Analysis of Intestinal Injuries Induced by Ricin in Vitro Using SPR Technology and MS Identification

Abstract
:
1. Introduction
2. Experimental Section
2.1. Materials
2.2. Brush Border Membrane Protein (BBMP) preparation [6]
2.3. Preparation of biosensor chips
2.4. Kinetics analysis
2.5. Mass-Spectrum identification
3. Results and Discussion
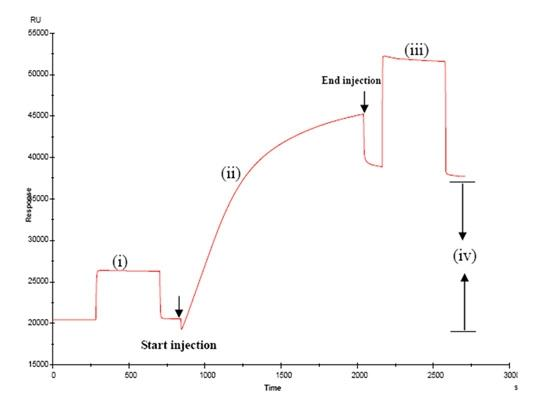
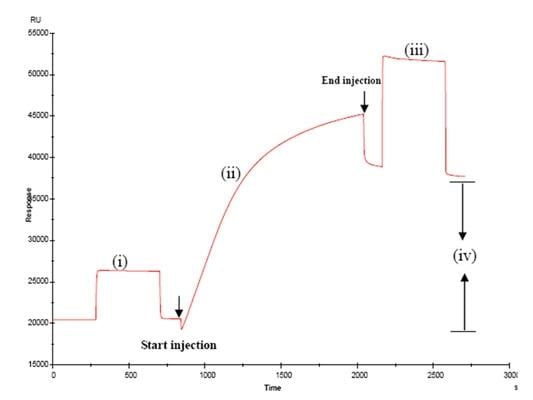
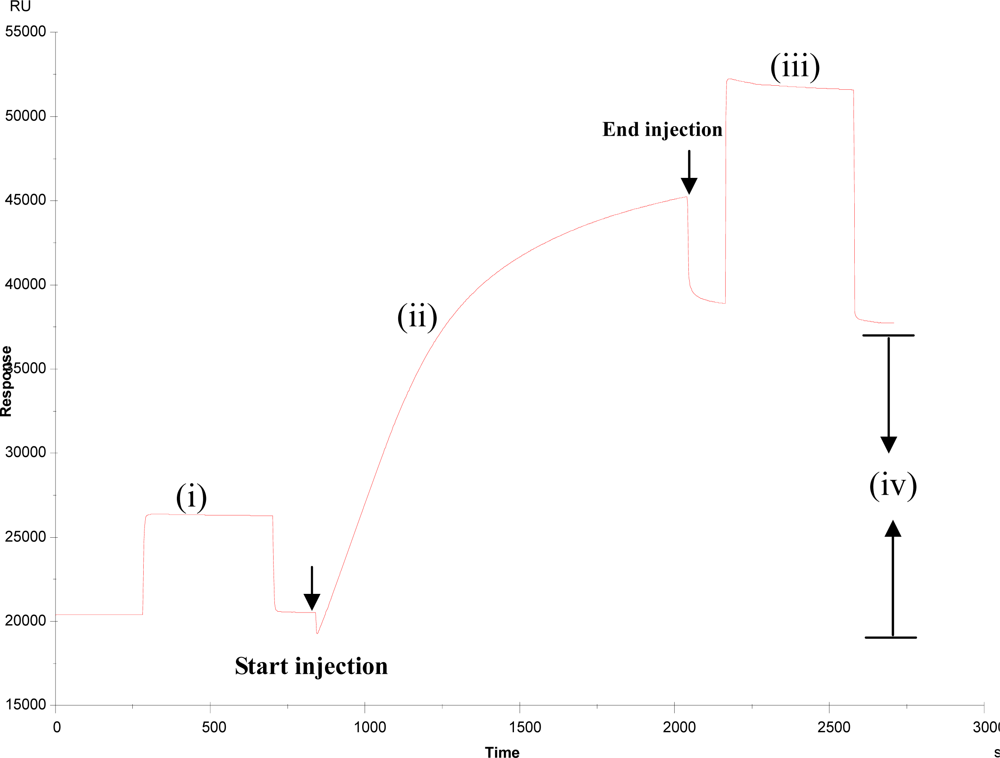
3.1. Immobilisation of BBMPs on the surface of sensor chip
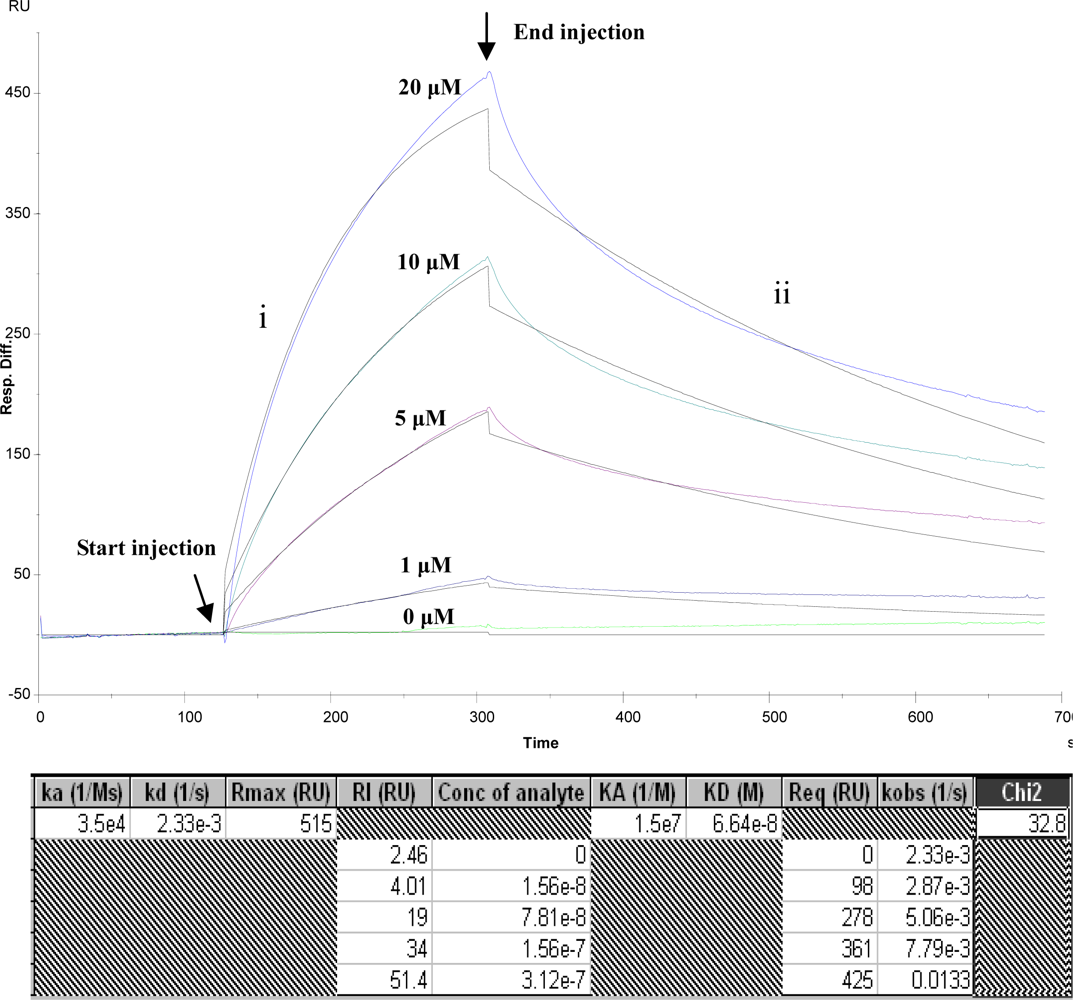
3.2. Kinetics analysis of BBMP and ricin
3.3. Recovery and identification of interaction components
4. Conclusions
Acknowledgments
Referances
- Terrettaz, S; Stora, T; Duschl, C; Vogel, H. Protein binding to supported lipid membranes: investigation of the cholera toxinganglioside interaction by simultaneous impedance spectroscopy and surface plasmon resonance. Langmuir 1993, 9, 1361–1369. [Google Scholar]
- Glasmästar, K; Larsson, C; Höök, F; Kasemo, B. Protein adsorption on supported phospholipid bilayers. J. Colloid Interface Sci 2002, 246, 40–47. [Google Scholar]
- Myszka, DG; Rich, RL. Implementing surface plasmon resonance biosensors in drug discovery. Pharm. Sci. Technol. Today 2000, 3, 310–317. [Google Scholar]
- Buijs, J; Franklin, GC. SPR-MS in functional proteomics. Brief. Funct. Genomic. Proteomic 2005, 4, 39–48. [Google Scholar]
- Summary of the NIAID Ricin Expert Panel Workshop, Bethesda, MD, USA, April, 1–2, 2004.
- Cho, S; Park, JH; Yu, J; Lee, YK; Byun, Y; Chung, H; Kwon, IC; Jeong, SY. Preparation and characterization of reconstructed small intestinal brush border membranes for surface plasmon resonance analysis. Pharm. Res 2004, 21, 55–62. [Google Scholar]
- Cooper, MA; Hansson, A; Löfås, S; Williams, DH. A vesicle capture sensor chip for kinetic analysis of interactions with membrane-bound receptors. Anal. Biochem 2000, 277, 196–205. [Google Scholar]
- Cooper, MA; Try, AC; Carroll, J; Ellar, DJ; Williams, DH. Surface plasmon resonance analysis at a supported lipid monolayer. Biochim. Biophy. Acta 1998, 1373, 101–111. [Google Scholar]
- Kramer, W; Wess, G; Neckermann, G; Schubert, G; Fink, J; Girbig, F; Gutjahr, U; Kowalewski, S; Baringhaus, KH; Bor, G. Intestinal absorption of peptides by coupling to bile acids. J. Biol. Chem 1994, 269, 10621–10627. [Google Scholar]
- Nowicki, MJ; Shneider, BL; Paul, JM; Heubi, JE. Glucocorticoids upregulate taurocholate transport by ileal brushborder membrane. Am. J. Physiol 1997, 273, G197–G203. [Google Scholar]
- Ibrahim, SA; Balasubramanian, KA. Comparative study on brush border membranes prepared from rat and monky small intestine by Ca2+ and Mg2+ precipitation. Comp. Biochem. Physiol 1995, 112B, 65–69. [Google Scholar]
- Prabhu, R; Balasubramanian, KA. A novel method of preparation of small intestinal brush border membrane vesicles by polyethylene glycol precipitation. Anal. Biochem 2001, 289, 157–161. [Google Scholar]
- Harley, SM; Beevers, H. Ricin inhibition of in vitro protein synthesis by plant ribosomes. Cell Biology 1982, 79, 5935–5938. [Google Scholar]
- Wesche, J; Rapak, A; Olsnes, S. Dependence of ricin toxicity on translocation of the toxin a-chain from the endoplasmic reticulum to the cytosol. J. Biol. Chem 1999, 274, 34443–34449. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
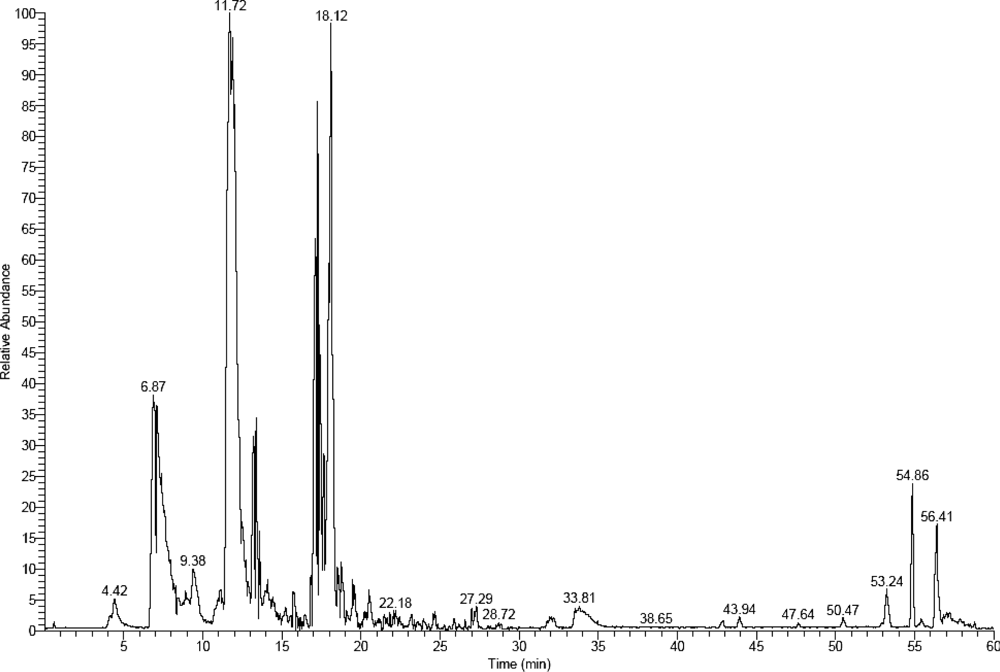{kind=link}
| NO. | Cover Percent (%) | MW(D) | PI | Identified Name |
|---|---|---|---|---|
| 1 | 1.27 | 208984.41 | 5.7 | sucrase-isomaltase |
| 2 | 2.35 | 112982.21 | 5.3 | Na+/K+ -ATPase alpha 1 subunit |
| 3 | 8.13 | 26134.64 | 4.75 | protease, serine, 1 |
| 4 | 1.18 | 87436.93 | 5.96 | dipeptidylpeptidase 4 |
| 5 | 2.82 | 81375.48 | 7.31 | Tripartite motif-containing protein 71 |
| 6 | 1.24 | 109651.12 | 5.62 | alanyl (membrane) aminopeptidase |
| 7 | 0.92 | 142634.26 | 4.26 | protocadherin 24 |
| 8 | 0.95 | 103676.35 | 5.49 | WD repeat domain 22 |
| 9 | 4.67 | 25110.87 | 9.14 | KDEL (Lys-Asp-Glu-Leu) ER protein retention receptor 3 |
| 10 | 1.28 | 79500.51 | 5.59 | meprin 1 beta |
| 11 | 1.57 | 71901.5 | 4.76 | Rab11-FIP4-like |
| 12 | 0.42 | 242072.22 | 6.54 | BAP28 protein |
| 13 | 0.32 | 278603.97 | 6.27 | X-linked nuclear protein |
| 14 | 2.67 | 43129.22 | 5.27 | guanylate binding protein 8 |
| 15 | 3.13 | 35989.95 | 5.43 | annexin A4 |
| 16 | 11.65 | 11368.23 | 11.18 | histone 4 protein |
| 17 | 2.37 | 49225.49 | 5.79 | secretion regulating guanine nucleotide exchange factor |
| 18 | 0.93 | 105319.96 | 5.6 | similar to neurofilament protein |
| 19 | 0.3 | 369730.12 | 4.97 | URE-binding protein 1 |
| 20 | 2.28 | 81229.48 | 9.28 | junctophilin 3 |
| 21 | 0.25 | 413981.02 | 5.46 | similar to maltase-glucoamylase |
| 22 | 0.57 | 221497.62 | 5.69 | Myosin cardiac muscle beta chain |
| 23 | 0.27 | 490969.47 | 6.11 | vacuolar protein sorting 13 D |
| 24 | 4.41 | 40820.65 | 5.32 | t-complex protein |
| 25 | 7.20 | 13591.96 | 9.21 | Chemokine-like protein TAFA-5. |
| 26 | 3.48 | 32599.11 | 5.95 | annexin A2 |
| 27 | 1.55 | 95321.42 | 5.21 | anaphase promoting complex subunit 2 |
| 28 | 6.39 | 31393.08 | 9.04 | killer cell lectin-like receptor, subfamily A, member 8 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, L.; Gao, H.; Li, J.; Dong, Y.; Liu, N.; Wan, J.; Liu, W.; Sun, Y.; Xu, M. Analysis of Intestinal Injuries Induced by Ricin in Vitro Using SPR Technology and MS Identification. Int. J. Mol. Sci. 2009, 10, 2431-2439. https://doi.org/10.3390/ijms10052431
Liu L, Gao H, Li J, Dong Y, Liu N, Wan J, Liu W, Sun Y, Xu M. Analysis of Intestinal Injuries Induced by Ricin in Vitro Using SPR Technology and MS Identification. International Journal of Molecular Sciences. 2009; 10(5):2431-2439. https://doi.org/10.3390/ijms10052431
Chicago/Turabian StyleLiu, Linna, Hongwei Gao, Jiping Li, Ying Dong, Ning Liu, Jiayu Wan, Wensen Liu, Yucheng Sun, and Ming Xu. 2009. "Analysis of Intestinal Injuries Induced by Ricin in Vitro Using SPR Technology and MS Identification" International Journal of Molecular Sciences 10, no. 5: 2431-2439. https://doi.org/10.3390/ijms10052431
APA StyleLiu, L., Gao, H., Li, J., Dong, Y., Liu, N., Wan, J., Liu, W., Sun, Y., & Xu, M. (2009). Analysis of Intestinal Injuries Induced by Ricin in Vitro Using SPR Technology and MS Identification. International Journal of Molecular Sciences, 10(5), 2431-2439. https://doi.org/10.3390/ijms10052431