
Development of Glycosylation-Modified DPPA-1 Compounds as Innovative PD-1/PD-L1 Blockers: Design, Synthesis, and Biological Evaluation

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
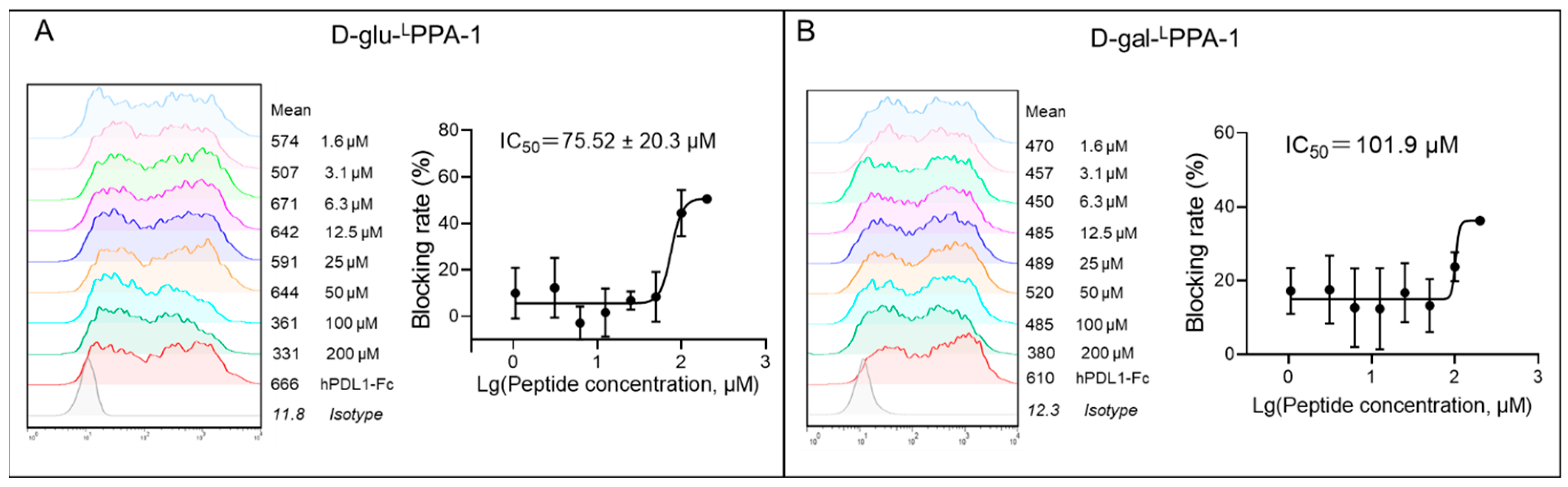
2.1. The Ability of D-glu-LPPA-1 and D-gal-LPPA-1 to Block PD-1/PD-L1 Interaction
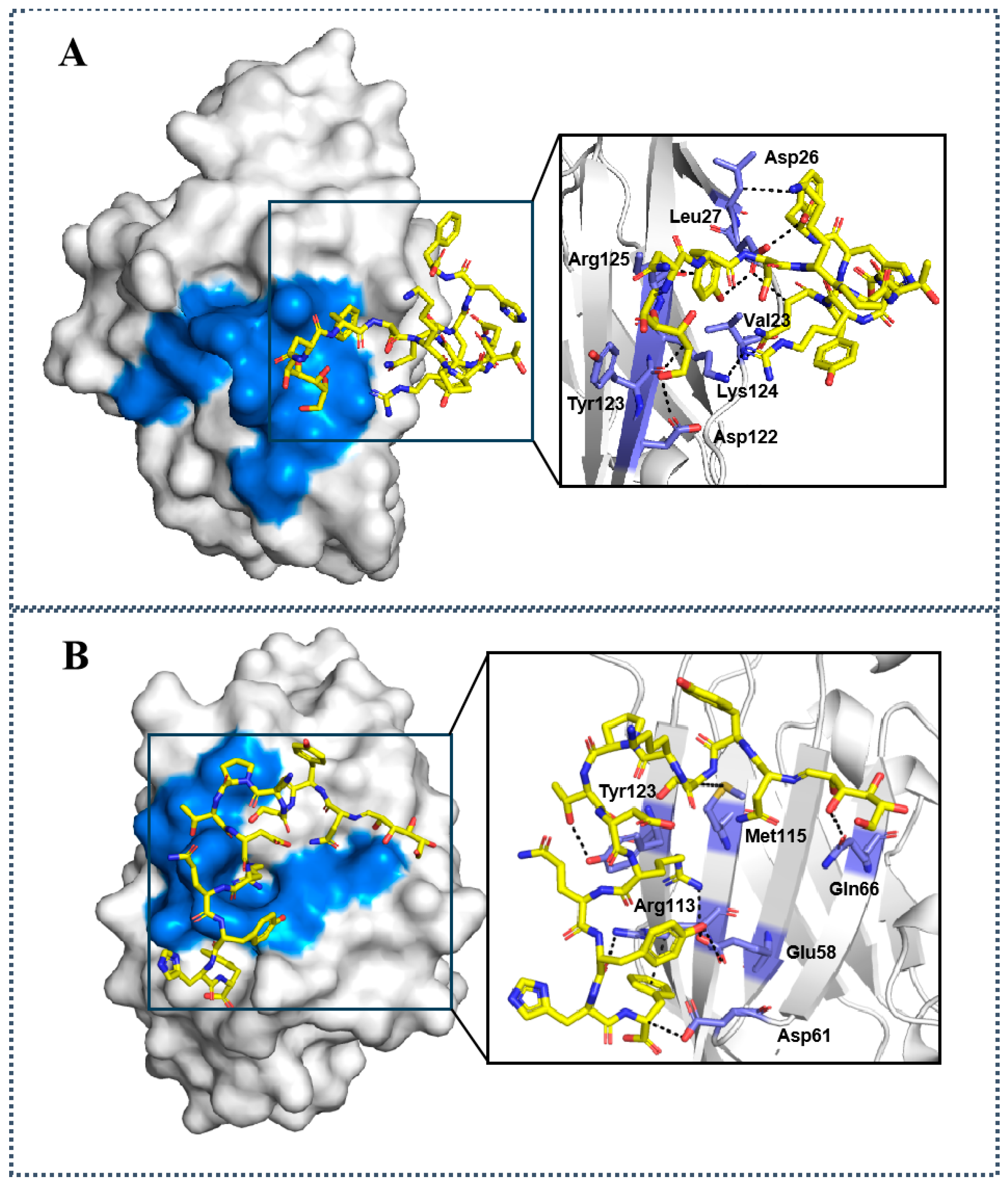
2.2. Molecular Docking Studies of D-glu-LPPA-1 and D-gal-LPPA-1 to PD-1/PD-L1
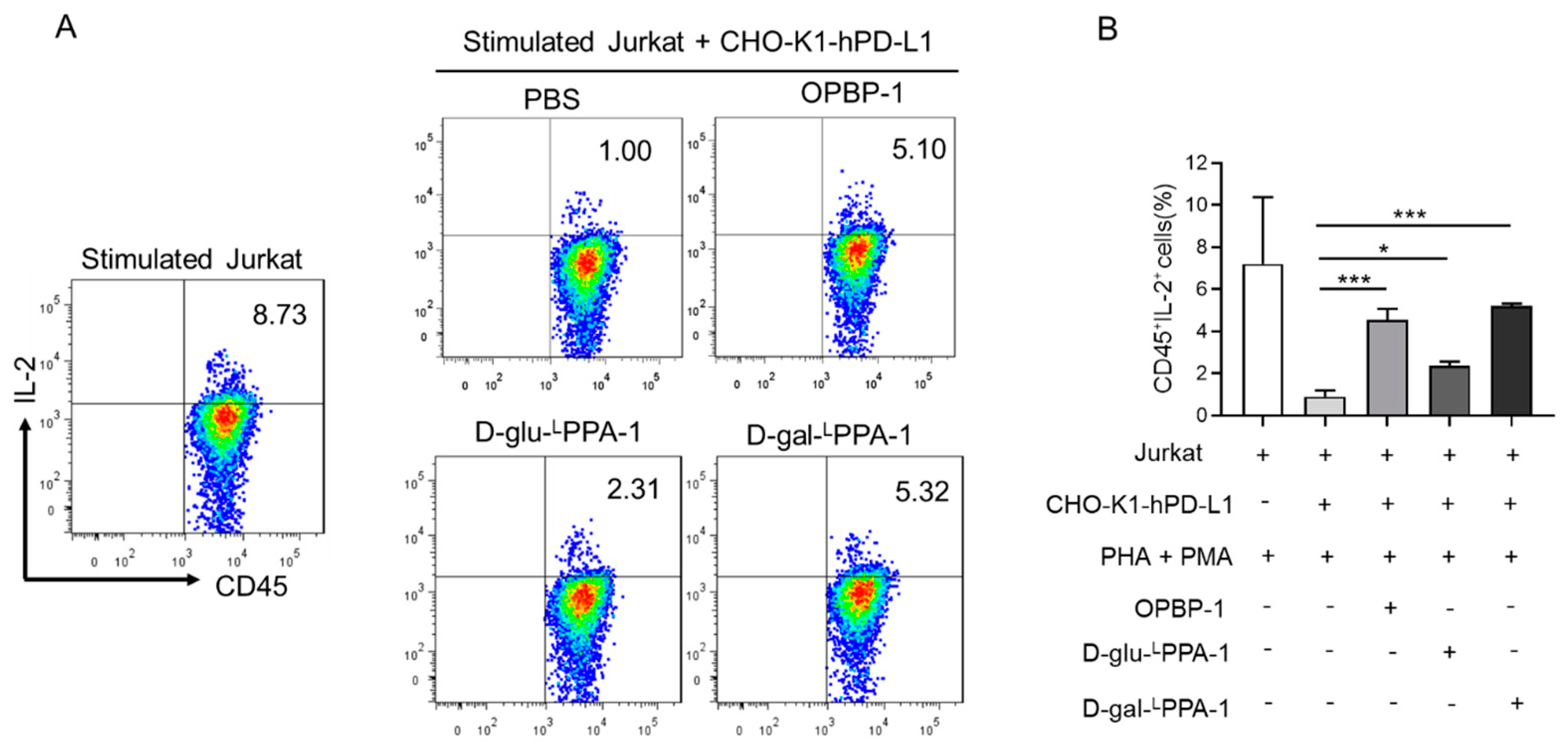
2.3. D-gal-LPPA-1 Enhanced IL-2 Secretion in a Jurkat Cell Coculture Assay
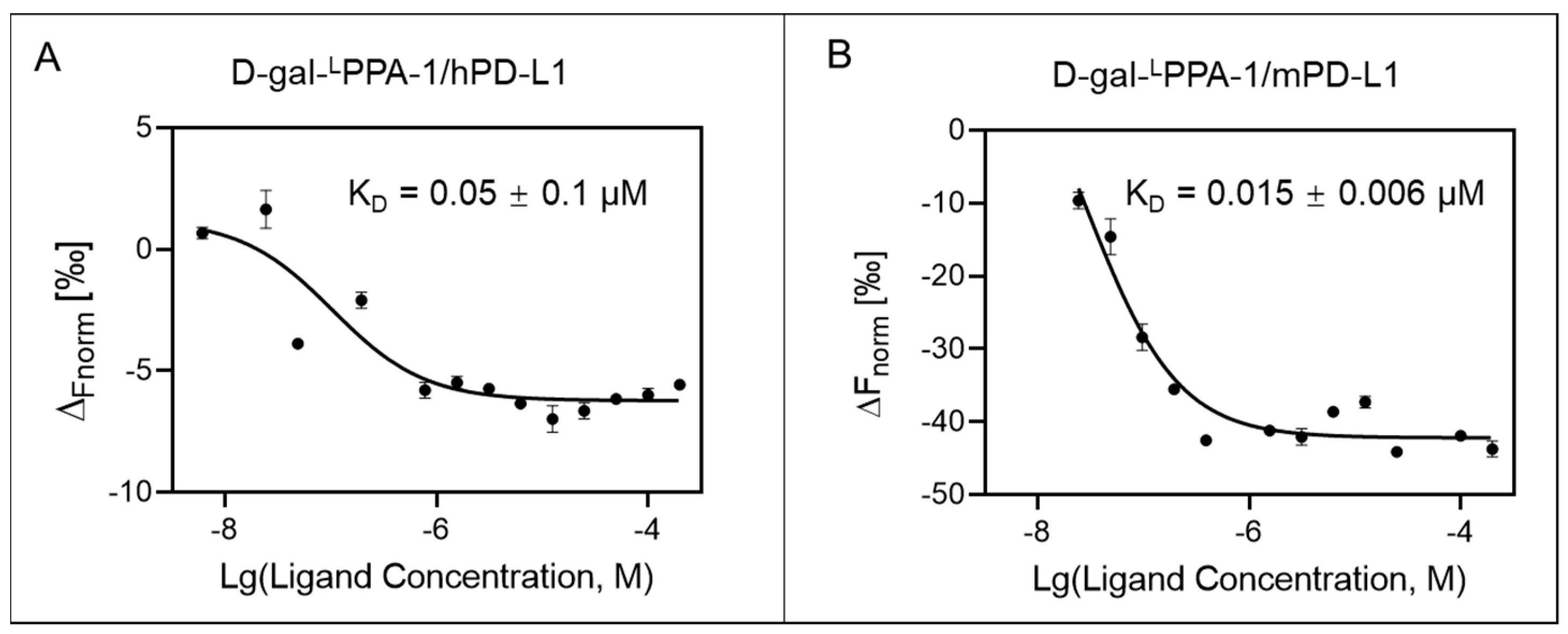
2.4. D-gal-LPPA-1 Exhibited High Binding Affinity for PD-L1
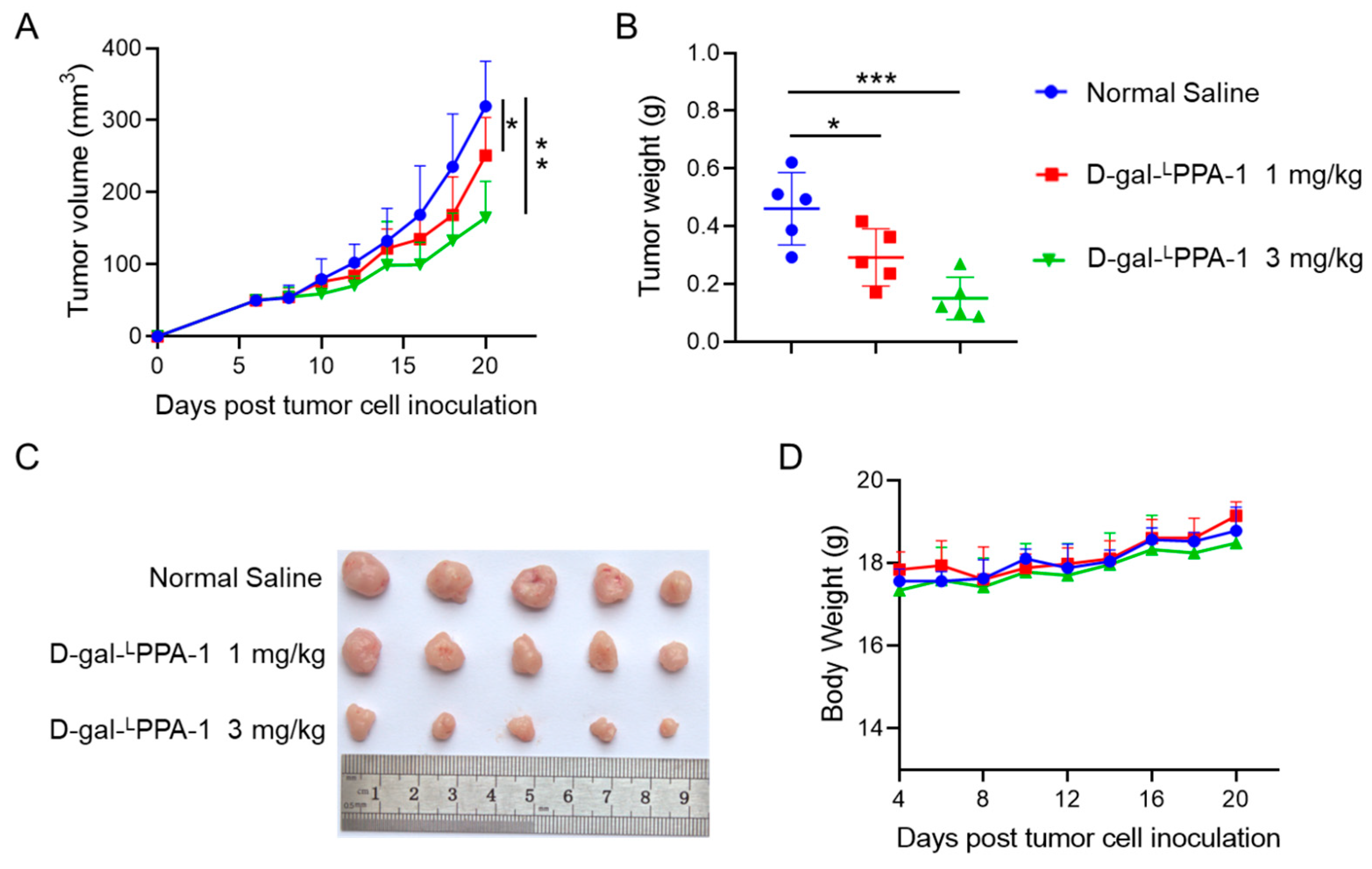
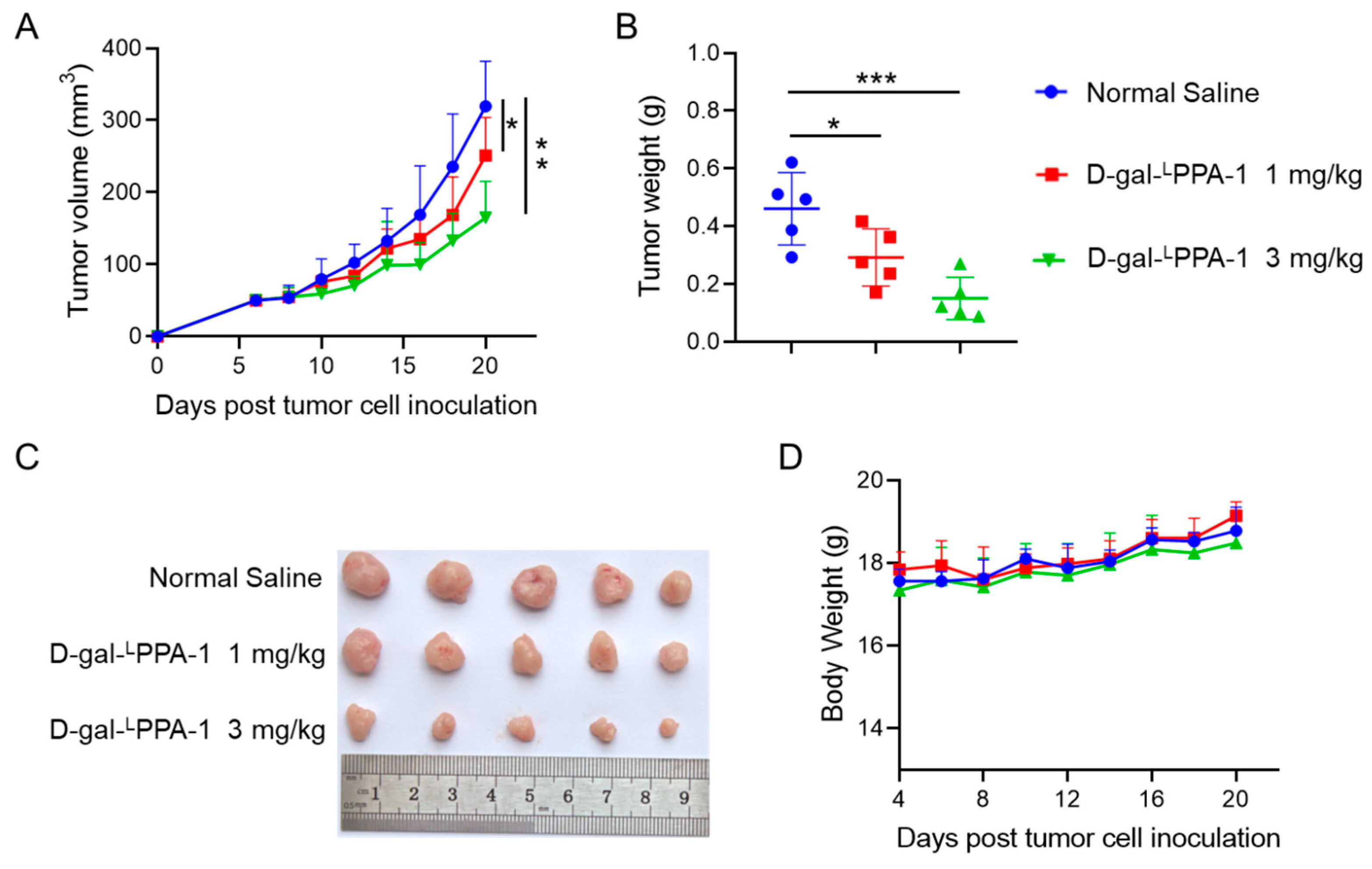
2.5. D-gal-LPPA-1 Significantly Inhibited MC38 Tumor Growth In Vivo
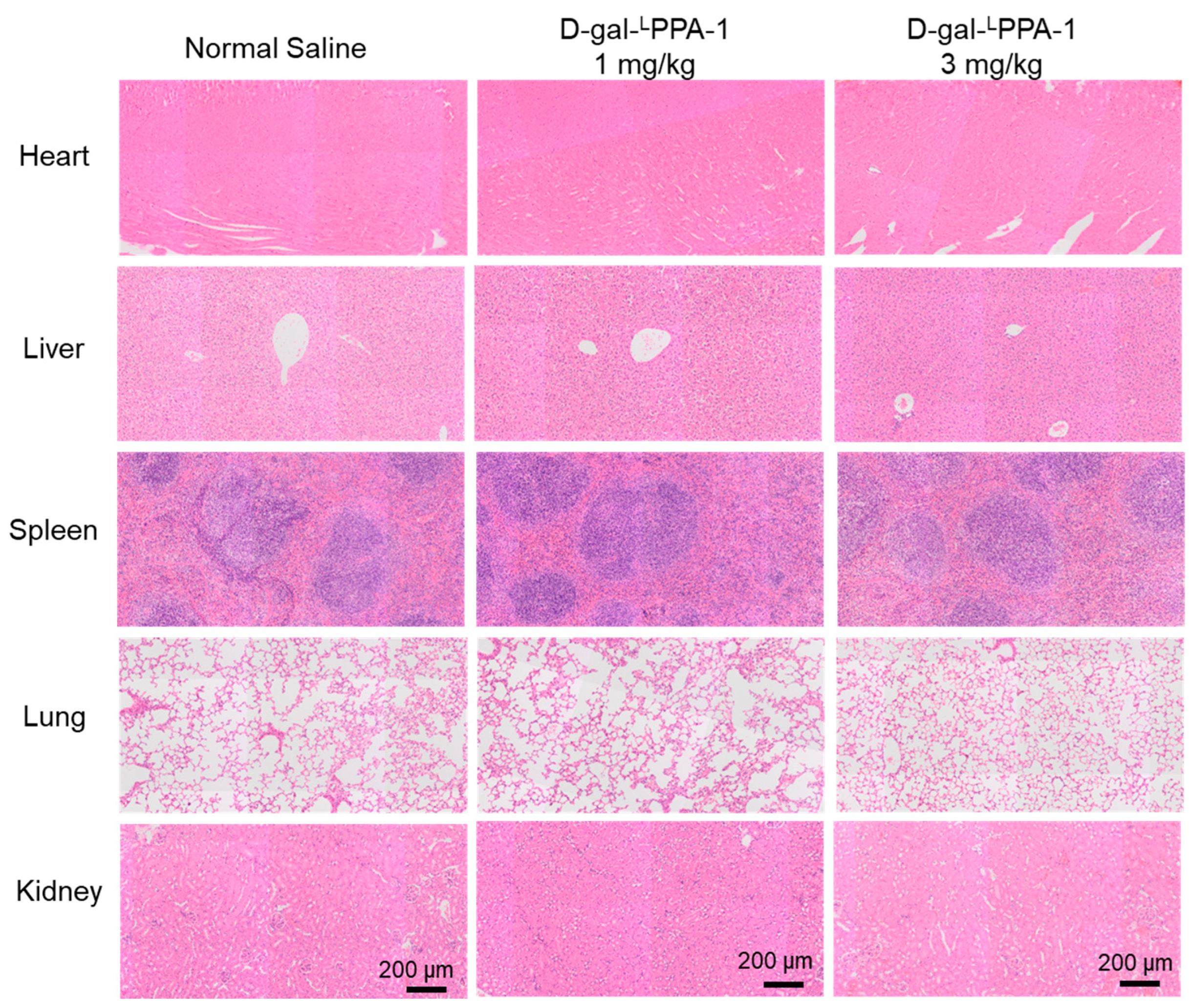
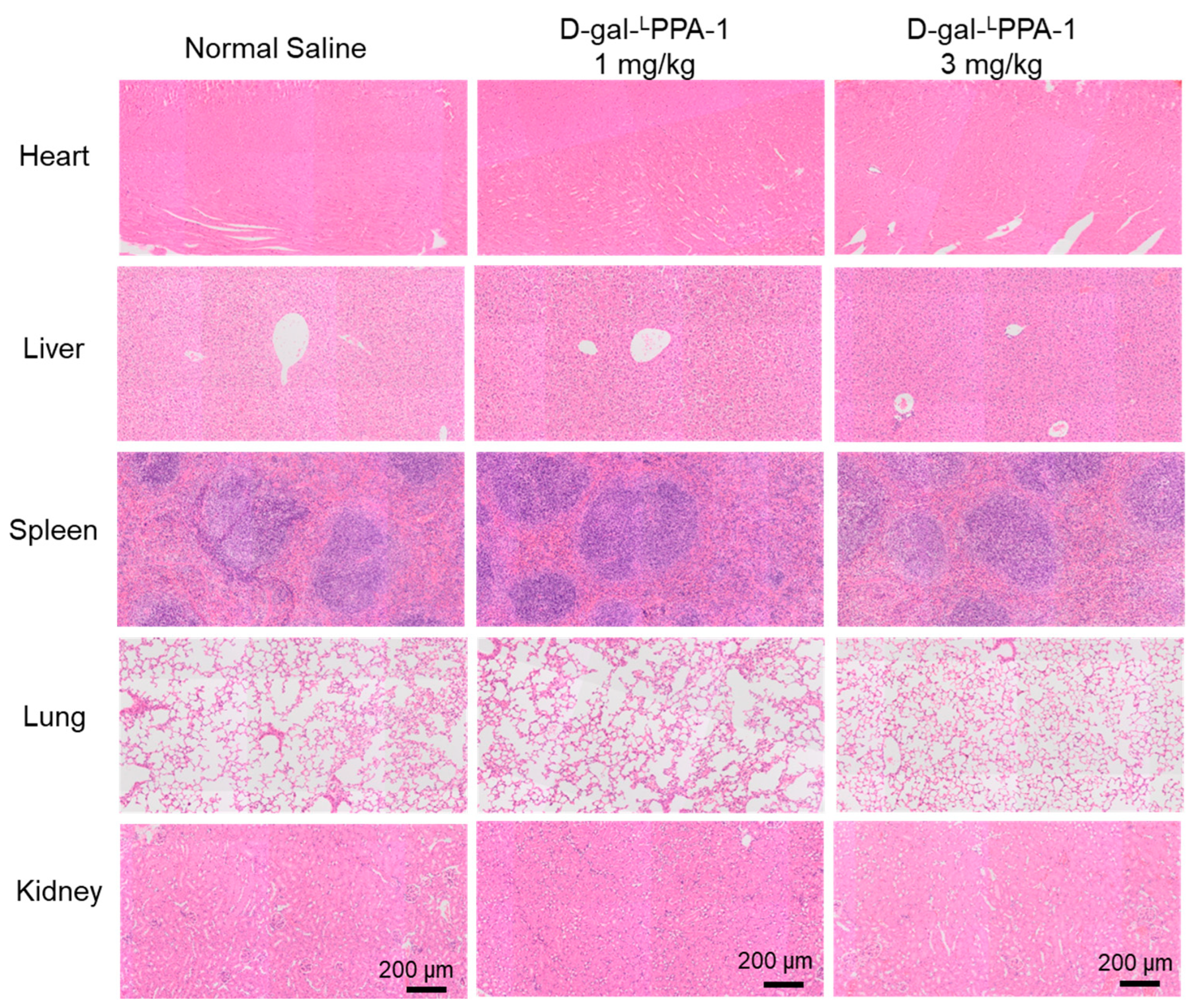
2.6. The Toxicity of D-gal-LPPA-1 In Vivo
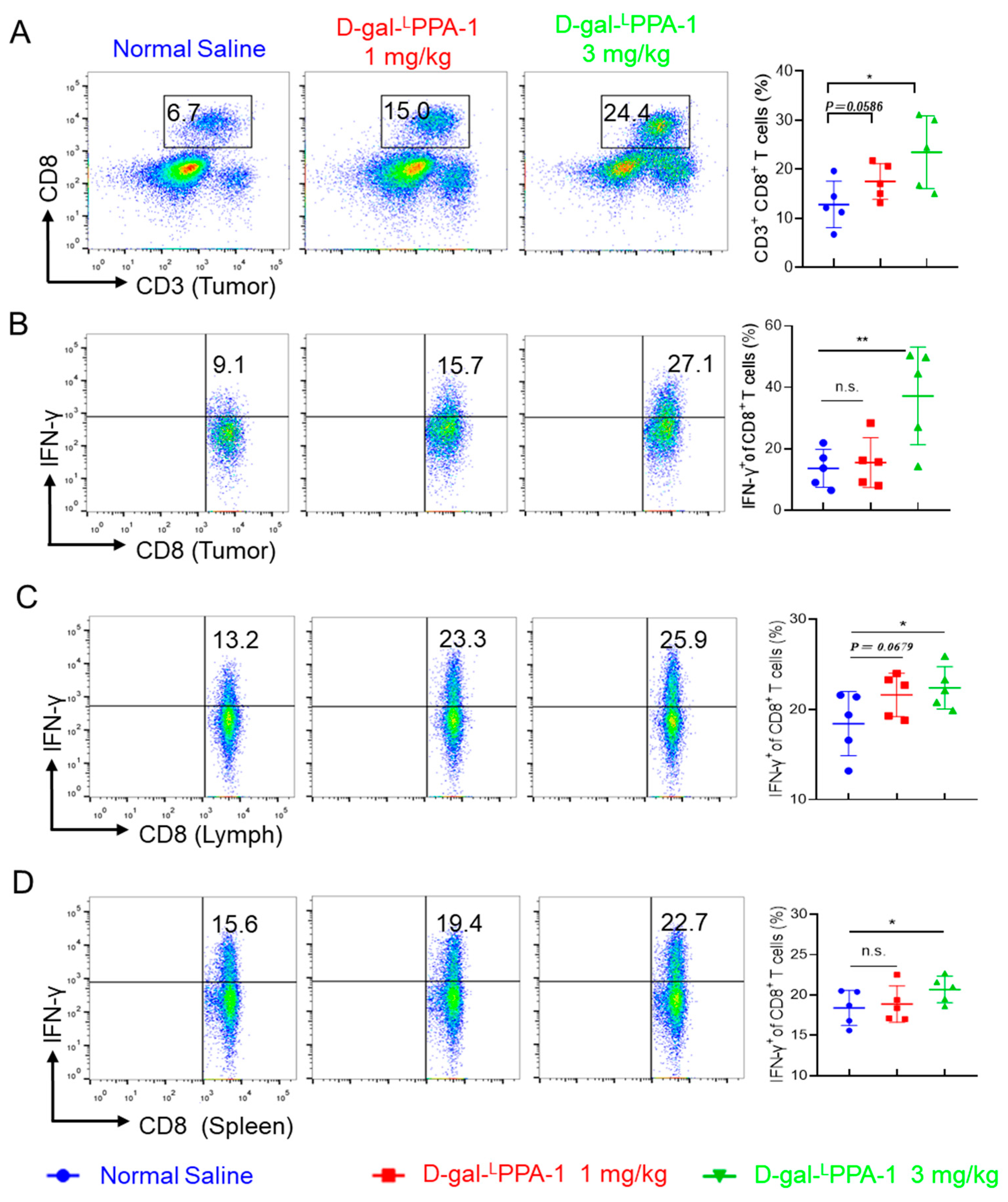
2.7. D-gal-LPPA-1 Significantly Enhances the Functionality of CD8+ T Cells
3. Materials and Methods
3.1. Solid-Phase Peptide Synthesis
3.1.1. Preparation of Fmoc-Phe-CTC Resin

3.1.2. Preparation of Phe-CTC Resin
3.1.3. Preparation of Fmoc-His-Phe-CTC Resin
3.1.4. Preparation of D-glu-Asn-Tyr-Ser-Lys-Pro-Thr-Asp-Arg-Gln-Tyr-His-Phe-CTC Resin
3.1.5. Preparation of D-gal-Asn-Tyr-Ser-Lys-Pro-Thr-Asp-Arg-Gln-Tyr-His-Phe-CTC Resin
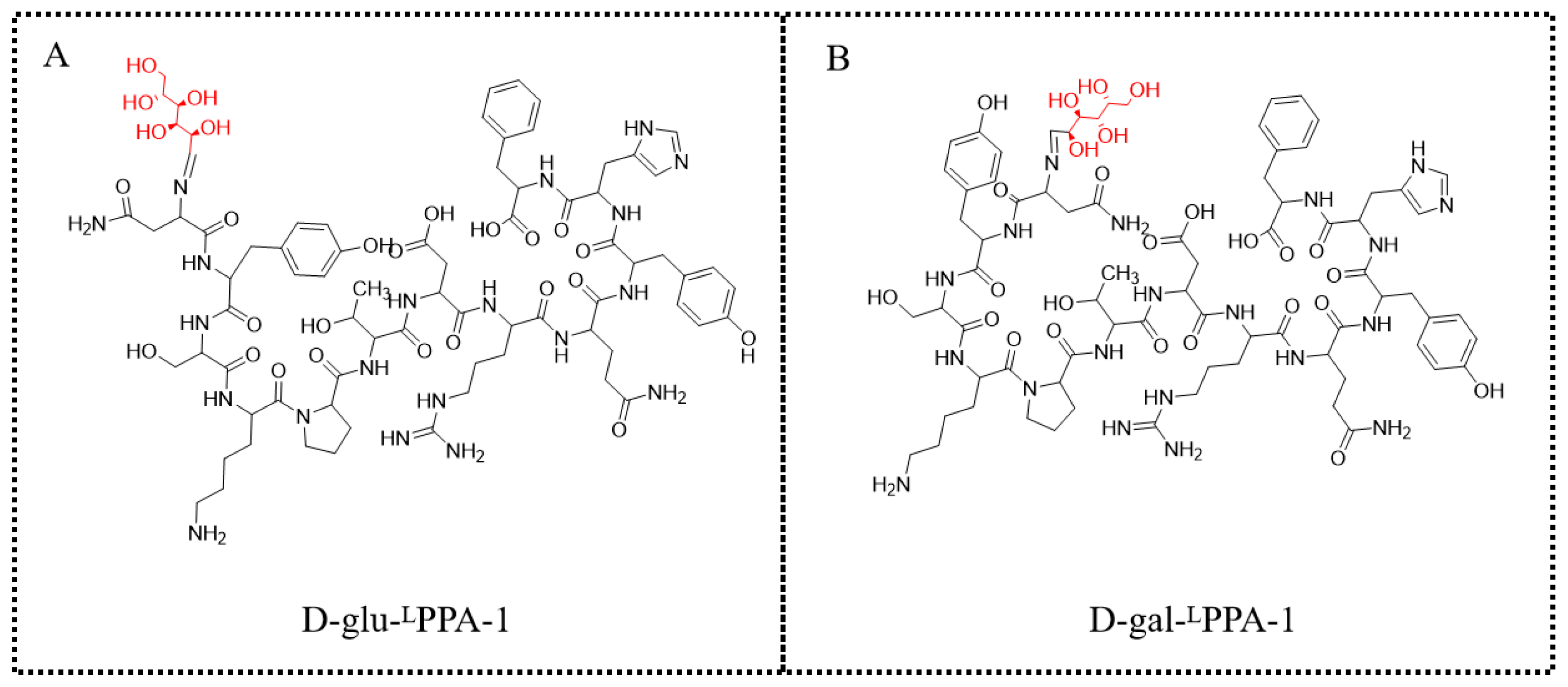
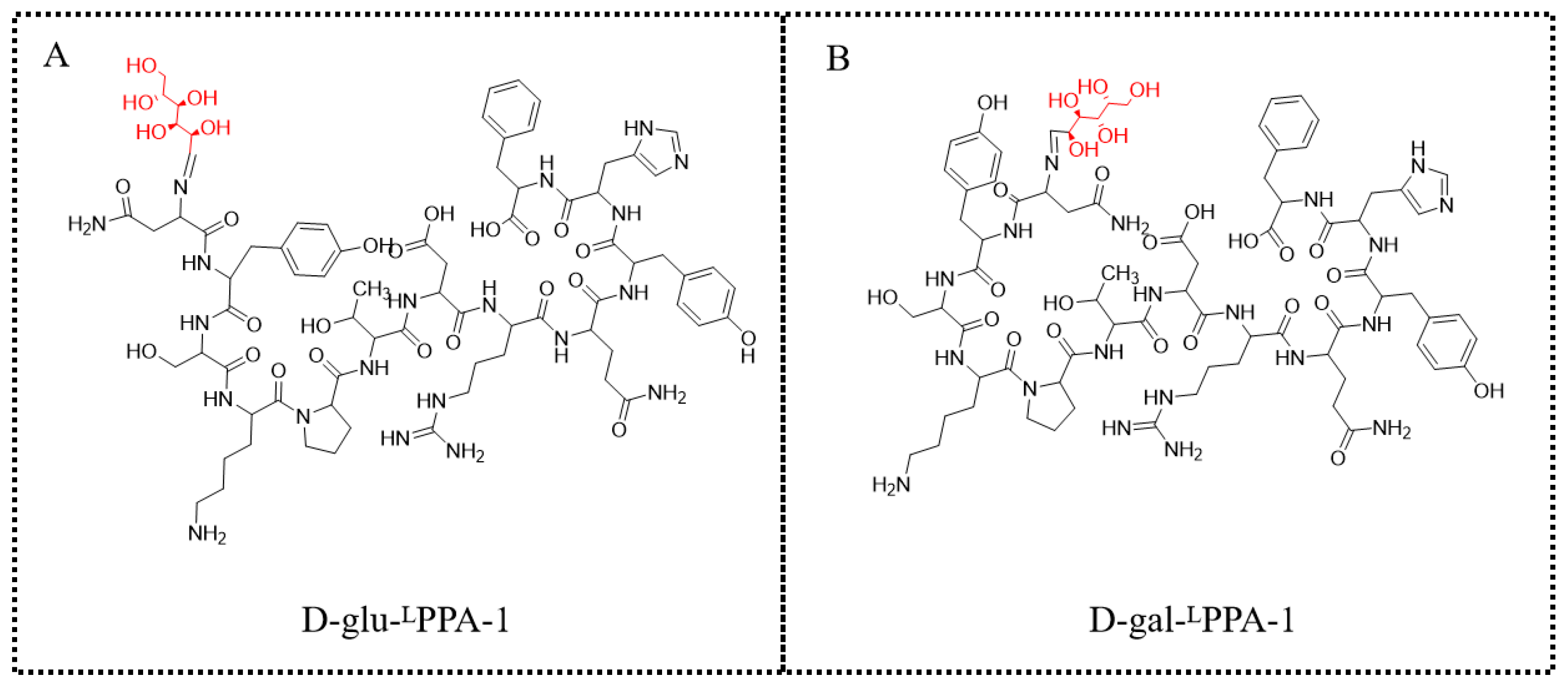
3.1.6. Preparation of D-glu-Asn-Tyr-Ser-Lys-Pro-Thr-Asp-Arg-Gln-Tyr-His-Phe (D-glu-LPPA-1, Figure 8A)
3.1.7. Preparation of D-gal-Asn-Tyr-Ser-Lys-Pro-Thr-Asp-Arg-Gln-Tyr-His-Phe (D-gal-LPPA-1, Figure 8B)
3.2. Chemicals and Reagents
3.3. Cell Culture
3.4. Peptide Blocking Assay
3.5. Binding Assay
3.6. Coculture Assay
3.7. In Vivo Antitumor Experiment
3.8. H&E Staining Analysis
3.9. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Hermjakob, H.; Sharon, N. On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim. Biophys. Acta 1999, 1473, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Biological roles of oligosaccharides: All of the theories are correct. Glycobiology 1993, 3, 97–130. [Google Scholar] [CrossRef] [PubMed]
- Satoshi, N.; Ginto, G.; Kazutoshi, M. Mechanisms of productive folding and endoplasmic reticulum-associated degradation of glycoproteins and non-glycoproteins. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129812. [Google Scholar]
- Grabarics, M.; Lettow, M.; Kirschbaum, C.; Greis, K.; Manz, C.; Pagel, K. Mass Spectrometry-Based Techniques to Elucidate the Sugar Code. Chem. Rev. 2022, 122, 7840–7908. [Google Scholar] [CrossRef] [PubMed]
- Herzner, H.; Reipen, T.; Schultz, M.; Kunz, H. Synthesis of glycopeptides containing carbohydrate and Peptide recognition motifs. Chem. Rev. 2000, 100, 4495–4538. [Google Scholar] [CrossRef] [PubMed]
- Doelman, W.; van Kasteren, S.I. Synthesis of glycopeptides and glycopeptide conjugates. Org. Biomol. Chem. 2022, 20, 6487–6507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L. Roles of glycans and glycopeptides in immune system and immunerelated diseases. Curr. Med. Chem. 2006, 13, 1141–1147. [Google Scholar] [CrossRef]
- Stergiou, N.; Urschbach, M.; Gabba, A.; Schmitt, E.; Kunz, H.; Besenius, P. The Development of Vaccines from Synthetic Tumor-Associated Mucin Glycopeptides and their Glycosylation-Dependent Immune Response. Chem. Rec. 2021, 21, 3313–3331. [Google Scholar] [CrossRef] [PubMed]
- Acharya, Y.; Bhattacharyya, S.; Dhanda, G.; Haldar, J. Emerging Roles of Glycopeptide Antibiotics: Moving beyond Gram-Positive Bacteria. ACS Infect. Dis. 2022, 8, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.V.; Hussein, W.M.; Varamini, P.; Simerska, P.; Toth, I. Glycosylation, an effective synthetic strategy to improve the bioavailability of therapeutic peptides. Chem. Sci. 2016, 7, 2492–2500. [Google Scholar] [CrossRef] [PubMed]
- Dermani, F.K.; Samadi, P.; Rahmani, G.; Kohlan, A.K.; Najafi, R. PD-1/PD-L1 immune checkpoint: Potential target for cancer therapy. J. Cell. Physiol. 2019, 234, 1313–1325. [Google Scholar] [CrossRef]
- Xu-Monette, Z.Y.; Zhou, J.; Young, K.H. PD-1 expression and clinical PD-1 blockade in Bcell lymphomas. Blood 2018, 131, 68–83. [Google Scholar] [CrossRef]
- Simon, S.; Labarriere, N. PD-1 expression on tumor-specific T cells: Friend or foe for immunotherapy? Oncoimmunology 2017, 7, e1364828. [Google Scholar] [CrossRef] [PubMed]
- Ai, L.; Xu, A.; Xu, J. Roles of PD-1/PD-L1 Pathway: Signaling, Cancer, and Beyond. Adv. Exp. Med. Biol. 2020, 1248, 33–59. [Google Scholar] [PubMed]
- Pauken, K.E.; Wherry, E.J. Overcoming T cell exhaustion in infection and cancer. Trends Immunol. 2015, 36, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef]
- Mimura, K.; Teh, J.L.; Okayama, H.; Shiraishi, K.; Kua, L.F.; Koh, V.; Smoot, D.T.; Ashktorab, H.; Oike, T.; Suzuki, Y.; et al. PD-L1 expression is mainly regulated by interferon gamma associated with JAK-STAT pathway in gastric cancer. Cancer Sci. 2018, 109, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Blank, C.; Gajewski, T.F.; Mackensen, A. Interaction of PD-L1 on tumor cells with PD-1 on tumor-specific T cells as a mechanism of immune evasion: Implications for tumor immunotherapy. Cancer Immunol. Immunother. 2005, 54, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Xin Yu, J.; Hodge, J.P.; Oliva, C.; Neftelinov, S.T.; Hubbard-Lucey, V.M.; Tang, J. Trends in clinical development for PD-1/PD-L1 inhibitors. Nat. Rev. Drug Discov. 2020, 19, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.N.; Liu, B.Y.; Qi, Y.K.; Zhou, Y.; Chen, Y.P.; Pan, K.M.; Li, W.W.; Zhou, X.M.; Ma, W.W.; Fu, C.Y.; et al. Blocking of the PD-1/PD-L1 Interaction by a D-Peptide Antagonist for Cancer Immunotherapy. Angew. Chem. Int. Ed. Engl. 2015, 54, 11760–11764. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Li, W.; Chen, S.; Chen, D.; Xu, R.; Zheng, D.; Yang, X.; Li, S.; Zhou, X.; Niu, X.; et al. Design of a novel chimeric peptide via dual blockade of CD47/SIRPα and PD-1/PD-L1 for cancer immunotherapy. Sci. China Life Sci. 2023, 66, 2310–2328. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Dong, Q.; Zhai, W.; Zhao, W.; Shi, P.; Wu, Y.; Zhou, X.; Gao, Y. A PD-L1 and VEGFR2 dual targeted peptide and its combination with irradiation for cancer immunotherapy. Pharmacol. Res. 2022, 182, 106343. [Google Scholar] [CrossRef]
- Li, W.; Zhu, X.; Zhou, X.; Wang, X.; Zhai, W.; Li, B.; Du, J.; Li, G.; Sui, X.; Wu, Y.; et al. An orally available PD-1/PD-L1 blocking peptide OPBP-1-loaded trimethyl chitosan hydrogel for cancer immunotherapy. J. Control. Release 2021, 334, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Muttenthaler, M.; King, G.F.; Adams, D.J.; Alewood, P.F. Trends in peptide drug discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Niu, X.; Zhou, X.; Gao, Y. Peptide drugs: A new direction in cancer immunotherapy. Cancer Biol. Med. 2024, 21, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Liu, C.A.; Ding, Y.Q.; Ni, Y.B.; Ji, F.F.; Lau, H.C.H.; Jiang, L.P.; Sung, J.J.; Wong, S.H.; Yu, J. Roseburia intestinalis generated butyrate boosts anti-PD-1 efficacy in colorectal cancer by activating cytotoxic CD8+ T cells. Gut 2023, 72, 2112–2122. [Google Scholar] [CrossRef]
- Pan, C.; Yang, H.; Lu, Y.; Hu, S.; Wu, Y.; He, Q.; Dong, X. Recent advance of peptide-based molecules and nonpeptidic small-molecules modulating PD-1/PD-L1 proteinprotein interaction or targeting PD-L1 protein degradation. Eur. J. Med. Chem. 2021, 213, 113170. [Google Scholar] [CrossRef] [PubMed]
- Dammeijer, F.; van Gulijk, M.; Mulder, E.E.; Lukkes, M.; Klaase, L.; van den Bosch, T.; van Nimwegen, M.; Lau, S.P.; Latupeirissa, K.; Schetters, S.; et al. The PD-1/PD-L1-Checkpoint Restrains T cell Immunity in Tumor-Draining Lymph Nodes. Cancer Cell 2020, 38, 685–700.e8. [Google Scholar] [CrossRef] [PubMed]
- Juneja, V.R.; McGuire, K.A.; Manguso, R.T.; LaFleur, M.W.; Collins, N.; Haining, W.N.; Freeman, G.J.; Sharpe, A.H. PD-L1 on tumor cells is sufficient for immune evasion in immunogenic tumors and inhibits CD8 T cell cytotoxicity. J. Exp. Med. 2017, 214, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Song, C.H.; Kim, N.; Nam, R.H.; Choi, S.I.; Jang, J.Y.; Kim, J.W.; Na, H.Y.; Lee, H.N. Combination treatment with 17β-estradiol and anti-PD-L1 suppresses MC38 tumor growth by reducing PD-L1 expression and enhancing M1 macrophage population in MC38 colon tumor model. Cancer Lett. 2022, 543, 215780. [Google Scholar] [CrossRef]
- Qin, S.; Yu, Y.; Guan, H.; Yang, Y.; Sun, F.; Sun, Y.; Zhu, J.; Xing, L.; Yu, J.; Sun, X. A preclinical study: Correlation between PD-L1 PET imaging and the prediction of therapy efficacy of MC38 tumor with 68Ga-labeled PD-L1 targeted nanobody. Aging 2021, 13, 13006–13022. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, B.; Li, L.; Wang, G.; Li, Y.; Fu, R.; Ming, Y.; Ni, R.; Wang, J.; Ye, G.; et al. The Immunostimulative Effect and Mechanisms of a Novel Mouse Anti-Human PD-1 Monoclonal Antibody on Jurkat Lymphocytic Cells Cocultured with Hepatoma Cells. OncoTargets Ther. 2020, 13, 12225–12241. [Google Scholar] [CrossRef]
- Shen, W.H.; Shi, P.S.; Dong, Q.Y.; Zhou, X.M.; Chen, C.X.; Sui, X.H.; Tian, W.T.; Zhu, X.Q.; Wang, X.X.; Jin, S.Z.; et al. Discovery of a novel dual-targeting D-peptide to block CD24/Siglec-10 and PD-1/PD-L1 interaction and synergize with radiotherapy for cancer immunotherapy. J. Immunother. Cancer 2023, 11, e007068. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Y.; Li, J.X.; Chen, B.; Shuai, Y.; He, X.Y.; Liu, K.; He, M.D.; Jin, L. Dental pulp stem cells induce anti-inflammatory phenotypic transformation of macrophages to enhance osteogenic potential via IL-6/GP130/STAT3 signaling. Ann. Transl. Med. 2023, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, Y.; Zhang, X.; Li, B.; Jin, S.; Wu, M.; Zhou, X.; Dong, Q.; Du, J.; Zhai, W.; et al. Hemin blocks TIGIT/PVR interaction and induces ferroptosis to elicit synergistic effects of cancer immunotherapy. Sci. China Life Sci. 2024. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, P.; Dong, X.; Wu, Z.; Hou, X.; Mao, L.; Guo, J.; Zhao, W.; Peng, C.; Zhang, Z.; Peng, L. Development of Glycosylation-Modified DPPA-1 Compounds as Innovative PD-1/PD-L1 Blockers: Design, Synthesis, and Biological Evaluation. Molecules 2024, 29, 1898. https://doi.org/10.3390/molecules29081898
Deng P, Dong X, Wu Z, Hou X, Mao L, Guo J, Zhao W, Peng C, Zhang Z, Peng L. Development of Glycosylation-Modified DPPA-1 Compounds as Innovative PD-1/PD-L1 Blockers: Design, Synthesis, and Biological Evaluation. Molecules. 2024; 29(8):1898. https://doi.org/10.3390/molecules29081898
Chicago/Turabian StyleDeng, Peng, Xiaodan Dong, Ziyuan Wu, Xixi Hou, Longfei Mao, Jingjing Guo, Wenshan Zhao, Chune Peng, Zhe Zhang, and Lizeng Peng. 2024. "Development of Glycosylation-Modified DPPA-1 Compounds as Innovative PD-1/PD-L1 Blockers: Design, Synthesis, and Biological Evaluation" Molecules 29, no. 8: 1898. https://doi.org/10.3390/molecules29081898
APA StyleDeng, P., Dong, X., Wu, Z., Hou, X., Mao, L., Guo, J., Zhao, W., Peng, C., Zhang, Z., & Peng, L. (2024). Development of Glycosylation-Modified DPPA-1 Compounds as Innovative PD-1/PD-L1 Blockers: Design, Synthesis, and Biological Evaluation. Molecules, 29(8), 1898. https://doi.org/10.3390/molecules29081898