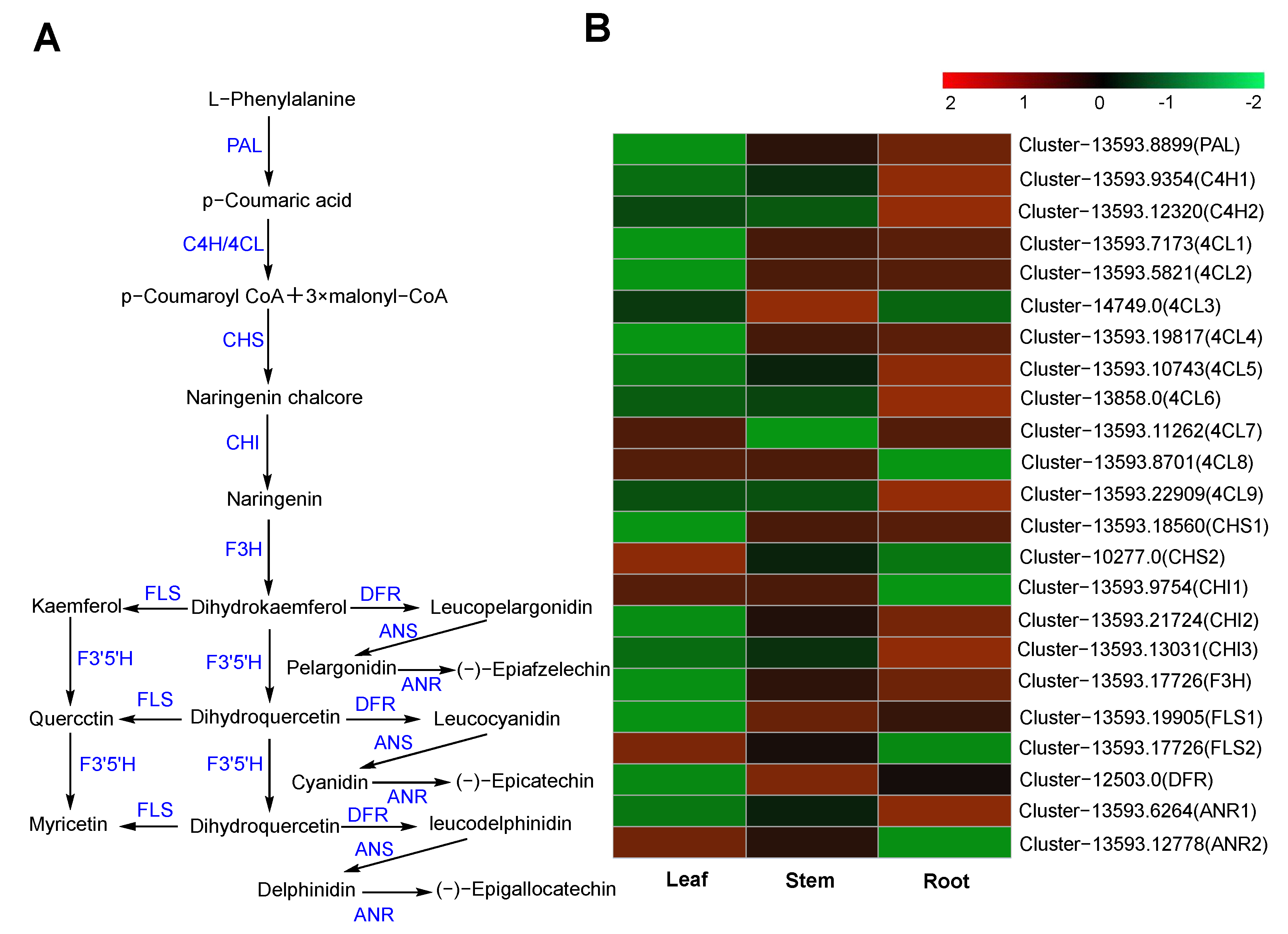
2.6.2. Flavonoid Biosynthesis
2.7. Identification of SSRs
2.8. Transcription Factor Analysis
2.9. qRT-PCR Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. UPLC-MS
4.3. Molecule Docking
4.4. Analysis of Different Tissue Components
4.5. Transcriptome Sequencing and Functional Annotation
4.6. Different Expression Analysis
4.7. Simple Sequence Repeat (SSR) Prediction
4.8. Quantitative Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hossain, S.; Urbi, Z.; Karuniawati, H.; Mohiuddin, R.B.; Moh Qrimida, A.; Allzrag, A.M.M.; Ming, L.C.; Pagano, E.; Capasso, R. Andrographis paniculata (Burm. f.) Wall. ex Nees: An Updated Review of Phytochemistry, Antimicrobial Pharmacology, and Clinical Safety and Efficacy. Life 2021, 11, 348. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.C.; Chan, T.K.; Ng, D.S.; Sagineedu, S.R.; Stanslas, J.; Wong, W.S. Andrographolide and its analogues: Versatile bioactive molecules for combating inflammation and cancer. Clin. Exp. Pharmacol. Physiol. 2012, 39, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.; Dar, L.; Kaushik, S.; Yadav, J.P. Identification and characterization of new potent inhibitors of dengue virus NS5 proteinase from Andrographis paniculata supercritical extracts on in animal cell culture and in silico approaches. J. Ethnopharmacol. 2021, 267, 113541. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Sun, M.; Mei, Y.; Gu, Y.; Xu, S.; Li, J.; Cai, S.; Wang, J. Research progress in the medicinal plant Andrographis paniculate. Guangdong Agric. Sci. 2021, 48, 9–16. [Google Scholar] [CrossRef]
- Jayakumar, T.; Hsieh, C.Y.; Lee, J.J.; Sheu, J.R. Experimental and Clinical Pharmacology of Andrographis paniculata and Its Major Bioactive Phytoconstituent Andrographolide. Evid. Based Complement. Alternat. Med. 2013, 2013, 846740. [Google Scholar] [CrossRef] [PubMed]
- Sudhakaran, M.V. Botanical pharmacognosy of Andrographis paniculata (Burm. F.) Wall. Ex. Nees. Pharmacogn. J. 2012, 4, 1–10. [Google Scholar] [CrossRef]
- Arsakhant, P.; Sirion, U.; Chairoungdua, A.; Suksen, K.; Piyachaturawat, P.; Suksamrarn, A.; Saeeng, R. Design and synthesis of C-12 dithiocarbamate andrographolide analogues as an anticancer agent. Bioorg. Med. Chem. Lett. 2020, 30, 127263. [Google Scholar] [CrossRef]
- Li, L.; Yue, G.G.; Lee, J.K.; Wong, E.C.; Fung, K.P.; Yu, J.; Lau, C.B.; Chiu, P.W. Gene expression profiling reveals the plausible mechanisms underlying the antitumor and antimetastasis effects of Andrographis paniculata in esophageal cancer. Phytother. Res. 2018, 32, 1388–1396. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, L.; Wu, H.; Xu, M.; Guo, R.; Yu, X.; Lu, Y.; Luo, H.; Liu, D.; Wang, Z.; et al. Development of modern research on Andrographis paniculate. Chin. J. Exp. Tradit. Med. Form. 2018, 24, 222–234. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef]
- Cai, N.; Li, Y.; Zhou, G.R.; Xin, C.; Xie, J.; Zhou, H.H.; Miao, X.; Zhou, S.P.; He, Y.; Wang, C. Theoretical basis and effect characteristics of andrographolide against COVID-19. Tradit. Herbal Drugs 2020, 51, 1159–1166. [Google Scholar] [CrossRef]
- Chen, D.L.; Zhong, C.; Lin, Y. Advances in germplasm resources, breeding and cultivation of medicinal plant Andrographis paniculata. J. Jiangsu Agric. Sci. 2020, 48, 34–40. [Google Scholar] [CrossRef]
- Hunter, W.N. The non-mevalonate pathway of isoprenoid precursor biosynthesis. J. Biol. Chem. 2007, 282, 21573–21577. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Liu, Q.; Li, C.; Fu, Y.; Wang, Q. Functional Characterization of ApCPS Involved in Andrographolides Biosynthesis by Virus-induced Gene Silencing. Acta Bot. Boreal.-Occident. Sin. 2016, 36, 17–22. [Google Scholar] [CrossRef]
- Sun, W.; Leng, L.; Yin, Q.; Xu, M.; Huang, M.; Xu, Z.; Zhang, Y.; Yao, H.; Wang, C.; Xiong, C.; et al. The genome of the medicinal plant Andrographis paniculata provides insight into the biosynthesis of the bioactive diterpenoid neoandrographolide. Plant J. 2019, 97, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lin, H.X.; Su, P.; Chen, T.; Guo, J.; Gao, W.; Huang, L.Q. Molecular cloning and functional characterization of multiple geranylgeranyl pyrophosphate synthases (ApGGPPS) from Andrographis paniculata. Plant Cell Rep. 2019, 38, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Agrawal, L.; Misra, R.C.; Sharma, S.; Ghosh, S. Andrographis paniculata transcriptome provides molecular insights into tissue-specific accumulation of medicinal diterpenes. BMC Genom. 2015, 16, 659. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, S.; Kumar, H.D.V.; Nandan, M.; Mantesh, M.; Shankarappa, K.S.; Venkataravanappa, V.; Basha, C.R.J.; Reddy, C.N.L. In silico docking analysis revealed the potential of phytochemicals present in Phyllanthus amarus and Andrographis paniculata, used in Ayurveda medicine in inhibiting SARS-CoV-2. 3 Biotech 2021, 11, 44. [Google Scholar] [CrossRef]
- Rajagopal, K.; Varakumar, P.; Baliwada, A.; Byran, G. Activity of phytochemical constituents of Curcuma longa (turmeric) and Andrographis paniculata against coronavirus (COVID-19): An in silico approach. Future J. Pharm. Sci. 2020, 6, 104. [Google Scholar] [CrossRef]
- Raghu, D.; Hamill, P.; Banaji, A.; McLaren, A.; Hsu, Y.T. Assessment of the binding interactions of SARS-CoV-2 spike glycoprotein variants. J. Pharm. Anal. 2022, 12, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Aftab, S.O.; Ghouri, M.Z.; Masood, M.U.; Haider, Z.; Khan, Z.; Ahmad, A.; Munawar, N. Analysis of SARS-CoV-2 RNA-dependent RNA polymerase as a potential therapeutic drug target using a computational approach. J. Transl. Med. 2020, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Báez-Santos, Y.M.; Barraza, S.J.; Wilson, M.W.; Agius, M.P.; Mielech, A.M.; Davis, N.M.; Baker, S.C.; Larsen, S.D.; Mesecar, A.D. X-ray structural and biological evaluation of a series of potent and highly selective inhibitors of human coronavirus papain-like proteases. J. Med. Chem. 2014, 57, 2393–2412. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.A. Anti-HCV, nucleotide inhibitors, repurposing against COVID-19. Life Sci. 2020, 248, 117477. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-dependent RNA polymerase from COVID-19 virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef]
- Shityakov, S.; Sohajda, T.; Puskás, I.; Roewer, N.; Förster, C.; Broscheit, J.A. Ionization states, cellular toxicity and molecular modeling studies of midazolam complexed with trimethyl-beta-cyclodextrin. Molecules 2014, 19, 16861–16876. [Google Scholar] [CrossRef]
- Srivastava, N.; Akhila, A. Biosynthesis of andrographolide in Andrographis paniculata. Phytochemistry 2010, 71, 1298–1304. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, L.; Wu, B.; Li, X.; Chen, S.; Lu, S. Genome-wide identification and characterization of novel genes involved in terpenoid biosynthesis in Salvia miltiorrhiza. J. Exp. Bot. 2012, 63, 2809–2823. [Google Scholar] [CrossRef]
- Misra, R.C.; Garg, A.; Roy, S.; Chanotiya, C.S.; Vasudev, P.G.; Ghosh, S. Involvement of an ent-copalyl diphosphate synthase in tissue-specific accumulation of specialized diterpenes in Andrographis paniculata. Plant Sci. 2015, 240, 50–64. [Google Scholar] [CrossRef]
- Kuang, X.; Sun, S.; Wei, J.; Li, Y.; Sun, C. Iso-Seq analysis of the Taxus cuspidata transcriptome reveals the complexity of Taxol biosynthesis. BMC Plant Biol. 2019, 19, 210. [Google Scholar] [CrossRef]
- Tu, L.; Su, P.; Zhang, Z.; Gao, L.; Wang, J.; Hu, T.; Zhou, J.; Zhang, Y.; Zhao, Y.; Liu, Y.; et al. Genome of Tripterygium wilfordii and identification of cytochrome P450 involved in triptolide biosynthesis. Nat. Commun. 2020, 11, 971. [Google Scholar] [CrossRef]
- Su, P.; Tong, Y.; Cheng, Q.; Hu, Y.; Zhang, M.; Yang, J.; Teng, Z.; Gao, W.; Huang, L. Functional characterization of ent-copalyl diphosphate synthase, kaurene synthase and kaurene oxidase in the Salvia miltiorrhiza gibberellin biosynthetic pathway. Sci. Rep. 2016, 6, 23057. [Google Scholar] [CrossRef]
- Kakizaki, T.; Kitashiba, H.; Zou, Z.; Li, F.; Fukino, N.; Ohara, T.; Nishio, T.; Ishida, M. A 2-Oxoglutarate-Dependent Dioxygenase Mediates the Biosynthesis of Glucoraphasatin in Radish. Plant Physiol. 2017, 173, 1583–1593. [Google Scholar] [CrossRef] [PubMed]
- Zi, J.; Mafu, S.; Peters, R.J. To gibberellins and beyond! Surveying the evolution of (di)terpenoid metabolism. Annu. Rev. Plant Biol. 2014, 65, 259–286. [Google Scholar] [CrossRef] [PubMed]
- Cherukupalli, N.; Divate, M.; Mittapelli, S.R.; Khareedu, V.R.; Vudem, D.R. De novo Assembly of Leaf Transcriptome in the Medicinal Plant Andrographis paniculata. Front. Plant Sci. 2016, 7, 1203. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Hamberger, B. P450s controlling metabolic bifurcations in plant terpene specialized metabolism. Phytochem. Rev. 2018, 17, 81–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, F.; Lian, Y.; Xiao, H.; Zheng, J. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.L. Study on Flavonoids from Andrographis paniculate; Guangdong Pharmaceutical University: Guangzhou, China, 2021. [Google Scholar]
- Choudhary, S.; Gaur, R.; Gupta, S. EST-derived genic molecular markers: Development and utilization for generating an advanced transcript map of chickpea. Theor. Appl. Genet. 2012, 124, 1449–1462. [Google Scholar] [CrossRef]
- Deng, C.; Wang, Y.; Huang, F.; Lu, S.; Zhao, L.; Ma, X.; Kai, G. SmMYB2 promotes salvianolic acid biosynthesis in the medicinal herb Salvia miltiorrhiza. J. Integr. Plant Biol. 2020, 62, 1688–1702. [Google Scholar] [CrossRef]
- Hao, X.; Pu, Z.; Cao, G.; You, D.; Zhou, Y.; Deng, C.; Shi, M.; Nile, S.H.; Wang, Y.; Zhou, W.; et al. Tanshinone and salvianolic acid biosynthesis are regulated by SmMYB98 in Salvia miltiorrhiza hairy roots. J. Adv. Res. 2020, 23, 1–12. [Google Scholar] [CrossRef]
- Xie, R.; Lin, Z.; Zhong, C.; Li, S.; Chen, B.; Wu, Y.; Huang, L.; Yao, H.; Shi, P.; Huang, J. Deciphering the potential anti-COVID-19 active ingredients in Andrographis paniculata (Burm. F.) Nees by combination of network pharmacology, molecular docking, and molecular dynamics. RSC. Adv. 2021, 11, 36511–36517. [Google Scholar] [CrossRef]
- Sa-Ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 Activity of Andrographis paniculata extract and its major component andrographolide in human lung epithelial cells and cytotoxicity evaluation in major organ cell representatives. J. Nat. Prod. 2021, 84, 1261–1270. [Google Scholar] [CrossRef]
- Murugan, N.A.; Pandian, C.J.; Jeyakanthan, J. Computational investigation on Andrographis paniculata phytochemicals to evaluate their potency against SARS-CoV-2 in comparison to known antiviral compounds in drug trials. J. Biomol. Struct. Dynam. 2020, 39, 4415–4426. [Google Scholar] [CrossRef] [PubMed]
- Intharuksa, A.; Arunotayanun, W.; Yooin, W.; Sirisa-Ard, P. A Comprehensive Review of Andrographis paniculata (Burm. f.) Nees and its constituents as potential lead compounds for COVID-19 drug discovery. Molecules 2022, 27, 4479. [Google Scholar] [CrossRef]
- Deng, S.; Zhang, Y.; Chen, J.; Wang, J.; Luo, J.; Fang, L.; Liu, D.; Lv, X.; Qian, D.; Zhou, J.; et al. Proteomics and UHPLC-DAD-Q/Orbitrap-MS used to identify impurities in andrographolide. Phytochem. Anal. 2022, 33, 735–745. [Google Scholar] [CrossRef]
- Worakunphanich, W.; Thavorncharoensap, M.; Youngkong, S.; Thadanipon, K.; Thakkinstian, A. Safety of Andrographis paniculata: A systematic review and meta-analysis. Pharmacoep. Drug Saf. 2021, 30, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Liu, J.; Zhang, Y.; Ma, H.; Li, Y.; Gong, Q.; Cao, Y.; Hu, G.; Xie, S.; Fu, S. Dehydroandrographolide inhibits mastitis by activating autophagy without affecting intestinal flora. Aging 2020, 12, 14050–14065. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Zhang, H.L.; Yu, F.F.; Yin, X.R.; Zhao, Y.F.; Ye, F.; Wu, X.Q. Amelioratory effect of neoandrographolide on myocardial ischemic-reperfusion injury by its anti-inflammatory and anti-apoptotic activities. Environ. Toxicol. 2021, 36, 2367–2379. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Jain, M. RNA-Seq for transcriptome analysis in non-model plants. Methods Mol. Biol. 2013, 1069, 43–58. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, S.; Wei, K.; Yang, Z.; Duan, S.; Du, Y.; Qu, P.; Miao, J.; Chen, W.; Dong, Y. Chromosome Level Genome Assembly of Andrographis paniculata. Front. Genet. 2020, 11, 701. [Google Scholar] [CrossRef] [PubMed]
- Hidayat, R.; Wulandari, P. Effects of Andrographis paniculata (Burm. F.) Extract on Diabetic Nephropathy in Rats. Rep. Biochem. Mol. Biol. 2021, 10, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.S.; Chern, H.J.; Damu, A.G.; Kuo, P.C.; Su, C.R.; Lee, E.J.; Teng, C.M. Flavonoids and ent-labdane diterpenoids from Andrographis paniculata and their antiplatelet aggregatory and vasorelaxing effects. J. Asian Nat. Prod. Res. 2008, 10, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Hirai, M.Y.; Yano, M.; Goodenowe, D.B.; Kanaya, S.; Kimura, T.; Awazuhara, M.; Arita, M.; Fujiwara, T.; Saito, K. Integration of transcriptomics and metabolomics for understanding of global responses to nutritional stresses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 10205–10210. [Google Scholar] [CrossRef] [PubMed]
- Pickens, L.B.; Tang, Y.; Chooi, Y.H. Metabolic engineering for the production of natural products. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 211–236. [Google Scholar] [CrossRef] [PubMed]
- Staniek, A.; Bouwmeester, H.; Fraser, P.D.; Kayser, O.; Martens, S.; Tissier, A.; van der Krol, S.; Wessjohann, L.; Warzecha, H. Natural products—Modifying metabolite pathways in plants. Biotechnol. J. 2013, 8, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Songvut, P.; Pholphana, N.; Suriyo, T.; Rangkadilok, N.; Panomvana, D.; Puranajoti, P.; Satayavivad, J. A validated LC-MS/MS method for clinical pharmacokinetics and presumptive phase II metabolic pathways following oral administration of Andrographis paniculata extract. Sci. Rep. 2023, 13, 2534. [Google Scholar] [CrossRef] [PubMed]
- Maurya, V.K.; Kumar, S.; Prasad, A.K.; Bhatt, M.L.B.; Saxena, S.K. Structure-based drug designing for potential antiviral activity of selected natural products from Ayurveda against SARS-CoV-2 spike glycoprotein and its cellular receptor. Virusdisease 2020, 31, 179–193. [Google Scholar] [CrossRef]
- Xia, M.; Guo, Y.; Li, J.; Ma, H.; Gao, W.; Wu, X. Correlation analysis of physicochemical properties with anti-inflammatory activity of Andrographis paniculata (Burm.f.) Nees based on HPLC-DAD, colorimeter and multivariate statistics: A comprehensive quality evaluation strategy. J. Pharm. Biomed. Anal. 2022, 210, 114565. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Jiao, C.; Sun, H.; Rosli, H.G.; Pombo, M.A.; Zhang, P.; Banf, M.; Dai, X.; Martin, G.B.; Giovannoni, J.J.; et al. iTAK: A Program for Genome-wide Prediction and Classification of Plant Transcription Factors, Transcriptional Regulators, and Protein Kinases. Mol. Plant. 2016, 9, 1667–1670. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Compounds | Retention Time | Fomula | Measured Mass (m/z) | MS/MS Fragment |
|---|---|---|---|---|---|
| Diterpene | 12S-Hydroxyandrographolide | 12.09 | C20H32O6 | 369.1532 | 351.2167, 333.2059, 315.1956, 297.1852, 285.1850, 257.1555, |
| Paniculide B | 12.48 | C15H20O5 | 281.1375 | 263.1280, 245.1176, 217.1226 | |
| Dehydroandrographolide | 12.73 | C20H28O4 | 333.2060 | 333.2062, 315.1956, 297.1851, 285.1847, 257.1542 | |
| Andrographolide | 14.03 | C20H30O5 | 351.2169 | 333.2058, 315.1975, 297.1849, 285.1849, 257.1534 | |
| 14-Deoxyandrographiside | 15.49 | C26H40O9 | 497.2746 | 335.2223, 317.2115, 299.2009, 287.2008 | |
| Andrographolactone | 16.35 | C20H24O2 | 297.1852 | 269.1539 | |
| 14-Deoxyandrographolide | 20.53 | C20H30O4 | 335.2216 | 335.2221, 317.2110 299.2006, 287.2007 259.1695 | |
| Bisandrographolide A | 22.23 | C40H56O8 | 665.4039 | 315.1956, 297.1852, 285.1852, 257.1539 | |
| Andrograpanin | 24.16 | C20H30O3 | 319.2269 | 301.2164, 289.2164, 261.1848 | |
| Flavonoid | Andrographidin A | 15.14 | C23H26O10 | 463.1220 | 301.1070, 197.0445 |
| 5, 4’-Dihydroxy-7, 8-dimethoxyflavone | 17.90 | C17H14O6 | 315.0864 | 197.0447, 119.0857 | |
| Dihydroscullcapflavone I | 21.88 | C17H16O6 | 317.1020 | 197.0443, 121.5874 | |
| 5-Hydroxy-7, 8-dimethoxy flavanone | 23.69 | C17H16O5 | 301.1069 | 197.0443, 105.0333 | |
| Apigenin-7,4’-dimethylether | 25.69 | C17H14O5 | 299.0915 | 167.1066, 133.1014, | |
| 5-Hydroxy-7, 2’, 6’-trimethoxy flavone | 26.92 | C18H16O6 | 329.1032 | 167.0705, 163.1516 |
| Group | Compounds | S Protein Binding Affinity (kcal/mol) | PLpro Binding Affinity (kcal/mol) | RdRp Binding Affinity (kcal/mol) | ACE2 Binding Affinity (kcal/mol) |
|---|---|---|---|---|---|
| Positive control | Remdesivir | −8.3 | −6.8 | −8.0 | −7.8 |
| Diterpenes | Andrographolide | −8.0 | −6.1 | −7.7 | −7.6 |
| Andrograpanin | −7.8 | −6.4 | −7.7 | −7.2 | |
| 14-Deoxyandrographiside | −9.0 | −6.8 | −8.2 | −8.2 | |
| 14-Deoxyandrographolide | −8.0 | −5.9 | −7.7 | −7.7 | |
| 14-Deoxy-11-12-didehydroandrographiside | −8.3 | −6.8 | −8.3 | −8.8 | |
| Neoandrographolide | −8.9 | −6.6 | −7.7 | −7.2 | |
| Dehydroandrographolide | −8.1 | −7.4 | −7.2 | −7.0 | |
| Andropanolide | −8.0 | −6.4 | −7.9 | −7.2 | |
| Bisandrographolide A | −9.0 | −5.8 | −7.9 | −8.4 | |
| Bisandrographolide D | −9.0 | −6.6 | −8.9 | −8.0 | |
| Isoandrographolide | −8.2 | −6.8 | −8.0 | −7.6 | |
| 12S-Hydroxyandrographolide | −7.9 | −6.0 | −7.5 | −7.0 | |
| Paniculide B | −7.1 | −6.9 | −6.9 | −6.9 | |
| Andrographolactone | −8.8 | −5.9 | −7.4 | −7.3 | |
| 3-O-β-D-Glucopyranosyl andrographolide | −9.0 | −7.2 | −8.0 | −8.0 | |
| Flavonoids | Andrographidin A | −8.6 | −7.6 | −7.7 | −8.0 |
| Andrographidine E | −8.3 | −6.7 | −7.3 | −7.9 | |
| Andrographidine C | −8.6 | −7.6 | −8.0 | −8.1 | |
| 5, 4’-Dihydroxy-7, 8-dimethoxyflavone | −7.5 | −7.5 | −7.6 | −7.7 | |
| Dihydroscullcapflavone I | −7.6 | −7.0 | −7.2 | −7.1 | |
| 5-Hydroxy-7, 8-dimethoxy flavanone | −8.0 | −7.2 | −7.4 | −7.3 | |
| Apigenin-7, 4’-dimethylether | −7.9 | −7.8 | −7.8 | −7.6 | |
| 5-Hydroxy-7, 2’, 6’-trimethoxy flavone | −7.7 | −6.5 | −7.0 | −7.1 | |
| 1-2-Dihydroxy-6-8-dimethoxy xanthone | −7.5 | −7.0 | −7.1 | −6.8 | |
| Methylswertianin | −7.9 | −6.6 | −7.1 | −6.8 | |
| 5-2’-6’-Trihydroxy-7-methoxyflavone | −8.2 | −6.9 | −7.5 | −7.9 | |
| 5-2’-Dihydroxy-7-8-dimethoxyflavone | −8.2 | −7.0 | −7.7 | −8.2 | |
| 5-7-4’-Trihydroxyflavone | −8.4 | −7.8 | −7.9 | −7.2 | |
| Isoswertisin | −8.4 | −7.4 | −7.7 | −7.4 | |
| Luteolin | −8.5 | −8.3 | −8.4 | −8.0 |
| Pathway Name | KO ID | Input Number | Leaf vs. Stem DEGs | Leaf vs. Root DEGs | Stem vs. Root DEGs |
|---|---|---|---|---|---|
| Phenylalanine metabolism | Ko00360 | 55 | 7 | 16 | 11 |
| Terpenoid backbone biosynthesis | Ko00900 | 93 | 5 | 20 | 20 |
| Diterpene biosynthesis | Ko00904 | 23 | 5 | 11 | 11 |
| Phenylpropanoid biosynthesis | Ko00940 | 114 | 17 | 48 | 37 |
| Flavonoid biosynthesis | Ko00941 | 26 | 2 | 10 | 8 |
| Flavone and flavonol biosynthesis | Ko00944 | 3 | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, K.; Liang, P.; Yu, H.; Liu, H.; Guo, J.; Yan, X.; Li, Z.; Li, G.; Wang, Y.; Wang, C. Integrating Transcriptome and Chemical Analyses to Provide Insights into Biosynthesis of Terpenoids and Flavonoids in the Medicinal Industrial Crop Andrographis paniculate and Its Antiviral Medicinal Parts. Molecules 2024, 29, 852. https://doi.org/10.3390/molecules29040852
Yu K, Liang P, Yu H, Liu H, Guo J, Yan X, Li Z, Li G, Wang Y, Wang C. Integrating Transcriptome and Chemical Analyses to Provide Insights into Biosynthesis of Terpenoids and Flavonoids in the Medicinal Industrial Crop Andrographis paniculate and Its Antiviral Medicinal Parts. Molecules. 2024; 29(4):852. https://doi.org/10.3390/molecules29040852
Chicago/Turabian StyleYu, Kuo, Pengjie Liang, Heshui Yu, Hui Liu, Jialiang Guo, Xiaohui Yan, Zheng Li, Guoqiang Li, Ying Wang, and Chunhua Wang. 2024. "Integrating Transcriptome and Chemical Analyses to Provide Insights into Biosynthesis of Terpenoids and Flavonoids in the Medicinal Industrial Crop Andrographis paniculate and Its Antiviral Medicinal Parts" Molecules 29, no. 4: 852. https://doi.org/10.3390/molecules29040852