

Purine but Not Pyrimidine De Novo Nucleotide Biosynthesis Inhibitors Strongly Enhance the Antiviral Effect of Corresponding Nucleobases Against Dengue Virus


Abstract
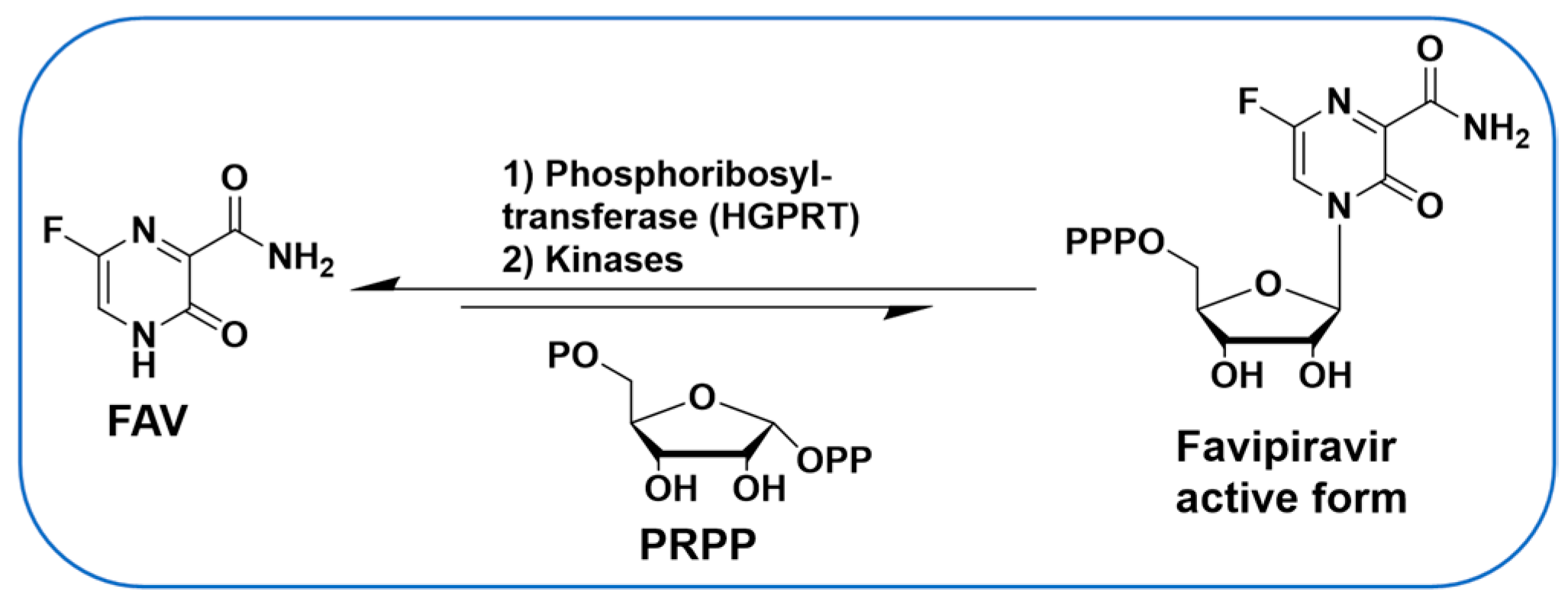
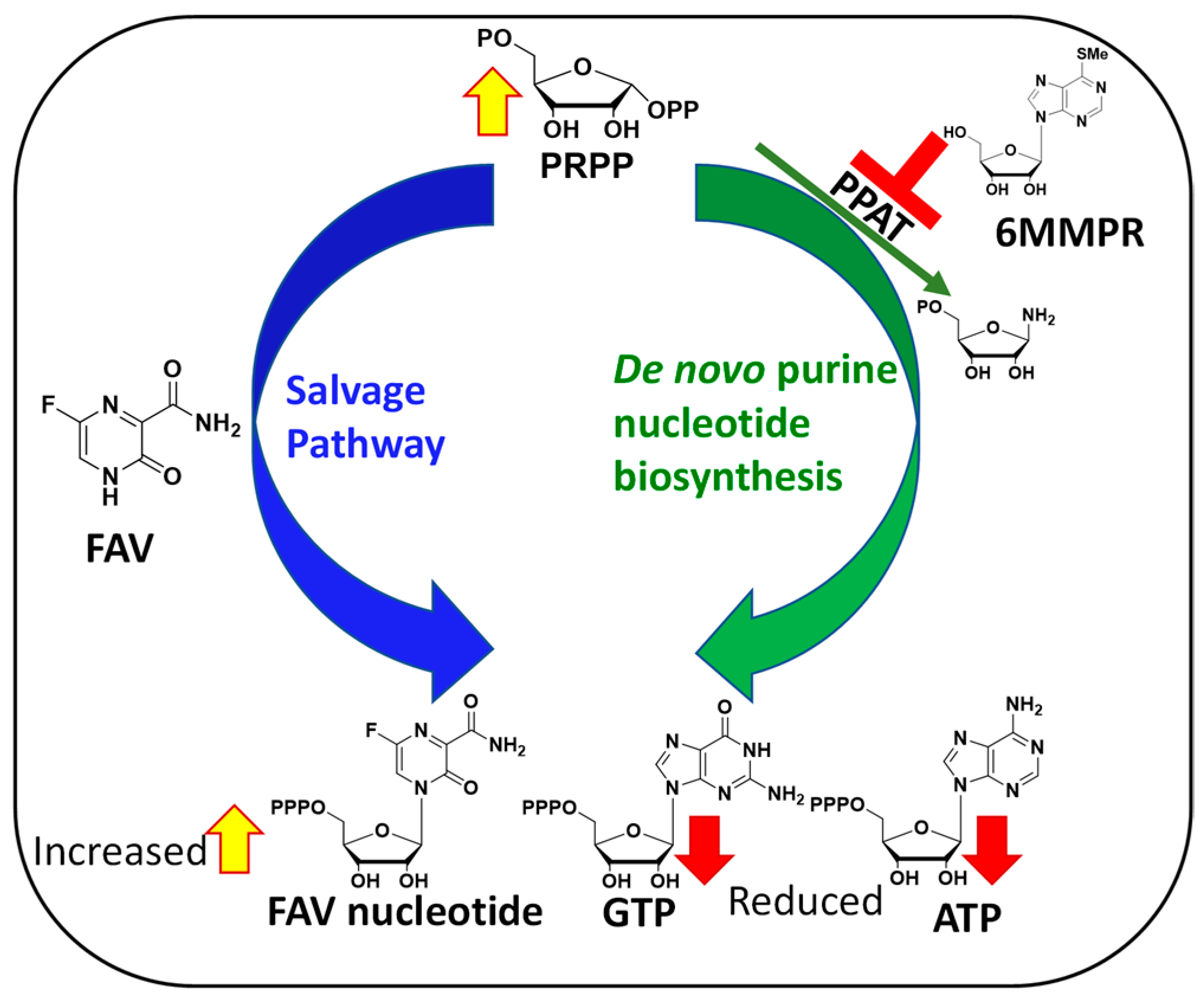
1. Introduction
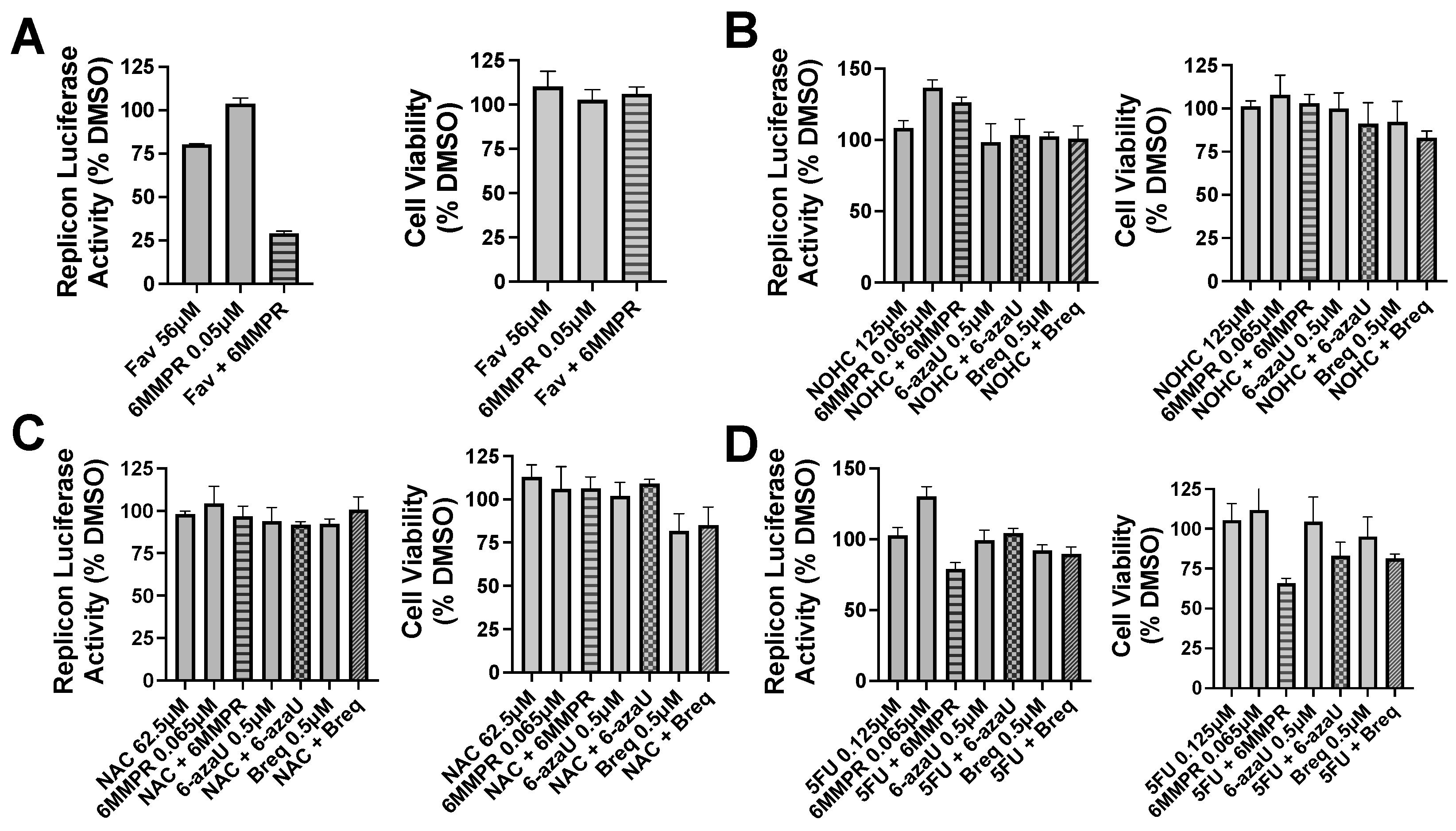
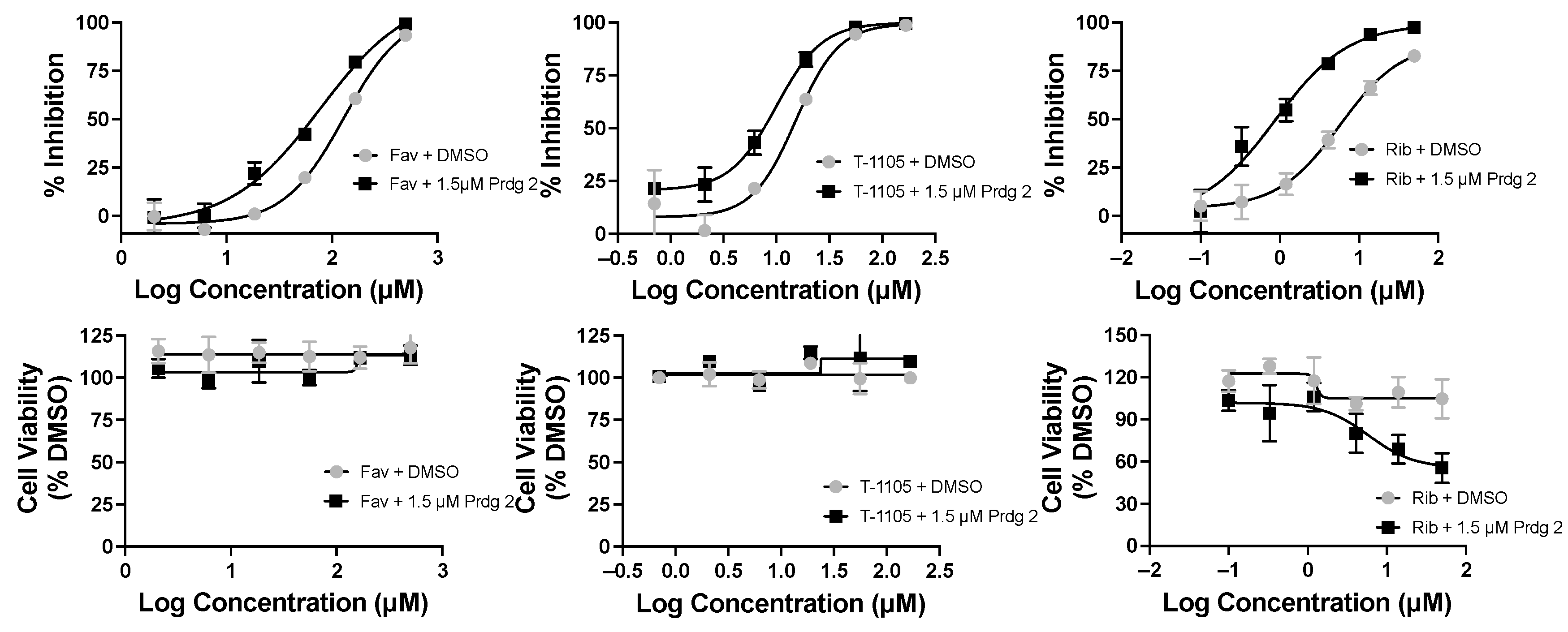
2. Results
3. Discussion
4. Materials and Methods
4.1. Cells and Viruses
4.2. Cell Viability Assay (MTS)
4.3. DENV Replicon Cell Assay
4.4. ZIKV Reporter Virus Assay
4.5. DENV CPE Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furuta, Y.; Takahashi, K.; Fukuda, Y.; Kuno, M.; Kamiyama, T.; Kozaki, K.; Nomura, N.; Egawa, H.; Minami, S.; Watanabe, Y.; et al. In Vitro and In Vivo Activities of Anti-Influenza Virus Compound T-705. Antimicrob. Agents Chemother. AAC 2002, 46, 977–981. [Google Scholar] [CrossRef]
- Furuta, Y.; Takahashi, K.; Kuno-Maekawa, M.; Sangawa, H.; Uehara, S.; Kozaki, K.; Nomura, N.; Egawa, H.; Shiraki, K. Mechanism of Action of T-705 against Influenza Virus. Antimicrob. Agents Chemother. AAC 2005, 49, 981–986. [Google Scholar] [CrossRef]
- Furuta, Y.; Gowen, B.B.; Takahashi, K.; Shiraki, K.; Smee, D.F.; Barnard, D.L. Favipiravir (T-705), a novel viral RNA polymerase inhibitor. Antivir. Res. 2013, 100, 446–454. [Google Scholar] [CrossRef]
- Julander, J.G.; Shafer, K.; Smee, D.F.; Morrey, J.D.; Furuta, Y. Activity of T-705 in a hamster model of yellow fever virus infection in comparison with that of a chemically related compound, T-1106. Antimicrob. Agents Chemother. 2009, 53, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Naesens, L.; Guddat, L.W.; Keough, D.T.; van Kuilenburg, A.B.P.; Meijer, J.; Vande Voorde, J.; Balzarini, J. Role of Human Hypoxanthine Guanine Phosphoribosyltransferase in Activation of the Antiviral Agent T-705 (Favipiravir). Mol. Pharmacol. 2013, 84, 615–629. [Google Scholar] [CrossRef]
- Stein, D.S.; Moore, K.H.P. Phosphorylation of Nucleoside Analog Antiretrovirals: A Review for Clinicians. Pharmacotherapy 2001, 21, 11–34. [Google Scholar] [CrossRef] [PubMed]
- Van Rompay, A.R.; Johansson, M.; Karlsson, A. Phosphorylation of nucleosides and nucleoside analogs by mammalian nucleoside monophosphate kinases. Pharmacol. Ther. 2000, 87, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Van Rompay, A.R.; Johansson, M.; Karlsson, A. Substrate specificity and phosphorylation of antiviral and anticancer nucleoside analogues by human deoxyribonucleoside kinases and ribonucleoside kinases. Pharmacol. Ther. 2003, 100, 119–139. [Google Scholar] [CrossRef]
- Qiu, L.; Patterson, S.E.; Bonnac, L.F.; Geraghty, R.J.; Abd-Alla, A.M.M. Nucleobases and corresponding nucleosides display potent antiviral activities against dengue virus possibly through viral lethal mutagenesis. PLoS Neglected Trop. Dis. 2018, 12, e0006421. [Google Scholar] [CrossRef]
- Soto-Acosta, R.; Edwards, T.C.; Dreis, C.D.; Krishna, V.D.; Cheeran, M.C.J.; Qiu, L.; Xie, J.; Bonnac, L.F.; Geraghty, R.J. Enhancing the Antiviral Potency of Nucleobases for Potential Broad-Spectrum Antiviral Therapies. Viruses 2021, 13, 2508. [Google Scholar] [CrossRef] [PubMed]
- Thaker, S.K.; Ch’ng, J.; Christofk, H.R. Viral hijacking of cellular metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Uemura, K.; Nobori, H.; Sato, A.; Toba, S.; Kusakabe, S.; Sasaki, M.; Tabata, K.; Matsuno, K.; Maeda, N.; Ito, S.; et al. 2-thiouridine is a broad-spectrum antiviral nucleoside analogue against positive-strand RNA viruses. Proc. Natl. Acad. Sci. USA 2023, 120, e2304139120. [Google Scholar] [CrossRef]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, P.; Solanki, K.; Li, Y.; Ma, Z.; Peppelenbosch, M.P.; Baig, M.S.; Pan, Q. Viral polymerase binding and broad-spectrum antiviral activity of molnupiravir against human seasonal coronaviruses. Virology 2021, 564, 33–38. [Google Scholar] [CrossRef]
- Karim, M.; Lo, C.-W.; Einav, S. Preparing for the next viral threat with broad-spectrum antivirals. J. Clin. Investig. 2023, 133, e170236. [Google Scholar] [CrossRef] [PubMed]
- Geraghty, R.J.; Geraghty, R.; Aliota, M.; Bonnac, L. Broad-Spectrum Antiviral Strategies and Nucleoside Analogues. Viruses 2021, 13, 667. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.-Q.; Mu, J.-S.; Kargbo, D.; Song, Y.-B.; Niu, W.-K.; Nie, W.-M.; Kanu, A.; Liu, W.-W.; Wang, Y.-P.; Dafae, F.; et al. Clinical and Virological Characteristics of Ebola Virus Disease Patients Treated With Favipiravir (T-705)-Sierra Leone, 2014. Clin. Infect. Dis. 2016, 63, 1288–1294. [Google Scholar] [CrossRef]
- Doi, Y.; Hibino, M.; Hase, R.; Yamamoto, M.; Kasamatsu, Y.; Hirose, M.; Mutoh, Y.; Homma, Y.; Terada, M.; Ogawa, T.; et al. A Prospective, Randomized, Open-Label Trial of Early versus Late Favipiravir Therapy in Hospitalized Patients with COVID-19. Antimicrob. Agents Chemother. 2020, 64, e01897. [Google Scholar] [CrossRef] [PubMed]
- Hayden, F.G.; Lenk, R.P.; Epstein, C.; Kang, L.L. Oral Favipiravir Exposure and Pharmacodynamic Effects in Adult Outpatients With Acute Influenza. J. Infect. Dis. 2024, 230, e395–e404. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.T.; Guedj, J.; Anglaret, X.; Laouénan, C.; Madelain, V.; Taburet, A.-M.; Baize, S.; Sissoko, D.; Pastorino, B.; Rodallec, A.; et al. Favipiravir pharmacokinetics in Ebola-Infected patients of the JIKI trial reveals concentrations lower than targeted. PLoS Neglected Trop. Dis. 2017, 11, e0005389. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, D.; Laouenan, C.; Folkesson, E.; M’Lebing, A.-B.; Beavogui, A.-H.; Baize, S.; Camara, A.-M.; Maes, P.; Shepherd, S.; Danel, C.; et al. Experimental Treatment with Favipiravir for Ebola Virus Disease (the JIKI Trial): A Historically Controlled, Single-Arm Proof-of-Concept Trial in Guinea. PLoS Med. 2016, 13, e1001967. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Y.; Khan, N.; Close, B.J.; Goel, R.K.; Blum, B.; Tavares, A.H.; Kenney, D.; Conway, H.L.; Ewoldt, J.K.; Chitalia, V.C.; et al. SARS-CoV-2 Disrupts Proximal Elements in the JAK-STAT Pathway. J. Virol. 2021, 95, e0086221. [Google Scholar] [CrossRef]
- Soto-Acosta, R.; Jung, E.; Qiu, L.; Wilson, D.J.; Geraghty, R.J.; Chen, L. 4,7-Disubstituted 7H-Pyrrolo[2,3-d]pyrimidines and Their Analogs as Antiviral Agents against Zika Virus. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | EC50 (μM) | CC50 (μM) |
|---|---|---|
| 2-thiouracil (2SU) | >50 | >50 |
| 2-thiouridine | >50 | >50 |
| 5-fluorouracil (5FU) | 0.74 | 0.93 |
| N-hydroxy-cytidine (NOHC) | >100 | >100 |
| Molnupiravir | 6.2 | 53 |
| N-amino-cytidine (NAC) | >100 | >100 |
| 6-azauridine | 1.9 | 8.8 |
| Brequinar | 3.0 | 4.5 |
| Compound | EC50 (μM) | CC50 (μM) |
|---|---|---|
| Favipiravir (FAV) | 114 | >500 |
| T-1105 | 13 | >500 |
| Ribavirin nucleobase | 5.7 | >100 |
| 6MMPR | 0.21 | 0.21 |
| Prdg 1 | 0.26 | 2.6 |
| Prdg 2 | 2.5 | 5.4 |
| Prdg 3 | 2.3 | 2.4 |
| Compounds | EC50 (μM) | CC50 (μM) |
|---|---|---|
| Favipiravir (Fav) | 128 | >500 |
| +6MMPR 0.065 μM | 63 | >500 |
| +Prdg 1 0.09 μM | 51 | >500 |
| +Prdg 2 1.5 μM | 47 | >500 |
| Favipiravir (Fav) | 90 | >500 |
| +Prdg 3 1.4 μM | 46 | >500 |
| T-1105 | 15 | >167 |
| +6MMPR 0.065 μM | 11 | >167 |
| +Prdg 1 0.09 μM | 8.3 | >167 |
| +Prdg 2 1.5 μM | 10 | >167 |
| +Prdg 3 1.4 μM | 9.6 | >167 |
| Ribavirin nucleobase | 5.7 | >50 |
| +6MMPR 0.065 μM | 3.4 | >50 |
| +Prdg 1 0.09 μM | 2.3 | >50 |
| +Prdg 2 1.5 μM | 0.88 | >50 |
| +Prdg 3 1.4 μM | 2.6 | >50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnac, L.F.; Dreis, C.D.; Rai, M.; Geraghty, R.J. Purine but Not Pyrimidine De Novo Nucleotide Biosynthesis Inhibitors Strongly Enhance the Antiviral Effect of Corresponding Nucleobases Against Dengue Virus. Molecules 2025, 30, 210. https://doi.org/10.3390/molecules30020210
Bonnac LF, Dreis CD, Rai M, Geraghty RJ. Purine but Not Pyrimidine De Novo Nucleotide Biosynthesis Inhibitors Strongly Enhance the Antiviral Effect of Corresponding Nucleobases Against Dengue Virus. Molecules. 2025; 30(2):210. https://doi.org/10.3390/molecules30020210
Chicago/Turabian StyleBonnac, Laurent F., Christine D. Dreis, Madhu Rai, and Robert J. Geraghty. 2025. "Purine but Not Pyrimidine De Novo Nucleotide Biosynthesis Inhibitors Strongly Enhance the Antiviral Effect of Corresponding Nucleobases Against Dengue Virus" Molecules 30, no. 2: 210. https://doi.org/10.3390/molecules30020210
APA StyleBonnac, L. F., Dreis, C. D., Rai, M., & Geraghty, R. J. (2025). Purine but Not Pyrimidine De Novo Nucleotide Biosynthesis Inhibitors Strongly Enhance the Antiviral Effect of Corresponding Nucleobases Against Dengue Virus. Molecules, 30(2), 210. https://doi.org/10.3390/molecules30020210